 2015, Vol. 35
2015, Vol. 35文章信息
- 张忠华, 胡刚, 倪健
- ZHANG Zhonghua, HU Gang, NI Jian
- 茂兰喀斯特常绿落叶阔叶混交林树种的空间分布格局及其分形特征
- Spatial distribution patterns and their fractal properties for trees in a subtropical mixed evergreen-deciduous broad-leaved karst forest in Maolan, southwestern China
- 生态学报, 2015, 35(24): 8221-8230
- Acta Ecologica Sinica, 2015, 35(24): 8221-8230
- http://dx.doi.org/10.5846/stxb201406061172
-
文章历史
- 收稿日期: 2014-06-06
- 网络出版日期: 2015-05-21






2. 广西师范学院北部湾环境演变与资源利用教育部重点实验室, 南宁 530001;
3. 中国科学院地球化学研究所环境地球化学国家重点实验室, 贵阳 550002;
4. 中国科学院普定喀斯特生态系统观测研究站, 普定 562100
2. Key Laboratory of Beibu Gulf Environmental Change and Resources Utilization of Ministry of Education, Guangxi Teachers Education University, Nanning 530001, China;
3. State Key Laboratory of Environmental Geochemistry, Institute of Geochemistry, Chinese Academy of Sciences, Guiyang 550002, China;
4. Puding Karst Ecosystem Observation and Research Station, Chinese Academy of Sciences, Puding 562100, China
植物种群的空间分布格局及其形成机制是生态学研究的重要内容[1, 2]。通过空间分布格局研究可以分析种群的生物学特征、种内和种间的竞争关系、种群的动态、群落的构建以及植被的演变过程等信息,从而推断种群时空格局形成的潜在生态过程和驱动机制[3, 4, 5]。种群的空间分布格局不仅因种而异,并且同一种在不同生长阶段、不同生境条件下也有明显差异,种群空间格局是时空尺度上生物与非生物因素及其交互作用结果的综合体现。因此,判断格局形成的关键因素和过程往往较为困难[4]。近年来,伴随格局类型判定的计测方法不断改进,国内外学者在全球的热带、亚热带和温带等区域的森林生态系统中进行了广泛的种群空间分布格局方面研究[2, 6, 7, 8, 9],但对格局形成机制的认识仍然十分有限。
植物种群的空间分布格局主要包括聚集、随机和均匀3种类型,其量化判定方法主要包括以植物个体频度和多度为数据基础的方差均值比、负二项分布等方法、以个体距离数据为基础的角尺度法和最近邻体法以及以个体空间相对坐标位点为数据基础的点格局分析法[1]。20世纪70年代,分形几何学中提出的分形维数可用于量度和刻画对象的几何复杂程度和充斥空间的能力,可对不规则但又具自相似特性的几何形体的复杂程度进行定量的分析和比较[10]。作为分析空间复杂性和变异程度的有效方法,分形维数已被广泛应用于定量描述植物体的形态结构[11]、种群空间分布[12]、群落结构动态[13]以及大尺度景观格局[14]等生态学领域。植物种群的空间分布形态可看作分形体,可运用分形维数来量度其空间结构的不规则性和复杂性[15, 16]。我国学者马克明[17]和祖元刚[18]较早将分形理论引入植物生态学领域,并在羊草(Aneurolepidium chinense)草原的群落格局研究方面取得了良好效果。张文辉[19]、梁士楚[20]和宋于洋[21]等学者分别对裂叶沙参(Adenophora lobophylla)、木榄(Bruguiera gymnorhiza)和梭梭(Haloxylon ammodendron)等不同植被类型中的优势种群分布格局的分形特征进行了研究,发现分形理论是研究种群空间分布格局的一种有效办法,弥补了传统方法中的某些不足。虽然,上述研究利用分形理论在不同生态系统中对种群空间分布进行了丰富探索,并推论出诸多重要研究结果,但也存在一定的不足。首先,多数研究仅选取群落中的某一建群种或少数几个优势种为研究对象,缺乏对群落中不同种群空间分布分形特征的整体认识;其次,罕有研究揭示树木不同生长阶段空间分布格局的分形属性变化规律;再次,种群空间分布格局的分形维数与种群聚集程度之间的相关关系也鲜有文献报道。
位于我国黔南茂兰国家级自然保护区的中亚热带喀斯特森林,是同纬度上仅存的保存最为完好的非地带性喀斯特植被,已知的维管植物多达143科501属1203种[22]。由于其独特的石灰岩地质地貌背景和复杂多样的生境类型,形成了丰富的植物种类组成、特殊的植物区系成份和复杂的群落结构,喀斯特植被对维持岩溶生态系统结构和功能的稳定性极为重要,同时对探索岩溶地质背景下诸多生态过程与驱动机制具有极高科研价值。目前为止,国内学者对茂兰喀斯特森林的植物种类组成[22, 23, 24]、种群结构与动态[25, 26]、群落分布与环境关系[27, 28]以及植被恢复重建[29, 30]等进行了丰富的研究。笔者曾运用点格局方法探讨茂兰喀斯特森林树种的空间分布格局[5, 26],但空间格局的分形特征尚未报道。本研究以茂兰国家级自然保护区的中亚热带喀斯特常绿落叶阔叶混交林为研究对象,以建立的1 hm2(100 m×100 m)永久样地为数据来源,在运用4种格局强度指数对样地中30种树木进行空间分布格局分析的基础上,着重应用分形理论中的计盒维数和信息维数分析空间分布的分形属性。本研究主要关注如下问题:(1)茂兰中亚热带喀斯特常绿落叶阔叶混交林中树种有着何种类型的空间分布格局及其分形特征?(2)树木不同生长阶段(幼树、中树和成树)空间分布的分形维数如何变化?(3)种群空间分布的分形维数是否可以表征聚集强度和多度等种群结构属性?
1 研究区概况茂兰国家级自然保护区位于贵州省南部黔、桂交界处(25°09′—25°21′N,107°52′—08°05′E),总面积约212.85 km2,属于中亚热带石灰岩区常绿落叶阔叶混交林生态系统,是世界上喀斯特地貌区幸存的连片面积最大、保存完整且原生性强的喀斯特森林,该保护区于1996年加入联合国教科文组织“人与生物圈”保护区网,2007年被联合国教科文组织审定为世界自然文化遗产。保护区地貌类型以峰丛洼地和峰丛漏斗为主,海拔430—1078.6 m,森林覆盖率约为87%。全区除局部地点覆盖有少量砂页岩外,主要是由纯质石灰岩及白云岩构成的典型喀斯特地貌,岩石裸露率可达90%以上。该区气候属于中亚热带季风湿润气候,气候温和,雨量充沛。年平均气温为15.3 ℃,7月平均气温26.4 ℃,1月平均气温8.3 ℃,≥10 ℃积温5727.9 ℃,平均年降水量1750 mm,平均年蒸发量1343.6 mm,全年平均相对湿度83%,年日照时数1272.8 h,日照百分率29%。
2 研究方法 2.1 样地设置于2008年7—10月份在茂兰国家级自然保护区内建立1个1 hm2(100 m×100 m)的永久样地(25°18′25″N,107°57′48″E),该样地保存有相对完好的原生性常绿落叶阔叶混交林,人为干扰少,样地位于一个山峰的坡面上,地形复杂,坡度较大,岩石裸露率高,样地的最低点位于山脚,最高点接近山顶。使用森林罗盘仪(DQL-1,哈尔滨市光学仪器厂)建立样地,并将样地划分为100个10 m×10 m样方,再将样方划分为4个5 m×5 m调查单元,调查每个样方内所有胸径(DBH)≥1 cm的木本植物,记录其种名、个体相对坐标、胸径、树高、冠幅、生长状况以及岩石裸露率、小生境类型、坡度和土壤厚度等指标。对于DBH<1 cm的乔木幼树和灌木以及草本层植物,记录其种类组成与盖度等特征。
2.2 数据分析 2.2.1 物种选择与龄级划分样地调查共记录到DBH≥1 cm的木本植物199种,4281株活个体,隶属于63科140属。计测所有物种的重要值,计测方法和结果参见文献[24]。选取重要值较大的前30种乔木树种为研究对象(表 1),分析种群空间分布格局及其分形特征。同时,为分析不同龄级树种空间分布的分形特征变化,以径级代替龄级,将30种树种中的小乔木和大乔木树种分别按照如下方式进行龄级划分:(1)小乔木树种:1 cm ≤ DBH < 2.5 cm定为幼树;2.5 cm ≤ DBH < 10 cm为中树;DBH ≥ 10 cm为成树。(2)大乔木树种:1 cm ≤ DBH < 5 cm定为幼树;5 cm ≤ DBH < 15 cm为中树;DBH ≥ 15 cm为成树。虽然龄级和径级是不同的,但在相对一致的环境条件下同一树种龄级和径级对环境的反应规律具有一致性[31]。因此,在年龄难以准确确定情况下,多数学者采用径级代替龄级进行分析[3, 31]。
2.2.2 格局强度指数的计测测定种群分布格局的聚集强度,判断分布格局的类型。这里以100个10 m×10 m样方中的物种多度数据为基础,选取4种常用的格局强度指数来分析30种树种的种群分布格局类型。4种指数分别为:聚块性指数(m*/m)、格林指数(GI)、扩散系数(C)和负二项参数(K),其详细计测过程可参见有关文献[32]。其中,当聚块性指数m*/m < 1时判定为逼近均匀分布,m*/m = 1时为随机分布,m*/m > 1时为逼近聚集分布;格林指数GI值介于0—1之间,GI趋于0时表示逼近随机分布,趋于1时为逼近最大聚集分布;扩散系数C < 1时趋于均匀分布,C = 1趋于随机分布,C > 1趋于聚集分布;负二项参数K < 0时趋于均匀分布,K > 0时趋于聚集分布,当K→+∞(一般为8以上)则逼近随机分布。4种格局强度指数的计算结果可进行对比分析,以确保格局分析结果的可靠性。上述4种格局强度指数利用Excel自带的Visual Basic编辑器进行编程运算完成。
2.2.3 分形维数的计测分形维数是对分形体空间填充程度的反映,其中计盒维数和信息维数是常用于刻画种群空间分布特征的两种重要指标[15]。其中计盒维数揭示出种群个体对水平空间的占据程度,信息维数揭示格局强度的尺度变化程度以及个体分布的非均匀状况[33]。计盒维数的计算公式为:
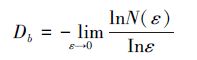
式中,Db为计盒维数,ε为对样地进行栅格化处理时的网格边长,N (ε)为对应于尺度ε的非空格子数。将样方边长100 m从2等分至50等分,所对应的尺度ε边长分别为50、33.33、25、…、2 m,统计每个尺度ε所对应的非空格子数N (ε),两者在双对数坐标下进行直线拟合,所得的直线拟合斜率的绝对值即为计盒维数的估计值。
在计盒维数的基础上,信息维数进一步考虑了每一非空格子中的个体数,并通过信息量公式给出尺度及其对应信息量间的幂律关系[13],其计算公式为:

选取30种树种中每种树木及其不同龄级(幼树、中树和成树)种群空间分布的计盒维数和信息维数,同时对不同种群各龄级间分形维数值的差异显著性进行多重比较(LSD法)。并且,利用Pearson相关分析(双尾检验)揭示分形维数(计盒维数和信息维数)与5种种群结构参数(多度、频度、平均高度、平均胸径和重要值)和4种格局强度指数间的相关关系。多重比较和Pearson相关分析在统计软件SPSS 17.0中完成。
3 结果与分析 3.1 种群的空间分布格局类型根据聚块性指数、格林指数、扩散系数和负二项参数综合判定茂兰1 hm2森林样地中30种树种的格局类型,结果表明28种树种的空间分布格局呈现聚集分布,仅皱叶海桐和油柿种群表现为随机分布(表 1)。格局强度指数虽难以反映格局类型随尺度变化的依赖关系,但也能准确量化不同树木聚集强度的变化,本研究结果与我们早期运用点格局方法甄别茂兰森林树种空间格局类型的结果基本一致[5]。茂兰中亚热带喀斯特森林树种以聚集格局为主,其格局的形成与所处的岩溶生境以及物种的生物和生态习性密切相关。
3.2 种群空间分布的分形维数计盒维数是采用网格覆盖方法得到的分形维数,反映了分形体占据面积随尺度变化的规律。因此,计盒维数揭示了种群对空间的占据程度和利用空间的能力。计盒维数的大小一般介于0—2之间,当值等于2时表明该物种已占据了整个水平空间。由表 1可见,30种树种种群在不同尺度与非空格子数(即种群占据的格子)的直线拟合达到了极显著相关水平,种群的空间分布格局具有明显的分形特征。30种乔木种群的Db值介于0.589—1.870之间,Db值较大的为天峨槭(1.870),其次为齿叶黄皮(1.767)、短序荚蒾(1.681)、青冈(1.647)和黄杞(1.618)等,其Db值>1.6。其中,天峨槭的Db值最大,接近2,表明该种在样地中对空间的占据能力最强,其次是齿叶黄皮和短序荚蒾。Db值较小的物种为油柿(0.589)、掌叶木(0.590)、小叶柿(0.768)、日本杜英(0.813)、毛萼红果树(0.891)和细叶青冈(0.916),其Db值<1,说明这些种群对空间占据以及利用能力较弱。
| 编号 No. | 种名 Species | 聚块性指数 Patchiness index | 格林指数 Green′ s index | 扩散系数 Dispersion index | 负二项参数 Negative binomial parameter | 格局类型 Pattern type | 计盒维数 Box-counting dimension | 信息维数 Information dimension | ||
| Db | 相关系数 Correlation coefficient | Di | 相关系数 Correlation coefficient | |||||||
| 1 | 圆果化香Platycarya longipes | 2.5272 | 0.0125 | 2.2371 | 0.6548 | 聚集 | 1.423 | 0.957* * | 1.307 | 0.941* * |
| 2 | 天峨槭Acer wangchii | 1.9210 | 0.0265 | 3.6248 | 1.0858 | 聚集 | 1.870 | 0.985* * | 1.711 | 0.960* * |
| 3 | 齿叶黄皮Clausena dunniana | 2.0609 | 0.0221 | 3.1854 | 0.9426 | 聚集 | 1.767 | 0.943* * | 1.669 | 0.968* * |
| 4 | 紫弹朴Celtis biondii | 1.3230 | 0.0029 | 1.2907 | 3.0961 | 聚集 | 1.065 | 0.909* * | 1.003 | 0.892* * |
| 5 | 青冈Cyclobalanopsis glauca | 1.9550 | 0.0279 | 3.7601 | 1.0471 | 聚集 | 1.647 | 0.937* * | 1.505 | 0.958* * |
| 6 | 短序荚蒾Viburnum brachybotryum | 2.6387 | 0.0356 | 4.5231 | 0.6103 | 聚集 | 1.681 | 0.981* * | 1.488 | 0.927* * |
| 7 | 长柱十大功劳Mahonia duclouxiana | 1.1309 | 0.0028 | 1.2814 | 7.6399 | 聚集 | 1.238 | 0.893* * | 1.048 | 0.876* * |
| 8 | 皱叶海桐Pittosporum crispulum | 1.0646 | 0.0011 | 1.1065 | 15.4900 | 随机 | 1.034 | 0.928* * | 0.959 | 0.872* * |
| 9 | 小花梾木Swida parviflora | 2.8720 | 0.0261 | 3.5834 | 0.5342 | 聚集 | 1.579 | 0.968* * | 1.386 | 0.950* * |
| 10 | 云贵鹅耳枥Carpinus pubescens | 2.4951 | 0.0103 | 2.0166 | 0.6689 | 聚集 | 1.083 | 0.954* * | 1.094 | 0.932* * |
| 11 | 香叶树Lindera communis | 1.8196 | 0.0100 | 1.9917 | 1.2201 | 聚集 | 1.122 | 0.911* * | 1.106 | 0.850* * |
| 12 | 小果润楠Machilus microcarpa | 5.1852 | 0.0444 | 5.3944 | 0.2389 | 聚集 | 1.591 | 0.994* * | 1.423 | 0.985* * |
| 13 | 山矾Symplocos sumuntia | 3.6913 | 0.0253 | 3.5029 | 0.3716 | 聚集 | 1.485 | 0.934* * | 1.369 | 0.945* * |
| 14 | 油柿Diospyros oleifera | 1.0380 | 0.0002 | 1.0232 | 26.3136 | 随机 | 0.589 | 0.877* * | 0.498 | 0.829* * |
| 15 | 粗丝木Gomphandra tetrandra | 3.1495 | 0.0267 | 3.6438 | 0.4652 | 聚集 | 1.583 | 0.921* * | 1.385 | 0.952* * |
| 16 | 栎叶罗伞Brassaiopsis quercifolia | 2.1367 | 0.0087 | 1.8639 | 0.8797 | 聚集 | 1.153 | 0.929* * | 1.102 | 0.911* * |
| 17 | 掌叶木Handeliodendron bodinieri | 1.8218 | 0.0022 | 1.2137 | 1.2168 | 聚集 | 0.590 | 0.872* * | 0.547 | 0.874* * |
| 18 | 枝花李榄Linociera ramiflora | 2.9516 | 0.0170 | 2.6784 | 0.5124 | 聚集 | 1.142 | 0.891* * | 1.110 | 0.931* * |
| 19 | 桂北木姜子Litsea subcoriacea | 4.0871 | 0.0243 | 3.4079 | 0.3239 | 聚集 | 1.351 | 0.934* * | 1.198 | 0.963* * |
| 20 | 黄杞Engelhardtia roxburghiana | 8.1903 | 0.0763 | 8.5498 | 0.1391 | 聚集 | 1.618 | 0.992* * | 1.462 | 0.989* * |
| 21 | 角叶槭Acer sycopseoides | 3.4917 | 0.0209 | 3.0682 | 0.4013 | 聚集 | 1.343 | 0.912* * | 1.143 | 0.928* * |
| 22 | 华南梾木Swida austrosinensis | 3.5607 | 0.0114 | 2.1267 | 0.3905 | 聚集 | 1.026 | 0.925* * | 0.929 | 0.913* * |
| 23 | 毛萼红果树Stranvaesia amphidoxa | 1.9401 | 0.0060 | 1.5923 | 1.0637 | 聚集 | 0.891 | 0.903* * | 0.875 | 0.906* * |
| 24 | 红翅槭Acer fabri | 6.4131 | 0.0257 | 3.5442 | 0.1847 | 聚集 | 1.221 | 0.966* * | 1.210 | 0.973* * |
| 25 | 贵州琼楠Beilschmiedia kweichowensis | 3.0388 | 0.0105 | 2.0398 | 0.4905 | 聚集 | 1.141 | 0.912* * | 1.151 | 0.926* * |
| 26 | 小叶柿Diospyros dumetorum | 1.4078 | 0.0021 | 1.208 | 2.4524 | 聚集 | 0.768 | 0.834* * | 0.684 | 0.893* * |
| 27 | 枇杷Eriobotrya japonica | 4.2269 | 0.0101 | 2.0003 | 0.3099 | 聚集 | 1.075 | 0.917* * | 1.020 | 0.937* * |
| 28 | 日本杜英Elaeocarpus japonicus | 2.7172 | 0.0052 | 1.5152 | 0.5824 | 聚集 | 0.813 | 0.895* * | 0.734 | 0.906* * |
| 29 | 细叶青冈Cyclobalanopsis myrsinaefolia | 3.3400 | 0.0080 | 1.7956 | 0.4273 | 聚集 | 0.916 | 0.911* * | 0.915 | 0.926* * |
| 30 | 川桂Cinnamomum wilsonii | 2.1839 | 0.0068 | 1.6748 | 0.8447 | 聚集 | 1.002 | 0.934* * | 0.906 | 0.927* * |
| * *P<0.01 | ||||||||||
信息维数表是计盒维数的推广,其不但考虑所取格子是“空”还是“非空”,而且考虑了不同的非空格子所提供的信息量大小,从而反映种群空间分布的非均匀性程度[34]。信息维数的大小可表征种群聚集格局的强度[15]。30个树种空间分布格局的信息维数计算结果见表 1,不同尺度与信息量的直线拟合效果均达到极显著水平。30种乔木种群的Di值介于0.498—1.711之间,Di值较大的为天峨槭(1.711)、齿叶黄皮(1.669)、青冈(1.505)、短序荚蒾(1.488)等;Di值较小的树种为油柿(0.498)、掌叶木(0.547)、小叶柿(0.684)、日本杜英(0.734),这说明天峨槭种群非均匀性强,聚集程度最高,齿叶黄皮聚集也较强,油柿种群格局的聚集程度最低。
3.3 不同龄级种群空间分布的分形维数变化对30种树种3个龄级空间分布的分形维数进行多重比较。从结果可看出,从幼树、中树到成树阶段,随着树龄的增加,30种树种的Db和Di平均值显著降低(图 1)。这一结果表明,随着树龄的增加,种群的空间占据能力显著减弱,个体分布趋于均匀,格局的强度明显降 低。此外,Db和Di值在各龄级内无显著差异,表明虽然Db和Di计测方法有差别,但均能较灵敏地量化种群空间分布的格局变化。随树龄的增加,种群往往由聚集分布逐渐转变为随机分布[35]。在本研究中,各龄级间分形维数的显著降低同样也印证了这一规律。

|
| 图1 喀斯特常绿落叶阔叶混交林30种树种不同龄级的分形维数 Fig.1 Fractal dimensions between different age stages of 30 tree species in the mixed evergreen-deciduous broad-leaved karst forest in Maolan |
2 种分形维数与5种种群结构参数和4种格局强度指数间的Pearson相关分析表明(表 2),种群多度和重要值与计盒维数和信息维数间均呈现极显著正相关关系,说明个体数越多、重要值越大的树种,即处于优势地位的树种,其分形维数值往往较高。因此,种群多度和重要值的变化与种群生态空间占据能力和聚集程度存在密切关系。而频度、平均高度和平均胸径与2种分形维数间无显著相关性。格林指数、扩散系数和负二项参数等指标均与分形维数存在极显著或显著的相关关系,而聚块性指数与分形维数间无显著相关关系。这一结果表明,分形维数的大小可以在一定程度上反映种群空间格局的变化,较大的分形维数值表明种群个体的聚集强度较高,而较小的分形维数则表明聚集强度相对较弱。此外,虽然频度和聚块性指标与分形维数间未达到统计学上的显著程度,但其P值均接近0.05,表明它们之间也存在一定的相关关系。
| 种群结构参数与格局指数 Population structure parameter and pattern index | 计盒维数 Box-counting dimension | 信息维数 Information dimension | ||
| 相关系数 Correlation coefficient | P | 相关系数 Correlation coefficient | P | |
| 多度 Abundance | 0.712 | < 0.01 | 0.681 | < 0.01 |
| 频度 Frequency | 0.351 | 0.057 | 0.321 | 0.083 |
| 平均高度 Mean height | -0.218 | > 0.05 | -0.205 | > 0.05 |
| 平均胸径 Mean DBH | -0.212 | > 0.05 | -0.201 | > 0.05 |
| 重要值 Important value | 0.546 | < 0.01 | 0.546 | < 0.01 |
| 聚块性指数 Patchiness index | 0.329 | 0.075 | 0.344 | 0.062 |
| 格林指数 Green′ s index | 0.729 | < 0.01 | 0.708 | < 0.01 |
| 扩散系数 Dispersion index | 0.729 | < 0.01 | 0.729 | < 0.01 |
| 负二项参数Negative binomial parameter | -0.387 | < 0.05 | -0.434 | < 0.05 |
聚集分布是森林生态系统中最为普遍的植物种群空间分布格局类型[2, 3, 4, 5]。例如,Condit等[2]检验了6块热带森林大样地中1768种树木的空间分布格局,发现大部分物种是聚集分布而非随机分布。Li等[8]和Wang等[9]发现我国鼎湖山的亚热带森林和长白山温带林树木的空间分布主要呈现聚集格局,同时随树龄的增加,聚集强度下降。笔者曾运用Condit等[2]提出的相对邻体密度系数(relative neighborhood density index)分析了茂兰喀斯特森林43种木本植物的空间分布,结果表明93.0%、81.3%和69.8%的树种分别在0—10 m、10—20 m和20—30 m空间尺度内表现为聚集分布[5]。本研究中,茂兰1 hm2喀斯特森林样地30种树种中,28种树种表现为聚集分布,仅2种为随机分布,这一结果进一步表明喀斯特森林树种的聚集分布特征。
生境异质性和扩散限制是导致植物种群聚集分布的两大主要原因[36]。茂兰喀斯特生境的岩石裸露高、地形起伏较大,微地形环境复杂,土壤分布极为不均,小生境异质性强。地形和生境的变化导致环境因子的空间变异,如光照、温度、水分、土壤厚度和养分等生态因子都随着地形或海拔的变化而存在差异[27, 37]。因此,这种复杂多样的生境变化结合不同植物的适合度(fitness),致使多数物种在不同生境斑块内维持生存[38]。如,圆果化香、齿叶黄皮和云贵鹅耳枥等多分布于样地中干旱瘠薄、海拔较高和多岩石的生境中,而短序荚蒾、小花梾木和山矾等树种在中低海拔区域分布较多。扩散限制是影响同种聚集的另一主要因素。本研究中的30种乔木树种,多数依靠重力传播种子,其种子初始扩散的范围以母树冠幅附近为主。此外,石槽、石沟和石缝等母树周围的岩溶小生境会阻止种子在地面的进一步扩散。局部扩散导致多数种子落在母株附近,即使天敌(食草动物、昆虫和病原菌)引起的距离和密度制约效应会降低母树周围种子的萌发率和幼苗的存活率[39, 40],但大部分新增补的个体仍会聚集在母树周围,导致种群的聚集分布[41]。
本研究认为,喀斯特森林树种的聚集分布与生境存在密切关联,多数树种表现出生境偏好性,同时扩散限制也助推了聚集格局的形成。生境异质性和扩散限制分别是生态位分化理论和中性理论所强调的植物群落构建方式。因此,生态位和中性过程在喀斯特森林树种空间分布与物种共存方面均具有重要贡献,但2种过程孰轻孰重还有待进一步研究。
4.2 树种空间分布的分形特征计盒维数和信息维数分别从种群的空间占据能力及种群个体分布的非均匀性的角度来揭示种群分布格局的变化特征,2种分形维数相辅相承相互补充,为深入理解种群格局提供了丰富信息[34]。张建亮等[42]研究表明,广西木论喀斯特森林中铁榄(Sinosideroxylon pedunculatum)种群的计盒维数与生境关系密切,铁榄种群在石面、石沟、石缝、石槽及其相互间的组合的生境中具有极强的生存适合度,因而对生境的占据能力最强。本研究同样发现,种群对空间环境的占据和利用能力大小、种群个体的分布不均匀和格局的聚集程度与所处的喀斯特生境条件及种群生物和生态习性密切相关。
天峨槭和齿叶黄皮种群的个体数分别为285株和206株,在群落中处于优势地位,适应多种小生境类型,特别对岩石裸露较高生境的占据及利用能力较强。因此,与其它种群相比,天峨槭和齿叶黄皮对环境的占据与利用能力最强,生长状况和更新状况较佳。短序荚蒾和青冈种群在群落中也处于优势地位,种群密度大,个体数分别为215株和289株,对土层较多和多岩石的生境的占据及利用能力较强,其计盒维数值较大。此外,天峨槭、齿叶黄皮、短序荚蒾和青冈等树种的果实相对较重且结实量大,扩散限制使得种子萌发成幼苗以及生长成为幼树、小树后在母树周围形成聚集分布,因此信息维数较大。而油柿和掌叶木种群的计盒维数和信息维数均较小,对空间环境的占据与利用能力弱,受到其它优势种的竞争排斥,个体数较少(掌叶木仅有26株个体),多分布在岩石裸露多的严酷生境,而且这些生境的空间分布不连续,个体之间聚集程度相对较低,在群落中处于劣势伴生地位。
从上述结果看,计盒维数和信息维数较为准确地量化了30种树种的种群空间占据能力和聚集(非均匀分布)强度的变化。分形维数值越大,表征种群对空间环境的占据利用能力越强,个体空间聚集强度越强,在群落中往往处于优势地位;反之则表明其对环境的占据能力弱,个体聚集程度越低,往往处于伴生地位。
4.3 不同龄级种群空间分布的分形维数变化树种在不同生长阶段会表现出不同的空间格局,这与竞争、Janzen-Connell效应、干扰以及环境的变化有密切关系[5]。通过对30种树种不同龄级空间分布的分形维数分析,表明从幼树、中树到成树阶段的计盒维数和信息维数值呈显著降低趋势。许多研究均发现随树龄的增大,聚集强度会下降[3, 8, 9],本研究中的分形维数变化也可清晰地表明这一现象。
一般情况下,以母株为中心的繁殖方式使幼树呈现聚集分布,但随着植物个体径级的增大,其对环境资源,如光、水、营养元素和空间等的需求也随之加剧,导致种内和种间相互作用增强而造成竞争性密度制约死亡效应。同时,幼年阶段的高密度植株也可能会增加天敌的种类和多度,从而也会造成非竞争性(天敌引起)的密度制约死亡效应[39, 40]。竞争性和非竞争性因素造成的密度制约性死亡效应是导致种群随径级的增加而聚集强度逐渐减弱的主要原因[43, 44]。从幼树到成树的种群个体数和聚集强度逐渐降低,其分形维数值也相应减小。因此,分形维数在不同龄级间的变化也可在一定程度上反映出种群的更新动态。中树和大树聚集强度的降低有利于幼树获得足够的环境资源,种群聚集强度的变化是种群的一种生存策略或适应机制[45]。
4.4 分形维数与格局强度指数的相关关系种群空间分布的分形维数是否可以表征其格局强度和多度等生态属性,少数学者做了相关研究。如,向悟生等[34]研究表明,负二项参数、扩散系数等指标测定的聚集强度与信息维数存在显著的相关关系,较大的信息维数表明种群的聚集强度较高,而较小的信息维数则反映出较弱的聚集强度。而王瑞波等[46]将格局强度指数与计盒维数相结合分析了2种百合(Lilium sp.)种群的空间分布格局,结果表明2种格局分析方法的结果基本一致,说明计盒维数可以很好地反映种群的空间分布格局和种群动态。本研究中,除了聚块性指数外,计盒维数和信息维数值分别与3种格局强度指数(格林指数、扩散系数和负二项参数)间呈现显著或极显著相关性,表明计盒维数和信息维数值均能够较为客观地表征茂兰喀斯特森林样地中种群分布的空间占据能力和聚集程度。因此,综合上述结果表明,计盒维数和信息维数2种分形分维方法结合格局强度指数在量化种群空间分布格局方面具有较高的准确性和可靠性。
| [1] | Dale M R T. Spatial Pattern Analysis in Plant Ecology. Cambridge: Cambridge University Press, 1999. |
| [2] | Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470):1414-1418. |
| [3] | He F L, Legendre P, Lafrankie J V. Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science, 1997, 8(1):105-114. |
| [4] | Wiegand T, Huth A, Martínez I. Recruitment in tropical tree species: revealing complex spatial patterns. The American Naturalist, 2009, 174(4): 106-140. |
| [5] | Zhang Z H, Hu G, Zhu J D, Ni J. Aggregated spatial distributions of species in a subtropical karst forest, southwestern China. Journal of Plant Ecology, 2013, 6(2): 131-140. |
| [6] | Wiegand T, Moloney K A. Rings, circles, and null-models for point pattern analysis in ecology. Oikos, 2004, 104(2): 209-229. |
| [7] | Nishimura S, Yoneda T, Fujii S, Mukhtara E, Kanzakia M. Spatial patterns and habitat associations of Fagaceae in a hill dipterocarp forest in Ulu Gadut, West Sumatra. Journal of Tropical Ecology, 2008, 24(5): 535-50. |
| [8] | Li L, Huang Z L, Ye W H, Cao H L, Wei S G, Wang Z G, Lian J Y, Sun I F, Ma K P, He F L. Spatial distributions of tree species in a subtropical forest of China. Oikos, 2009, 118(4): 495-502. |
| [9] | Wang X G, Ye J, Li B H, Zhang J, Lin F, Hao Z Q. Spatial distributions of species in an old-growth temperate forest, northeastern China. Canadian Journal of Forest Research, 2010, 40(6): 1011-1019. |
| [10] | Mandelbrot B B. The Fractal Geometry of Nature. New York: W.H. Freeman & Co Ltd, 1983. |
| [11] | Vedyushkin M A. Fractal properties of forest spatial structure. Vegetatio, 1994, 113(1): 65-70. |
| [12] | Brown J H, Grupta V K, Li B L, Milne B T, Restrepo C, West G B. The fractal nature of nature: power laws, ecological complexity and biodiversity. Philosophical Transactions of the Royal Society of London B, 2002, 357(1421): 619-626. |
| [13] | Alados C L, Pueyo Y, Giner M L, Navarro T, Escos J, Barroso F, Cabezudo B, Emlen J M. Quantitative characterization of the regressive ecological succession by fractal analysis of plant spatial patterns. Ecological Modelling, 2003, 63(1-2): 1-17. |
| [14] | Halley J M, Hartley S, Kallimanis A S, Kunin W E, Lennon J J, Sgardelis S P. Uses and abuses of fractal methodology in ecology. Ecology Letters, 2004, 7(3): 254-271. |
| [15] | 马克明, 祖元刚. 植被格局的分形特征. 植物生态学报, 2000, 24(1): 111-117. |
| [16] | Nalakarn P, Tang I M, Triampo W. Fractal studies on the spatial patterns of trees: A case study of Khao Yai National Park, Thailand. ScienceAsia, 2008, 34(4): 409-415. |
| [17] | 马克明, 张喜军, 陈继红, 祖元刚. 东北羊草草原群落格局的分数维理论研究 // 辛厚文. 分形理论及其应用. 合肥: 中国科学技术大学出版社, 1993: 258-264. |
| [18] | 祖元刚, 马克明. 分形理论与生态学 // 李博. 现代生态学讲座. 北京: 科学出版社, 1995: 65-72. |
| [19] | 张文辉, 祖元刚, 马克明. 裂叶沙参与泡沙参种群分布格局分形特征的分析. 植物生态学报, 1999, 23(1): 31-39. |
| [20] | 梁士楚, 董鸣, 王伯荪, 张炜银. 红树植物木榄种群分布格局关联维数的研究. 海洋科学, 2003, 27(6): 51-54. |
| [21] | 宋于洋, 李园园, 张文辉. 基于Ripley的K(r)函数和分形维数的梭梭种群空间格局. 应用生态学报, 2010, 21(4): 827-835. |
| [22] | 玉屏, 兰洪波, 冉景丞, 姚正明. 茂兰自然保护区生物多样性现状及保护对策. 现代农业科技, 2011, (15): 233-234, 236-236. |
| [23] | 周政贤. 茂兰喀斯特森林科学考察集. 贵阳: 贵州人民出版社, 1987: 1-23. |
| [24] | Zhang Z H, Hu G, Zhu J D, Ni J. Stand structure, woody species richness and composition of subtropical karst forests in Maolan, south-west China. Journal of Tropical Forest Science, 2012, 24(2): 498-506. |
| [25] | 杜道林, 刘玉成, 苏杰. 茂兰喀斯特山地广东松种群结构和动态初步研究. 植物生态学报, 1996, 20(2): 159-166. |
| [26] | Zhang Z H, Hu G, Zhu J D, Luo D H, Ni J. Spatial patterns and interspecific associations of the dominant tree species in two old-growth karst forests, SW China. Ecological Research, 2010, 25(6): 1151-1160. |
| [27] | 张忠华, 胡刚, 祝介东, 倪健. 喀斯特森林土壤养分的空间异质性及其对树种分布的影响. 植物生态学报, 2011, 35(10): 1038-1049. |
| [28] | Zhang Z H, Hu G, Ni J. Effects of topographical and edaphic factors on the distribution of plant communities in two subtropical karst forests, southwestern China. Journal of Mountain Science, 2013, 10(1): 95-104. |
| [29] | 朱守谦. 喀斯特森林生态研究(Ⅰ). 贵阳: 贵州科技出版社, 1993: 1-11. |
| [30] | 喻理飞, 朱守谦, 叶镜中. 退化喀斯特森林自然恢复评价研究. 林业科学, 2000, 36(6): 12-19. |
| [31] | 张金屯, 孟东平. 芦芽山华北落叶松林不同龄级立木的点格局分析. 生态学报, 2004, 24(1): 35-40. |
| [32] | 江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992. |
| [33] | 祖元刚, 马克明, 张喜军. 植被空间异质性的分形分析方法. 生态学报, 1997, 17(3): 333-337. |
| [34] | 向悟生, 李先琨, 苏宗明, 欧祖兰, 陆树华. 元宝山南方红豆杉克隆种群分布格局的分形特征. 植物生态学报, 2007, 31(4): 568-575. |
| [35] | Gavrikov V, Stoyan D. The use of marked point processes in ecological and environmental forest studies. Environmental and Ecological Statistics, 1995, 2(4): 331-344. |
| [36] | Shen G C, He F L, Waagepetersen R, Sun I F, Hao Z Q, Chen Z S, Yu M J. Quantifying effects of habitat heterogeneity and other clustering processes on spatial distributions of tree species. Ecology, 2013, 94(11): 2436-2443. |
| [37] | 周运超, 潘根兴. 茂兰森林生态系统对岩溶环境的适应与调节. 中国岩溶, 2001, 20(1): 47-52. |
| [38] | Queenborough S A, Burslem D F R P, Garwood N C, Valencia R. Habitat niche partitioning by 16 species of Myristicaceae in Amazonian Ecuador. Plant Ecology, 2007, 192(2):193-207. |
| [39] | Janzen D H. Herbivores and the number of tree species in tropical forests. The American Naturalist, 1970, 104(940): 501-528. |
| [40] | Connell J H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees // den Boer P J, Gradwell G R. Dynamics of Populations. Netherlands: Centre for Agricultural Publishing and Documentation, 1971: 298-312. |
| [41] | Hubbell S P. The Unified Neutral Theory of Biodiversity and Biogeography. Princeton: Princeton University Press, 2001. |
| [42] | 张建亮, 李先琨, 吕仕洪, 叶铎, 钟军弟, 焦继飞, 尤业明. 广西木论自然保护区铁榄群落主要乔木种群格局的分形特征. 武汉植物学研究, 2009, 27(6): 622-628. |
| [43] | Comita L S, Muller-Landau H C, Aguilar S, Hubbell S P. Asymmetric density dependence shapes species abundances in a tropical tree community. Science, 2000, 329(5989): 330-332. |
| [44] | Piao T F, Comita L S, Jin G Z, Kim J H. Density dependence across multiple life stages in a temperate old-growth forest of northeast China. Oecologia, 2013, 172(1): 207-217. |
| [45] | Hubbell S P. Neutral theory and the evolution of ecological equivalence. Ecology, 2006, 87(6): 1387-1398. |
| [46] | 王瑞波, 张燕平, 胡世俊, 何平, 张春平, 邓洪平. 两种百合种群空间分布格局对高温干旱气候的响应.林业科学研究, 2009, 22(2): 249-255. |