 2015, Vol. 35
2015, Vol. 35文章信息
- 葛晓改, 曾立雄, 肖文发, 黄志霖, 周本智
- GE Xiaogai, ZENG Lixiong, XIAO Wenfa, HUANG Zhilin, ZHOU Benzhi
- 三峡库区森林凋落叶化学计量学性状变化及与分解速率的关系
- Dynamic of leaf litter stoichiometric traits dynamic and its relations with decomposition rates under three forest types in Three Gorges Reservoir Area
- 生态学报, 2015, 35(3): 779-787
- Acta Ecologica Sinica, 2015, 35(3): 779-787
- http://dx.doi.org/10.5846/stxb201304090653
-
文章历史
- 收稿日期:2013-4-9
- 网络出版日期:2014-04-03






2. 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091
2. State Forestry Administration Key Laboratory of Forest Ecology and Environment; Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
森林凋落物分解在森林生态系统物质循环、养分平衡[1]、维持土壤肥力中起着重要作用[2]。凋落物分解过程受基质质量、分解者种类和数量、环境因素(温度和湿度)和土壤养分供应的影响。在立地尺度上,凋落物基质质量是最重要的影响因素,例如,在森林生态系统中,凋落物N和P含量对分解有重要影响(尤其在分解的早期阶段),N、P养分的有效性是调节植物凋落物分解速率和生态系统碳平衡的一个主要因素。凋落叶化学计量学性状特征对分解者的生长和活动有重要的养分限制并广泛研究[3, 4],如Moore 等[5]在加拿大12a的凋落物分解研究表明:多数凋落物P比N释放快(和C紧密相关),C/N比在33—68时N释放,且与凋落物起始C/N比成正相关,而凋落物C/P比率为800—1200时,P可能释放;李雪峰等[6]对长白山次生针阔混交林凋落叶分解与C、N、P释放关系研究表明,分解速率与凋落物起始N含量、N/P比呈显著正相关,而与C/N比、C/P比呈显著负相关,凋落物N、P剩余百分率与凋落叶干重剩余百分率呈显著二项式回归关系。这些研究探讨了凋落物化学计量学性状与分解速率的关系及其影响,对凋落物分解养分释放过程的研究具有重要意义。也有一些研究,如闫恩荣等[7]、王希华等[8]、陈法霖等[9]探讨了凋落物起始化学计量学性状与分解速率关系,但对凋落物化学计量学性状变化过程与分解速率关系探讨较少,后者某种程度上更能真实反映凋落物化学计量学性状变化与分解速率关系。因此,开展凋落物化学计量学性状的动态变化及与分解速率关系的研究,对凋落物分解过程及凋落物-土壤生物化学连续体的深层理解有重要意义,对揭示C、N、P元素之间的交互作用、平衡制约关系和促进生态化学计量学理论的发展具有科学意义[10]。
三峡库区地处中亚热带北缘,气候和植被处于过渡状态,是我国生态环境保护的重点地区。该地区复杂的地形变化,使该地区的生态系统十分敏感和脆弱。特别是,2010年10月三峡大坝蓄水为175 m后,长江大坝蓄水面积增加,蒸发量也必然大幅增加,降水量很可能改变库区气候环境,对该区域范围内凋落物分解及生态系统养分循环等存在潜在影响。对研究区域生态系统凋落叶化学计量学性状和分解动态变化的研究,可以深入认识植物-凋落物-土壤相互作用的养分调控因素,对认识生态系统碳汇潜力和生态系统如何响应未来气候变化具有重要意义。本研究从不同林型凋落叶分解和养分释放动态的角度,对三峡库区典型树种马尾松(Pinus massoniana,PM)林、栓皮栎(Quercus variabilis,QV)林和马尾松-栓皮栎混交林(P. massoniana- Q. variabilis mixed forest,PQ)凋落叶化学计量学性状动态变化及与分解速率关系进行研究,为了解凋落叶自身性质对分解速率及养分释放等物质循环状况有重要意义,为揭示凋落叶分解过程中C、N、P的释放机制和凋落物分解模型构建及修订奠定基础,为三峡库区森林的养分循环和可持续经营提供理论依据。
1 研究地区与研究方法 1.1 研究区概况研究区位于湖北省秭归县(30°38′14″—31°11′31″N,110°00′14″—111°18′41″E),海拔40—2057 m,该地区属于亚热带大陆性季风气候,年均气温16.9 ℃,极端最高气温达44 ℃,极端最低气温-2.5 ℃,年降水量1000—1250 mm,降水集中在4—9月,空气湿度达60%—80%,无霜期300—340 d[11, 12]。秭归县海拔500 m以下以茶园、板栗、柑橘和农田为主,海拔500 m以上主要为灌丛、马尾松、杉木(Cunninghamia lanceolata)和松栎等[13]。马尾松林集中分布于1200 m以下山坡的中、下部或丘陵低山的上部;栓皮栎林主要分布于海拔1350 m以下低山、丘陵地带,多为纯林,林分均匀整齐,郁闭度0.5—0.7,一般10—15 m,有些地区与马尾松组成马尾松-栓皮栎混交林。
以长江三峡库区秭归森林生态定位站附近的马尾松林、栓皮栎林、马尾松-栓皮栎混交林为研究对象,马尾松林下灌木多为油茶(Camellia oleifera)和檵木(Loropetalum chinense)、毛黄栌(Cotinus coggygria)等,草本多为稗草(Echinochloa crusgalli)、腹水草(Veronicastrum axillare)等[14];栓皮栎林下灌木层主要有映山红(Rhododendron simsii)、猫儿刺(Ilex cornuta)、马桑(Coriaria nepalensis)等,草本主要有白茅(Imperata cylindrica)、芒(Miscanthus sinensis)、苔草(Carex gmelinii)等;马尾松-栓皮栎混交林中有少量的杉木、枫香(Liquidambar styraciflua),且马尾松和栓皮栎分别占60%和40%;草本多为稗草、腹水草等。研究样地概况见表 1。
| 林分 Stand | 海拔/m Elevation | 林龄/a Age | 胸径/cm DBH | 坡度/(°) Slope | 坡向 Aspect | 郁闭度 Canopy density | 土壤厚度/cm Soil depth |
| DBH:胸径Diameter at breast height; PM:马尾松Pinus massoniana; QV:栓皮栎Quercus variabilis; PQ:马尾松-栓皮栎混交林P. massoniana-Q. variabilis mixed forest; S: 阳坡Sunny; Ss: 半阴坡Semi-sunny; Sh: 阴坡Shady | |||||||
| 马尾松林 PM | 357 | 30 | 25.3 | 25 | Ss | 0.95 | 60—80 |
| 栓皮栎林 QV | 342 | 19 | 16.1 | 15 | S | 0.60 | 50—80 |
| 马尾松-栓皮栎混交林PQ | 290 | 22 | 20.4 | 10 | Sh | 0.80 | 70—100 |
凋落叶分解研究采用分解袋法。在马尾松林、栓皮栎林和马尾松-栓皮栎混交林中分别设置3个20 m×30 m的样方。于2010年11月在3种林地样方外分别收集自然凋落、上层未分解的马尾松和栓皮栎凋落叶(马尾松-栓皮栎混交林指马尾松和栓皮栎凋落叶),同一样地收集的凋落叶充分混合风干后,称取凋落叶15 g,直接装入尼龙分解袋中。根据研究区域马尾松-栓皮栎混交林中马尾松和栓皮栎分别占60%和40%,本研究确定混合凋落物中马尾松和栓皮栎凋落叶重量比为3 ∶ 2,与马尾松-栓皮栎混交林中栓皮栎与马尾松树种比例保持一致。尼龙网袋大小为20 cm×20 cm,孔径1 mm。同时,称取3林分凋落叶各15 g,重复3次,在70 ℃下烘干至恒重,算出风干与烘干重转换系数。
于2010年11月底将装有凋落叶的分解袋随机放置在各自的林地样方中,除去地表凋落物层,使凋落物分解袋贴近地面,让凋落物自然分解;放置凋落物分解袋后每月底进行新凋落物清理。每种林地放置90袋,3种林分共270袋。自放置日起,每3个月每林分取回18袋凋落物;取回凋落物分解袋后,清除侵入的根系、泥沙,烘至恒量,测干重并计算干质量剩余率和分解速率。
1.2.2 样品测定凋落叶化学分析:将收集的样品烘干磨碎,过0.2 mm筛,用重铬酸钾氧化-外加热法测全C;用凯氏定氮法测全N(意大利,UK152);用等离子发射光谱法测全P(美国,IRIS Intrepid ⅡXSP)。凋落叶样品共测定6次,即起始凋落叶样品和5次分解样品。
1.3 数据处理与统计分析凋落叶分解干质量剩余率计算公式采用:
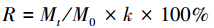
式中,R为凋落叶分解干质量剩余率(%);k为凋落叶风干样品与烘干样品转换系数;Mt为凋落叶t时间烘干样品重量;M0为凋落叶起始风干样品重量。
凋落叶分解率计算公式采用:

式中,L为凋落物分解率(%);R为凋落物分解干质量剩余率(%)。
凋落叶干质量剩余率用Olson[15]负指数衰减模型进行拟合:
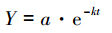
式中,Y为干质量剩余率(%);a为拟合参数;k为年分解系数(kg kg-1a-1);t为分解时间(a)。凋落叶分解半衰期(50%分解)计算式,t0.05=ln0.5/(-k);完全分解时间(95%分解)计算式,t0.95=ln0.05/(-k)。
凋落叶分解过程中养分释放速率按下式计算:
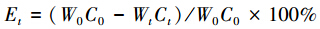
式中,W0为凋落叶初始质量,C0为凋落叶初始养分质量分数;Wt为分解t时刻剩余凋落叶质量;Ct为分解t时刻的养分质量分数。
利用Excell 2003 和SPSS 16.0软件对数据进行统计分析和作图。凋落叶分解速率、基质质量含量的显著性检验采用单因子方差分析(ANOVA)和最小显著差异法(LSD),显著性水平设定为α=0.05。图表中数据均用平均值±标准差表示。
2 结果与分析 2.1 不同林型凋落叶分解动态不同林型凋落叶分解速率大小顺序是马尾松林<栓皮栎林<马尾松-栓皮栎混交林(图 1)。随着分解的进行,分解速率差距越来越大,分解450 d后马尾松林、栓皮栎林和马尾松-栓皮栎混交林凋落叶干质量剩余率分别是67.59%、64.75%、62.13%;马尾松林、栓皮栎林、马尾松-栓皮栎混交林凋落叶年分解常数k分别是0.33、0.35和0.38,凋落叶分解50%和95%的时间分别是2.11 a和9.15 a,1.93 a和8.45 a,1.76 a和7.77 a(表 2)。

|
| 图 1 三峡库区不同林型凋落叶分解动态 Fig. 1 Litter leaf decomposition dynamic on different forest types in the Three Gorges Reservoir Area |
| 林分 Stand | 回归方程 Regression equation | R2 | k | 50% | 95% |
| PM:马尾松Pinus massoniana; QV:栓皮栎Quercus variabilis; PQ:马尾松-栓皮栎混交林P. massoniana-Q. variabilis mixed forest | |||||
| 马尾松林PM | y=99.762 e-0.3272 t | 0.9689 | 0.327 | 2.11 | 9.15 |
| 栓皮栎林QV | y=98.81 e-0.353 t | 0.9866 | 0.353 | 1.93 | 8.45 |
| 马尾松-栓皮栎混交林 PQ | y=98.03 e-0.383 t | 0.9781 | 0.383 | 1.76 | 7.77 |
在3种林地中,随着凋落叶分解,剩余凋落叶中C、P含量明显降低、而N增加不显著(图 2)。各自林下凋落叶起始C含量大小顺序是马尾松-栓皮栎混交林<栓皮栎林<马尾松林,C含量在分解过程中均降低且先慢后快,分解450 d后是栓皮栎林<马尾松林<马尾松-栓皮栎混交林,分别释放12.34%、13.31%和16.45%,马尾松和栓皮栎林在分解180—270 d期间释放最快,马尾松-栓皮栎混交林则在分解270—360 d间释放最快。起始凋落叶N含量是栓皮栎林最高,马尾松林最低,分解450 d后马尾松林、栓皮栎林、马尾松-栓皮栎混交林分别增加了66.67%、44.91%和44.52%,马尾松林N含量持续增加,混交林则波动较大。栓皮栎林、马尾松-栓皮栎混交林凋落叶起始P含量分别是马尾松林分的2.50、2.12倍;马尾松林降低较缓慢,栓皮栎林和马尾松-栓皮栎林在分解的0—90 d中降低较快,分解450 d后,P含量大小顺序是马尾松林<马尾松-栓皮栎混交林<栓皮栎林,分别释放了30.80%、42.29%、38.89%。

|
| 图 2 三峡库区不同林型凋落叶化学计量学性状动态变化 Fig. 2 The stoichiometric traits dynamics of litter leaf on different forest types in the Three Gorges Reservoir Area |
不同林型凋落叶化学计量学性状动态变化明显(图 2)。在分解的起始阶段,不同林型凋落叶C/N比是栓皮栎林<马尾松-栓皮栎混交林<马尾松林,随着分解的进行,栓皮栎林、马尾松林凋落叶C/N比持续降低,分解450 d后栓皮栎林、马尾松林、马尾松-栓皮栎混交林凋落叶C/N比分别下降了39.61%、47.48%、42.20%。起始马尾松林凋落叶C/P比分别是栓皮栎林、马尾松-栓皮栎混交林2.69倍、2.39倍,随着分解时间的进行,C/P比均增加,马尾松和栓皮栎林波动较马尾松-栓皮栎混交林大,分解450 d后栓皮栎林、马尾松林和马尾松-栓皮栎混交林分别增加43.67%、23.19%、44.01%。起始凋落叶N/P比是马尾松-栓皮栎混交林<栓皮栎林<马尾松林,马尾松林与栓皮栎林、马尾松-栓皮栎混交林差异显著,分解450 d后马尾松林分别是栓皮栎林、马尾松-栓皮栎混交林的2.04倍、2.10倍,分别增加137.67%、141.34%、150.89%。
2.3 不同林型凋落叶化学计量学性状动态变化与分解速率的关系凋落叶不同化学计量学性状与分解速率关系不同(图 3)。3种林型凋落叶C含量、C/N比均与凋落叶分解速率呈负相关,即C含量、C/N比越高,分解越慢(P<0.01)。3种林型凋落叶N含量均与凋落叶分解速率呈正相关,即N含量越高,分解越快(P<0.05)。3种林型凋落叶基质质量中的P含量动态与凋落叶分解速率关系不一致但差异显著,与栓皮栎林、马尾松林呈负相关,与马尾松-栓皮栎混交林呈二次函数关系。3种林型凋落叶分解中N/P比动态与凋落叶分解速率呈极显著负二次函数关系(P<0.01),而C/P比动态与凋落叶分解速率均不显著(P>0.05)。

|
| 图 3 三峡库区不同林型凋落叶分解速率与化学计量学性状动态变化关系 Fig. 3 Relationships between litter leaf stoichiometric traits dynamic and decomposition rate on different forest types in the Three Gorges Reservoir Area |
不同林型凋落叶化学性状与分解速率关系不同(表 3)。3种林型凋落叶C、C/N比、N/P比动态与分解速率一致且均为极显著水平;3种林型凋落叶N含量均呈极显著正相关,3林型凋落叶P含量动态与分解速率关系、显著水平均不一致。
| 化学计量学性状 Stoichiometric traits | 林型 Stand type | 回归方程 Regression equation | R2 | P |
| PM:马尾松Pinus massoniana; QV:栓皮栎Quercus variabilis; PQ:马尾松-栓皮栎混交林P. massoniana- Q. variabilis mixed forest | ||||
| C | 马尾松PM | y=-2.55x+143.60 | 0.57 | 0.00 |
| 栓皮栎QV | y=-3.07x+166.98 | 0.56 | 0.00 | |
| 马尾松-栓皮栎混交林 PQ | y=-2.21x+121.36 | 0.59 | 0.00 | |
| N | 马尾松PM | y=39.96x-27.49 | 0.79 | 0.00 |
| 栓皮栎 QV | y=47.90x-43.80 | 0.79 | 0.00 | |
| 马尾松-栓皮栎混交林 PQ | y=34.42x-17.4 | 0.61 | 0.00 | |
| P | 马尾松PM | y=-906.96x+56.38 | 0.47 | 0.00 |
| 栓皮栎 QV | y=-257.59x+44.20 | 0.41 | 0.00 | |
| 马尾松-栓皮栎混交林 PQ | y=-72.26x2+210.21x-120.57 | 0.45 | 0.02 | |
| C/N比 C/N ratio | 马尾松PM | y=-0.96x+61.46 | 0.80 | 0.00 |
| 栓皮栎 QV | y=-1.27x+67.44 | 0.78 | 0.00 | |
| 马尾松-栓皮栎混交林 PQ | y=-1.13x+68.34 | 0.70 | 0.00 | |
| C/P比 C/P ratio | 马尾松PM | y=0.03x-14.66 | 0.19 | 0.07 |
| 栓皮栎 QV | y=0.02x+11.07 | 0.12 | 0.18 | |
| 马尾松-栓皮栎混交林 PQ | y=0.02x+12.57 | 0.02 | 0.59 | |
| N/P比 N/P ratio | 马尾松PM | y=-0.01x2+1.92x-24.71 | 0.76 | 0.00 |
| 栓皮栎 QV | y=-0.09x2+5.27x-34.85 | 0.87 | 0.00 | |
| 马尾松-栓皮栎混交林 PQ | y=-0.23x2+8.37x-46.68 | 0.64 | 0.00 | |
凋落叶N、P含量动态对分解速率影响明显。Berg和Matzner[16]研究表明分解初期,N含量高的凋落物比N含量低的凋落物分解快。陈瑾等[17]对内蒙古草原凋落叶分解研究表明,单物种凋落叶分解速率与初始N 含量呈显著正相关;这与本研究中起始N含量高的栓皮栎林比N含量低的马尾松林分解快一致。相反, Hoorens等[18]和Xuluc-Tolosa等[19]研究均表明凋落物分解率和内源P浓度相关,和N浓度不相关。Li等[20]对白草(Pennisetum accidum)、猪毛蒿(Artemisia scoparia)、尖头藜科(Chenopodium acuminatum)、大麻(Cannabis sativa)的凋落叶分解研究发现凋落物分解速率与起始阶段的凋落叶起始N含量呈正相关,与分解后期阶段凋落物N、P含量呈负相关。Huang等[21]在中国东部凋落叶分解研究表明凋落物分解率与凋落叶起始N但不是P呈正相关。本研究表明凋落叶分解速率与N、P含量均紧密相关,这说明凋落叶基质质量对分解速率的影响与物种种类、环境条件有很大关系。此外,也可能因为土壤养分对凋落叶分解有调节作用,低P土壤比低N土壤对分解的限制作用更强[20];凋落叶在养分含量高的林地中分解更快,在养分贫瘠土壤中的植物凋落叶分解慢,因为土壤中的养分含量越低,凋落叶基质质量C/N比越高,耐分解化合物的含量越多,凋落叶分解越慢[3]。
不同凋落叶化学计量学性状对分解速率的影响不同。李雪峰等[6]研究表明凋落叶分解速率与C/N比、C/P比呈负相关;Mooshammer等[22]研究表明山毛榉(Fagus sylvatica)凋落叶C/N比对凋落物分解速率影响较大,C/P比、N/P比则影响不明显。本研究中3种林型凋落叶分解速率与C/N比呈负相关,与C/P比差异不显著(表 3),前者与Melillo等[23]研究结果一致,即起始凋落叶C/N比在含量N最高时下降到86,在净矿化后继续下降;后者与李雪峰等[6]研究结果不太一致;这可能因为凋落叶P含量在分解初期开始释放,很难确定P释放时关键的C/P比,且森林地面养分影响N和P动态,土壤表面C/N比、C/P比越高,凋落叶N、P释放越快(和C相关)[5]。潘复静等[24]研究表明N/P比值是制约凋落叶分解和养分循环的重要因素,凋落叶分解速率与N/P比成反比,与本研究中3种林型凋落叶N/P比与凋落叶分解速率呈二次函数关系且达到极显著水平(表 3)不太一致,说明凋落叶化学计量学性状与分解速率的相关关系因所研究的凋落物类型、研究地点和试验周期的不同而存在差异,即外界环境条件会显著改变凋落物的分解规律[6]。
3.2 凋落叶分解过程化学计量学性状动态变化不同林型凋落叶分解主要养分释放模式不同。林开敏等[25]对凋落叶分解360 d的研究结果表明:木荷(Schima superba)叶凋落物中N、P变化较大,木荷N含量变化幅度比杉木(Cunninghamia lanceolata)和楠木(Phoebe bournei)叶凋落物大;本研究中栓皮栎林N固持(44.91%)较马尾松林(66.67%)大,说明研究时间长短、植被类型对养分利用效率有较大的影响;Aerts[26]研究表明常绿树叶的光合作用时间较长,与落叶树种相比,具有较高的养分利用效率。李雪峰等[6]对长白山次生针阔混交林凋落叶中有机物分解与C、N、P释放的关系表明,凋落叶N、P含量随分解逐渐增加并达到峰值,9种类型凋落叶分解剩余率的变化范围分别在45.3%—58.9%和44.4%—64.4%之间时,N和P进入矿化阶段;可能因为高纬度地区的植物更易受N元素限制,而低纬度地区的植物更易受P元素的限制,但受N或P限制的植物并不一定具有高的N或P再吸收效率,环境因子对不同类型植物化学计量性状的影响不相同[27]。
同一物种凋落叶分解主要养分释放模式不同。McClaugherty等[28]在美国威斯康星洲对6类型森林凋落叶分解研究表明,凋落叶N含量是先积累,但所有凋落叶N含量开始释放是在培养2 a的末期;N含量随着失重的积累线性增加;Quested等[29]研究表明凋落叶N在生长季释放,春季和秋季对凋落叶养分释放和N动态很重要;邓仁菊等[30]对季节性冻融期间凋落物分解研究表明:冷杉(Abies faxoniana)和白桦(Betula platyphylla)凋落物C、N、P均释放。刘颖等[31]于2003年5月—2004年9月在长白山对4种森林凋落物分解研究表明C含量变化不大,N和P含量先上升后下降。本研究中N含量在研究期间一直固持,C、P含量一直释放,与季节变化没有明显关系,可能因为不同地区树种不同分解阶段所处的环境(如水分、温度、分解的微生物和动物数量等)和凋落物自身特性(如质地和化学组成等)有关。因此,凋落物混合分解相互作用是一个较为复杂的问题,究竟表现何种形式相互作用,是促进、抑制或无影响,仍无规律可循[32]。
3.3 化学计量学性状对凋落叶分解养分动态的影响化学计量学能更好地揭示生态系统各组分(植物、凋落物和土壤)养分比例的调控机制,阐明生态系统碳氮磷平衡的元素化学计量比格局,对于揭示元素相互作用变化规律,实现自然资源的可持续利用具有重要的现实意义[10]。Manzoni等[33]研究表明化学计量学在不同的气候区域和生态系统对凋落物养分动态的影响普遍存在;在分解的起始阶段,在北方和温暖气候区域N和P易于固持(如C/N比和C/P比与起始阶段相比较低),而在热带区域N通常释放,P通常固持或释放,且在关键C/N比和C/P比以下,养分释放,随着起始凋落物N和P浓度的减少而增加。本研究中凋落叶分解450 d后,马尾松林凋落叶C/N比下降速率与N固持的速率呈正比,与P含量释放的速率呈反比,C/P比增加的速率则相反,栓皮栎林和混交林变化类似但不明显;因为凋落叶分解速率与凋落物起始化学组成关系密切,不同的植物物种凋落物化学计量学性状对凋落物分解养分动态影响不同;低木质素含量、高氮磷含量的阔叶树种有利于加速凋落物的分解速率和土壤肥力的恢复进程[5]。Moore等[5]研究表明:凋落物起始P浓度与P释放模式明显相关,高P浓度凋落物释放P,低P浓度凋落物固持P,关键的C/P比是700—900,N释放的关键C/N比是37—51。Güsewell和Verhoeven[34]研究表明:凋落物N、P浓度和化学计量学性状影响凋落物早期分解速率,禾本科植物凋落物关键N/P比是25;当凋落物分解速率与N含量相关且N/P比较低,或与P相关且有N/P比较高时,N/P比可以作为N或P含量是否限制分解的指标。由于本研究中凋落叶分解研究仅进行了15个月,可能仍处于分解的早期阶段,没有观测到N含量释放的化学计量学性状关键值,仍需进一步深入研究。
总之,在森林生态系统中凋落叶分解过程中N和P是常见的限制因素;本研究中凋落叶N含量固持,C、P含量释放,且与季节变化没有明显关系。3种林型凋落叶分解速率均与N含量呈正相关 (P<0.01),与C含量 (P<0.01)、C/N比 (P<0.01)呈负相关,与N/P比呈负二次函数关系 (P<0.01),而P含量与3种林型关系不同。混交林中栓皮栎和马尾松林凋落叶起始化学计量学性状差异较大是马尾松-栓皮栎混交林分解较快的原因之一,化学计量学性状对揭示元素相互作用变化规律及养分循环和生态系统功能上的研究很重要,可为三峡库区森林的养分循环和可持续经营提供理论依据。
| [1] | 宋新章, 江洪, 张慧玲, 余树全, 周国模, 马元丹, Chang S X. 全球环境变化对森林凋落物分解的影响. 生态学报, 2008, 28(9): 4414-4423. |
| [2] | 潘开文, 何静, 吴宁. 森林凋落物对林地微生境的影响. 应用生态学报, 2004, 15(1): 153-158. |
| [3] | 彭少麟, 刘强. 森林凋落物动态及其对全球变暖的响应. 生态学报, 2002, 22(9): 1534-1544. |
| [4] | Moore T R, Trofymow J A, Taylor B, Prescott C, Camiré C, Duschene L, Fyles J, Kozak L, Kranabetter M, Morrison I, Siltanen M, Smith S, Titus B, Visser S, Wein R, Zoltai S. Litter decomposition rates in Canadian forests. Global Change Biology, 1999, 5(1): 75-82. |
| [5] | Moore T R, Trofymow J A, Prescott C E, Titus B D, Cidet Working Group. Nature and nurture in the dynamics of C, N and P during litter decomposition in Canadian forests. Plant and Soil, 2011, 339(1-2): 163-175. |
| [6] | 李雪峰, 韩士杰, 胡艳玲, 赵玉涛. 长白山次生针阔混交林叶凋落物中有机物分解与碳、氮和磷释放的关系. 应用生态学报, 2008, 19(2): 245-251. |
| [7] | 阎恩荣, 王希华, 郭明, 仲强, 周武. 浙江天童常绿阔叶林、常绿针叶林与落叶阔叶林的C:N:P化学计量特征. 植物生态学报, 2010, 34(1): 48-57. |
| [8] | 王希华, 黄建军, 闫恩荣. 天童国家森林公园常见植物凋落叶分解的研究. 植物生态学报, 2004, 28(4): 457-467. |
| [9] | 陈法霖, 江波, 张凯, 郑华, 肖燚, 欧阳志云, 屠乃美. 退化红壤丘陵区森林凋落物初始化学组成与分解速率的关系. 应用生态学报, 2011, 22(3): 565-570. |
| [10] | 王绍强, 于贵瑞. 生态系统碳氮磷元素的生态化学计量学特征. 生态学报, 2008, 28(8): 3937-3947. |
| [11] | 雷静品, 肖文发, 黄志霖, 曾立雄. 三峡库区秭归县不同海拔马尾松径向生长对气候的响应. 林业科学, 2009, 45(2): 33-39. |
| [12] | 姜欢欢, 李继红, 范文义, 鲍晨光. 三峡库区秭归县景观格局变化及模拟预测. 应用生态学报, 2009, 20(2): 474-479. |
| [13] | 曾立雄, 肖文发, 黄志霖, 沈亲, 马德举. 兰陵溪小流域主要退耕还林植被土壤渗透特征. 水土保持学报, 2010, 24(3): 199-202. |
| [14] | 肖文发, 李建文, 于长青, 程瑞梅, 刘少英, 王金锡, 葛继稳. 长江三峡库区陆生动植物生态. 重庆: 西南师范大学出版社, 2000. |
| [15] | Olson J S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 1963, 44(2): 322-331. |
| [16] | Berg B, Matzner E. Effect of N deposition on decomposition of plant litter and soil organic matter in forest systems. Environmental Reviews, 1997, 5(1): 1-25. |
| [17] | 陈瑾, 李扬, 黄建辉. 内蒙古典型草原4种优势植物凋落物的混合分解研究. 植物生态学报, 2011, 35(1): 9-16. |
| [18] | Hoorens B, Aerts R, Stroetenga M. Does initial litter chemistry explain litter mixture effects on decomposition? Oecologia, 2003, 137(4): 578-586. |
| [19] | Xuluc-Tolosa F J, Vester H F M, Ramírez-Marcial N, Castellanos-Albores J, Lawrence D. Leaf litter decomposition of tree species in three successional phases of tropical dry secondary forest in Campeche, Mexico. Forest Ecology and Management, 2003, 174(1-3): 401-412. |
| [20] | Li L J, Zeng D H, Yu Z Y, Fan Z P, Yang D, Liu Y X. Impact of litter quality and soil nutrient availability on leaf decomposition rate in a semi-arid grassland of Northeast China. Journal of Arid Environments, 2011, 75(9): 787-792. |
| [21] | Huang J J, Wang X H, Yan E R. Leaf nutrient concentration, nutrient resorption and litter decomposition in an evergreen broad-leaved forest in eastern China. Forest Ecology and Management, 2007, 239(1-3): 150-158. |
| [22] | Mooshammer M, Wanek W, Schnecker J, Wild B, Leitner S, Hofhansl F, Blöchl A, HLämmerle I, Frank A H, Fuchslueger L, Keiblinger K M, Zechmester-Boltenstern S, Richter A. Stoichiometric controls of nitrogen and phosphorus cycling in decomposing beech leaf litter. Ecology, 2012, 93(4): 770-782. |
| [23] | Melillo J M, Aber J D, Muratore J F. Nitrogen and lignin control of hardwood leaf litter decomposition dynamics. Ecology, 1982, 63(3): 621-626. |
| [24] | 潘复静, 张伟, 王克林, 何寻阳, 梁士楚, 韦国富. 典型喀斯特峰丛洼地植被群落凋落物C:N:P生态化学计量特征. 生态学报, 2011, 31(2): 335-343. |
| [25] | 林开敏, 章志琴, 叶发茂, 林艳, 李卿叁. 杉木人工林下杉木、楠木和木荷叶凋落物分解特征及营养元素含量变化的动态分析. 植物资源与环境学报, 2010, 19(2): 34-39. |
| [26] | Aerts R. Nutrient use efficiency in evergreen and deciduous species from heathlands. Oecologia, 1990, 84(3): 391-397. |
| [27] | 王晶苑, 王绍强, 李纫兰, 闫俊华, 沙丽清, 韩士杰. 中国四种森林类型主要优势植物的C:N:P化学计量学特征. 植物生态学报, 2011, 35(6): 587-595. |
| [28] | McClaugherty C A, Pastor J, Aber J D, Melillo J M. Forest litter decomposition in relation to soil nitrogen dynamics and litter quality. Ecology, 1985, 66(1): 266-275. |
| [29] | Quested H M, Callaghan T V, Cornelissen J H C, Press M C. The impact of hemiparasitic plant litter on decomposition: direct, seasonal and litter mixing effects. Journal of Ecology, 2005, 93(1): 87-98. |
| [30] | 邓仁菊, 杨万勤, 冯瑞芳, 胡建利, 秦嘉励, 熊雪晶. 季节性冻融期间亚高山森林凋落物的质量损失及元素释放. 生态学报, 2009, 29(10): 5730-5735. |
| [31] | 刘颖, 韩士杰, 林鹿. 长白山4种森林凋落物分解过程中养分动态变化. 东北林业大学学报, 2009, 37(8): 28-30. |
| [32] | 林开敏, 章志琴, 邹双全, 曹光球. 杉木与阔叶树叶凋落物混合分解对土壤性质的影响. 土壤通报, 2006, 37(2): 258-262. |
| [33] | Manzoni S, Trofymow J A, Jackson R B, Porporato A. Stoichiometric controls on carbon, nitrogen, and phosphorus dynamics in decomposing litter. Ecological Monographs, 2010, 80(1): 89-106. |
| [34] | Güsewell S, Verhoeven J T A. Litter N:P ratios indicate whether N or P limits the decomposability of graminoid leaf litter. Plant and Soil, 2006, 287(1-2): 131-143. |