 2015, Vol. 35
2015, Vol. 35文章信息
- 姜会超, 陈海刚, 宋秀凯, 刘宁, 何健龙, 程玲, 王月霞
- JIANG Huichao, CHEN Haigang, SONG Xiukai, LIU Ning, HE Jianlong, CHENG Ling, WANG Yuexia
- 莱州湾金城海域浮游动物群落结构及与环境因子的关系
- Zooplankton community structure in Jincheng area of Laizhou Bay and its relationship with environmental factors
- 生态学报, 2015, 35(22): 7308-7319
- Acta Ecologica Sinica, 2015, 35(22): 7308-7319
- http://dx.doi.org/10.5846/stxb201403260565
-
文章历史
- 收稿日期: 2014-03-26
- 网络出版日期: 2015-04-20






2. 广东省渔业生态环境重点实验室, 广州 510300;
3. 烟台市芝罘区渔业技术推广站, 烟台 264001
2. Key Laboratory of Fishery Ecology and Environment, Guangdong Province, Guangzhou 510300, China;
3. Zhifu Fishery Technical Extension Station, Yantai 264001, China
莱州湾地处渤海南岸,内有黄河、小清河、界河等多条河流注入,这些河流带来大量营养物质的同时,也对莱州湾造成了一定程度的污染,加上对海洋资源的过度捕捞,莱州湾的海洋生态环境不断恶化。近年来针对莱州湾海域的生态调查多集中在渔业资源上,对浮游动物的调查较少见有报道[1, 2, 3, 4]。浮游动物是具有有限的活动能力的异养生物,一方面通过捕食作用控制浮游植物数量,另一方面作为鱼类等高层营养者的饵料,其数量变化可以直接影响鱼类等的资源量,在海洋生态系统结构和功能中起重要的调控作用[5, 6]。摸清浮游动物的群落结构特征及其变化规律对渔情预报及生态学等方面均具有重要意义。
近岸海域中浮游动物种类繁多、环境因子复杂多变,水域中的浮游动物群落同时受周围多种环境因素(如物理、化学和生物等)的影响,其群落结构与动态是多个环境因子在时间和空间尺度上综合作用的结果[7]。在以往的研究中人们多采用简单的线性模型来处理浮游动物与单个理化因子(物理、化学等)之间的关系。但由于近岸海域环境条件复杂多变,这种线性相关分析从单一的环境因子去讨论,难以给出全面的结果[8]。近年来,典范对应分析 (CCA) 因具有能同时结合多个环境因子分析生物与环境因子间的复杂关系、信息包含量大、结果明确直观等优点而得到越来越多国外学者的青睐,并在生态学领域中得到广泛运用[5, 9, 10]。本文选取莱州湾重点贝类养殖区-金城海域,在2009—2010年进行了长达两年共计20个航次的浮游动物调查,并结合理化因子数据,采用典范对应分析(CCA)技术对莱州湾金城海域近岸浮游动物种类组成、群落结构、优势种更替和数量分布及其与环境理化因子的关系进行研究,旨在较全面、系统地阐明近岸浮游动物的群落结构特征,探讨浮游动物群落演替规律及其与环境理化因子的关系,并为深入研究渤海浮游动物的长期变化规律、海洋生态系统动态变化机制及生态环境保护措施的制订等提供基础资料和理论依据。
1 材料与方法 1.1 采样区域、时间与方法调查海区为莱州湾金城海域,共设15个采样站位(J1—J15),水深范围为3.5 m—13.0 m,由南向北逐渐加深。于2009年3—12月和2010年3—12月对15采样站位进行两年共20个月(航次)的水质及浮游动物调查,使用船只为当地渔民渔船。
浮游动物采集方法及标本处理等均按《海洋调查规范》[11]进行,用浅I浮游生物网自底至表层作垂直拖网采集,样品经5%甲醛溶液固定后实验室进行分类、鉴定、计数和统计(不包括夜光虫)。各监测站位同步调查水温(WT)、盐度(Salinity)、透明度(Transparency)、化学耗氧量(COD)、pH值、叶绿素(Chla) 及总氮(TN)总磷(TP)等环境参数,所有操作均按照《海洋监测规范》[12]进行。
1.2 数据分析浮游动物丰度以每立方米出现的个体数表示(个/m3)。
优势种的优势度:
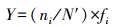
式中,ni 为第i种的丰度,fi为该种在各站位中出现的频率,N′为总丰度。根据种类优势度公式计算各种生物的优势度,将Y > 0.02的生物定为优势种。
物种多样性指数的计算采用Shannon-Winner指数:

物种丰度指数采用Margalef指数(dMa)计算公式:
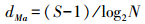
均匀度指数采用Pielou指数计算公式:
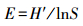
式中,N为采集样品中所有物种的总个体数,S为样品中的种类总数,Pi为第i种的个体数与样品中的总个体数的比值。
多元统计方法包括聚类分析和典范对应分析(CCA)等,均是基于种类数据和环境数据集合的排序方法,已逐步应用于浮游生物群落结构分析中。在数值分析前,对数据进行标准化处理。选择出现频率大于15%的种类。环境参数数据库包括8个环境指标,分别为水温、盐度、pH值、透明度、COD、TN、TP、Chla。对种类数据和环境参数采用对数log(x+1)转换。典范对应分析(CCA,canonical correspondence analysis)是探讨浮游动物群落与环境因子之间对应关系的一种方法,通过直接梯度分析,可将物种、样点及环境指标之间的关系反映在坐标轴上,直观地反映生物与环境之间的相关程度。
全部数据的统计分析用SPSS 17.0软件完成,群落结构聚类分析使用PRIMER 6.0软件,典范对应分析采用CANOCO4.5软件完成。
2 结果 2.1 环境因子2009及2010年水质理化指标结果如表1所示,2009年水质透明度要略高于2010年,年均透明度分别为2.95 m和2.57 m,2009年9月达到最高值(4.33±0.2)m,2010年12月达到最低值(1.51±0.06)m。pH值在2010年4月达到最高值(8.33±0),同年12月降低到最低值(7.84±0.01),2009年与2010年的年均pH值分别为8.12、8.11。水温在2009年8月达到最高值(27.32±0.04)℃,同年12月降到最低值(2.24 ±0.18)℃,2009年与2010年的年均水温分别为15.67 ℃、16.06 ℃。COD在2009年10月达到最高值(1.67±0.07)mg/L,在2010年4月降到最低值(1.12±0.05)mg/L,其年均COD值分别为1.33、1.29 mg/L。盐度在2010年3月达到最高值(31.9 ±0.01),2009年12月降到最低值(29.71±0.05),2009年与2010年的年均盐度分别为30.37、30.82。Chla在2009年4月达到最低值(0.88±0.24)μg/L,同年12月达到最高值(4.19±1.36)μg/L。2009年与2010年的年均Chla含量分别为2.14、2.27 μg/L。总氮与总磷两种营养盐在2009年的波动情况比较剧烈,而在2010年的各月份之间则无明显波动。从两年的水质监测数据看,除总氮与总磷外,其他各项指标在两年之间并无明显差异,且各指标在年际间的变化趋势也较为相似。
|
年份 Years | 月份 Months | 透明度/m Transparency | pH | 水温/℃ Water temperature | 化学耗氧量/(mg/L) Chemical oxygen demand | 盐度 Salinity | 叶绿素/(μg/L) Chl a | 总氮/(mg/L) Total nitrogen | 总磷/(mg/L) Total phosphorus |
| 2009 | 03 | 3.08±0.11 | 8.18±0.01 | 3.14 ±0.04 | 1.22±0.03 | 30.63±0.02 | 0.89±0.18 | 0.67±0.03 | 0.01±0.00 |
| 04 | 2.84±0.10 | 8.16±0.01 | 6.55 ±0.17 | 1.12±0.05 | 30.51±0.03 | 0.88±0.24 | 1.01±0.16 | 0.03±0.00 | |
| 05 | 4.17±0.17 | 7.88±0.02 | 16.93±0.22 | 1.65±0.02 | 29.78±0.04 | 1.46±0.18 | 0.63±0.05 | 0.02±0.00 | |
| 06 | 2.83±0.22 | 8.17±0.01 | 25.25±0.19 | 1.45±0.04 | 30.98±0.01 | 2.77±0.36 | 0.51±0.05 | 0.01±0.00 | |
| 07 | 2.55±0.14 | 8.14±0.02 | 26.10±0.08 | 1.44±0.07 | 30.34±0.02 | 1.74±0.37 | 0.68±0.08 | 0.01±0.00 | |
| 08 | 2.73±0.10 | 8.12±0.01 | 27.32±0.04 | 1.23±0.04 | 30.73±0.05 | 2.03±0.12 | 0.32±0.02 | 0.03±0.00 | |
| 09 | 4.33±0.20 | 8.17±0.02 | 23.35±0.13 | 1.21±0.05 | 30.69±0.02 | 1.97±0.42 | 1.05±0.07 | 0.06±0.00 | |
| 10 | 2.07±0.06 | 8.24±0.04 | 19.33±0.10 | 1.67±0.07 | 30.34±0.19 | 3.14±0.38 | 0.75±0.06 | 0.06±0.00 | |
| 11 | 2.80±0.11 | 8.13±0.02 | 6.44 ±0.18 | 1.19±0.03 | 30.03±0.11 | 2.32±0.19 | 0.50±0.01 | 0.01±0.00 | |
| 12 | 2.10±0.11 | 8.02±0.00 | 2.24 ±0.18 | 1.15±0.03 | 29.71±0.05 | 4.19±1.36 | 0.65±0.03 | 0.02±0.00 | |
| 2010 | 03 | 2.35±0.05 | 8.16±0.01 | 4.01 ±0.06 | 1.36±0.03 | 31.20±0.01 | 2.40±0.26 | 0.62±0.01 | 0.03±0.00 |
| 04 | 2.13±0.40 | 8.33±0.00 | 7.71 ±0.07 | 1.12±0.04 | 31.14±0.05 | 2.53±0.33 | 0.62±0.01 | 0.03±0.00 | |
| 05 | 2.41±0.41 | 8.19±0.00 | 15.26±0.06 | 1.19±0.04 | 31.90±0.01 | 3.59±0.19 | 0.63±0.01 | 0.03±0.00 | |
| 06 | 3.23±0.10 | 8.11±0.02 | 24.55±0.23 | 1.38±0.04 | 31.19±0.01 | 2.17±0.26 | 0.66±0.01 | 0.03±0.00 | |
| 07 | 2.43±0.07 | 8.10±0.01 | 25.89±0.08 | 1.56±0.06 | 31.20±0.01 | 1.69±0.10 | 0.73±0.01 | 0.04±0.00 | |
| 08 | 2.51±0.07 | 8.04±0.01 | 26.65±0.05 | 1.27±0.02 | 31.04±0.05 | 2.11±0.16 | 0.70±0.05 | 0.04±0.00 | |
| 09 | 2.85±0.08 | 8.11±0.03 | 25.83±0.09 | 1.29±0.05 | 30.38±0.14 | 2.30±0.13 | 0.64±0.01 | 0.04±0.00 | |
| 10 | 3.05±0.17 | 8.07±0.03 | 16.79±0.12 | 1.25±0.02 | 30.51±0.09 | 2.03±0.18 | 0.65±0.01 | 0.04±0.00 | |
| 11 | 3.27±0.08 | 8.18±0.02 | 10.40±0.13 | 1.33±0.03 | 29.88±0.12 | 1.34±0.17 | 0.77±0.01 | 0.02±0.00 | |
| 12 | 1.51±0.06 | 7.84±0.01 | 3.51 ±0.09 | 1.17±0.02 | 29.72±0.06 | 2.49±0.10 | 1.38±0.02 | 0.02±0.00 |
本调查共鉴定出75种不同种类的浮游动物,分属12大类。其中以桡足类和浮游幼虫居多,分别鉴定出22种、20类,占总种数29.3%、26.7%。其次为水螅水母类,鉴定出11种,占总种数14.7%。端足类、被囊动物、腹足类和栉水母类分别鉴定出6种、4种、4种、2种,占总种数8%、5.3%、5.3%、2.7%,其它枝角类、毛颚动物、等足类、糠虾类、磷虾类以及钵水母类各1种。虽然毛颚动物只鉴定到一种强壮箭虫,但其几乎在每个站位均有分布,占总数量的24.2%,为绝对优势种(表2)。
|
物种 Species | 缩略词 Abbreviate | 丰度/(个/m3) Average abundance | 出现频率 Occurrence frequency | 优势度 Dominant |
| 毛颚动物Chaetognatha | ||||
| 强壮箭虫Sagitta crassa | SCRA | 75.042 | 1 | 0.242 |
| 水螅水母类Hydromedusa | ||||
| 贝氏真囊水母Euphysora bigelowi | EBIG | 1.401 | 0.1 | <0.02 |
| 八斑芮氏水母Rathkea octopunctata | ROCT | 9.494 | 0.15 | <0.02 |
| 鲍氏水母Bougainvillia muscus | BAUT | 1.114 | 0.05 | <0.02 |
| 带玛拉水母Malagazzia taeniogonia | PTAE | 0.009 | 0.1 | <0.02 |
| 两手鲍氏水母Bougainvillia bitentaculata | BBIT | 0.007 | 0.05 | <0.02 |
| 卡玛拉水母Malagazzia carolinae | PVIR | 0.084 | 0.05 | <0.02 |
| 单囊美螅水母Clytia folleata | PDIS | 1.935 | 0.25 | <0.02 |
| 囊状全水母Catablema vesicarium | TVES | 0.010 | 0.05 | <0.02 |
| 双叉薮枝螅水母Obelia dichotoma | ODIC | 0.022 | 0.05 | <0.02 |
| 薮枝螅水母Obelia sp. | OBSP | 1.341 | 0.55 | <0.02 |
| 水螅水母类Hydromedusae | HYSP | 0.644 | 0.5 | <0.02 |
| 钵水母类Scyphozoa | ||||
| 海月水母Aurelia aurita | AAUR | 0.010 | 0.05 | <0.02 |
| 栉水母类Ctenophora | ||||
| 刺胞栉水母Euchlora rubra | ERUB | 0.405 | 0.05 | <0.02 |
| 球形侧腕水母Pleurobrachia globosa | PGLO | 4.303 | 0.25 | <0.02 |
| 桡足类Copepoda | ||||
| 背针胸刺水蚤Centropages dorsispinatus | CDOR | 1.762 | 0.4 | <0.02 |
| 短角长腹剑水蚤Oithona brevicornis | OBRE | 0.776 | 0.55 | <0.02 |
| 海洋伪镖水蚤Pseudodiaptomus marinus | PMAR | 0.108 | 0.25 | <0.02 |
| 巨大怪水蚤Monstrilla grandis | MGRA | 0.012 | 0.2 | <0.02 |
| 克氏纺锤水蚤Acartia clausi | AGIE | 3.741 | 0.4 | <0.02 |
| 孔雀唇角水蚤Labidocera pavo | LPAV | 0.050 | 0.1 | <0.02 |
| 墨氏胸刺水蚤Centropages mcmurrichi | CMCM | 46.313 | 0.45 | 0.067 |
| 拟长腹剑水蚤Oithona similis | OSIM | 0.881 | 0.55 | <0.02 |
| 挪威小毛猛水蚤Microsetella norvegica | MNOR | 0.172 | 0.4 | <0.02 |
| 钳状歪水蚤Tortanus forcipatus | TFOR | 0.028 | 0.1 | <0.02 |
| 强额拟哲水蚤Parvocalanus crassirostris | PCRA | 0.453 | 0.3 | <0.02 |
| 近缘大眼剑水蚤Corycaeus affinis | CAFF | 0.562 | 0.65 | <0.02 |
| 瘦尾歪水蚤Tortanus gracilis | TGRA | 0.016 | 0.05 | <0.02 |
| 洪氏唇角水蚤Labidocera rotunda | LROT | 0.933 | 0.55 | <0.02 |
| 洪氏纺锤水蚤Acartia hongi | ABIF | 19.057 | 0.35 | 0.022 |
| 太平洋纺锤水蚤Acartia pacifica | APAC | 1.119 | 0.4 | <0.02 |
| 太平洋真宽水蚤Eurytemora pacifica | EPAC | 21.169 | 0.25 | <0.02 |
| 汤氏长足水蚤Calanopia thompsoni | CTHO | 4.392 | 0.6 | <0.02 |
| 小拟哲水蚤Paracalanus parvus | PPAR | 4.298 | 0.9 | <0.02 |
| 真刺唇角水蚤Labidocera euchaeta | LEUC | 0.883 | 0.5 | <0.02 |
| 中华哲水蚤Calanus sinicus | CSIN | 42.477 | 0.95 | 0.130 |
| 左突唇角水蚤Labidocera sinilobata | LSIN | 0.018 | 0.05 | <0.02 |
| 枝角类Cladocera | ||||
| 鸟喙尖头溞Penilia avirostris | PAVI | 5.936 | 0.25 | <0.02 |
| 浮游幼虫Pelagic larva | ||||
| 阿利玛幼体Alima larva | ALIL | 0.391 | 0.35 | <0.02 |
| 桡足类幼体Copepodite larva | COPL | 2.086 | 0.7 | <0.02 |
| 磁蟹溞状幼体Porcellana larva | PORL | 0.728 | 0.5 | <0.02 |
| 长尾类幼体Maeruran larva | MAEL | 14.027 | 0.75 | 0.034 |
| 短尾类幼体Brachyura larva | BRAL | 6.952 | 0.5 | <0.02 |
| 多毛类幼体Polychaete larvae | POLL | 1.175 | 0.85 | <0.02 |
| 海参耳状幼体Holothuroidea larva | HOLL | 0.168 | 0.05 | <0.02 |
| 海胆长腕幼虫Echinoidea larva | ECHL | 10.405 | 0.3 | <0.02 |
| 海蛇尾长腕幼虫Ophiuroidea larva | OPHL | 0.118 | 0.3 | <0.02 |
| 海星纲羽腕幼体Asteroidea larva | ASTL | 0.283 | 0.05 | <0.02 |
| 舌贝幼虫Lingula larva | LINL | 0.035 | 0.05 | <0.02 |
| 双壳类壳顶幼虫Bivalvia larva | BIVL | 2.179 | 0.85 | <0.02 |
| 藤壶六肢幼虫Balanidae sp nauplius larva | BALL | 0.542 | 0.5 | <0.02 |
| 幼螺Nail | NAIL | 0.295 | 0.4 | <0.02 |
| 鱼卵Fish egg | FISE | 0.445 | 0.55 | <0.02 |
| 仔稚鱼Fish larva | FISL | 0.471 | 0.5 | <0.02 |
| 枝角类幼体Cladocera larva | CLAL | 0.026 | 0.15 | <0.02 |
| 帚毛虫幼体Actinotrocha larva | ACTL | 0.089 | 0.2 | <0.02 |
| 珠形幼虫Arachnactis larva | ARAL | 0.120 | 0.1 | <0.02 |
| 碟状体Ephyra | EPHY | 0.003 | 0.05 | <0.02 |
| 被囊动物Tunicate | ||||
| 长尾住囊虫Oikopleura longicauda | OLON | 0.021 | 0.1 | <0.02 |
| 红住囊虫Oikopleura rufescens | ORUF | 3.851 | 0.05 | <0.02 |
| 异体住囊虫Oikopleura dioica | ODIO | 2.131 | 0.35 | <0.02 |
| 软拟海樽Dolioletta gegenbauri | DGEG | 2.208 | 0.05 | <0.02 |
| 端足类Amphipoda | ||||
| 端足类Amphipod | AMPH | 0.023 | 0.05 | <0.02 |
| 钩虾Gammarid | GAMM | 0.930 | 0.4 | <0.02 |
| 螺赢蜚Corophium sp. | CORO | 0.058 | 0.15 | <0.02 |
| 麦杆虫Caprellid | SSHR | 1.135 | 0.35 | <0.02 |
| 内海拟钩虾Gammaropsis utinomii | GUTI | 2.945 | 0.2 | <0.02 |
| 细足法Themisto gracilipes | TGRA | 0.157 | 0.25 | <0.02 |
| 腹足类Gastropoda | ||||
| 腹足类Gastropoda | GAST | 4.126 | 0.3 | <0.02 |
| 泥螺Bullacta exarata | BEXA | 0.002 | 0.05 | <0.02 |
| 翼足螺Pteropod | PTER | 0.010 | 0.05 | <0.02 |
| 马蹄螺Limacina trochiformis | LTRO | 0.026 | 0.1 | <0.02 |
| 等足类Isopoda | ||||
| 等足类Isopoda | ISOP | 1.002 | 0.5 | <0.02 |
| 糠虾类Mysidacea | ||||
| 糠虾Mysidacea | MYSI | 0.006 | 0.05 | <0.02 |
| 磷虾类Euphausiacea | ||||
| 磷虾Euphausid | EUPH | 0.003 | 0.05 | <0.02 |
如图1所示,生物丰度变化呈现出比较明显地季节变化规律,2009年与2010年的数据显示,3—4月份生物丰度均比较低,到5月份迅速升高达到一年当中的最高值(2009年951.65个/m3,2010年1348.14个/m3),从6月到12月,生物密度则基本表现出依次下降趋势,2009年与2010年浮游动物生物密度的总体变化趋势较为相似,年均生物丰度分别为310.81个/m3和314.57个/m3。

|
| 图 1 2009—2010年浮游动物群落参数(生物密度、生物量、种类数、丰富度指数、多样性指数、均匀度指数)动态变化 Fig.1 Zooplankton community variation in 2009 and 2010 including density, biomass, species, richness, diversity and evenness |
生物量在3—8月份的变化趋势与生物密度较为相似,在5月份达到全年最高值(2009年1751.88 mg/m3,2010年854.95 mg/m3)。2009年与2010年的生物量在继8月份下降之后又迅速上升,分别在10月份和12月份达到一年中的第2高峰。呈现出比较明显的双峰变化模型。2009年的生物量较2010年略高,年均生物量分别为648.48 mg/m3和489.32 mg/m3。
浮游动物种类数的动态变化呈现出明显的单峰模型,在3、4月份种类比较少,平均每个月份只监测到十几种浮游动物,5月份开始增加,到6—9月份达到全年的最高值(2009年48种,2010年40种),从9月份开始浮游动物种类数开始缓慢下降,一直到11、12月下降到最低值。
浮游动物丰富度指数表现出与种类数相似的动态变化趋势,在3—5月份丰富度较低,在6—10月份丰富度较高,分别在2009年的7月份以及2010年的6月份达到了年度的最高值2.31和1.97。11、12月浮游动物丰富度指数则又迅速下降到与3、4月份持平。2009年与2010年浮游动物年均丰富度指数分别为1.31和1.32。
2009年与2010年的浮游动物多样性变化趋势均表现出比较明显的双峰模型,即在5、6月份和9、10月份多样性指数比较高,而在其他月份多样性指数则较低。2009年多样性指数最高值出现在10月份为2.6,2010年多样性指数最高值出现在5月份为2.7,2009年多样性指数较2010年略高,其年均值分别为1.83和1.61。
浮游动物均匀度指数变化趋势与多样性指数较为相似,均匀度指数在5、6月份和9、10月份比较高,而在7、8月份比较低,表现出比较明显的双峰模型。2009年均匀度指数较2010年略高,其年均值分别为0.57和0.48。
2.4 优势种动态变化在75种浮游动物中共监测到4种优势种(表2),分别为强壮箭虫(Y=0.24)、中华哲水蚤(Y=0.13)、墨氏胸刺水蚤(Y=0.07)以及洪氏纺锤水蚤(Y=0.02),各优势种的动态变化如图2所示。

|
| 图 2 2009—2010年主要优势种的数量动态变化 Fig.2 Abundance variation of dominate species in 2009—2010 |
强壮箭虫在2009年变化趋势为单峰模型,5月份达到全年最低值20.4个/m3,到7月份迅速上升到最高峰155.2个/m3,年均值为75.8个/m3。2010年表现出双峰的变化趋势,在5月份达到一年中的次高峰156.4个/m3,到8月份达到最高峰215.4个/m3,年均值为74.3个/m3。
中华哲水蚤在2009年与2010年表现出极其相似的变化趋势,在3、4月份丰度比较低,而后迅速增加,分别在5月、6月达到全年的最高值250.4、294.7个/m3。中华哲水蚤丰度在6月份之后迅速下降,在8—12月份一直保持较低的丰度分布。
墨氏胸刺水蚤变化趋势与中华哲水蚤非常相似,2009年与2010年均在5月份达到最高值294.4、340.5个/m3。进入6月份后迅速下降,在7—12月份几乎监测不到其存在。墨氏胸刺水蚤在2009年和2010年年均丰度分别为62.1、123.1个/m3。
2009年与2010年洪氏纺锤水蚤丰度均在5月份达到全年最高值,但是两年丰度相差较大,2009年洪氏纺锤水蚤的年均丰度为37.6个/m3,而2010年仅为2.9个/m3。洪氏纺锤水蚤在5月份丰度最高,而在10—12月份基本监测不到其存在,表现出明显的季节变化特征。
2.5 浮游动物群落聚类分析对不同月份采集到的浮游动物群落组成进行聚类分析,结果如图3所示,以40%相似度为界限,各月份的浮游动物群落组成可以明显的划分为四大类别,第Ⅰ类别以3、4、5月份样品为代表,为春季样品,第Ⅱ类别以6、7、8月份样品为代表,为夏季样品,第Ⅲ类别以9、10月份样品为代表,为秋季样品,第Ⅳ类别以11、12月份样品为代表,为冬季样品。四大类别内部均表现出比较高的聚类效果。从图3可以看出,浮游动物的群落组成体现出了非常明显的季节特性,且不同年份间同一季节,甚至同一月份的浮游动物群落结构具有高度的相似性。如2009年8月与10年8月以及2009年7月与2010年7月样品,其群落结构组成相似度均在60%以上。说明浮游动物群落组成随季节变化的分布特征具有良好的稳定性与重现性。

|
| 图 3 2009—2010年莱州湾金城海域20个月份浮游动物聚类树状图 Fig.3 Cluster dendrogram of zooplankton in 20 sampling months in Jinchen area of Laizhou Bay 2009—2010 |
为了探讨浮游动物与环境因子的关系,对浮游动物群落的4项指标(丰度、种类数、生物量以及多样性指数)与水温、盐度等8项环境指标做相关性分析,结果如表3所示。水温与浮游动物种类数有极强的相关性,相关系数达到0.87(P < 0.01)。盐度与浮游动物的丰度有一定的正相关,相关系数为0.484。生物量与COD正相关,而与pH值负相关,其相关系数分别为0.466、-0.480。多样性指数与Chla有较好的正相关性,而与其它各环境因子的相关性较低。从表可知,水温与浮游动物种类数相关性最强,而透明度、TN、TP与浮游动物各个指标均无明显相关性。
| 理化指标 Environment factors | 丰度 Abundance | 种类数 Number of species | 生物量 Biomass | 多样性指数 Diversity index | ||||
| R | P | R | P | R | P | R | P | |
| *0.05显著水平(双尾); **0.01显著水平(双尾) | ||||||||
| pH | -0.095 | 0.692 | 0.185 | 0.434 | -0.480* | 0.032 | 0.070 | 0.771 |
| 水温 WT | 0.246 | 0.295 | 0.870** | 0.000 | 0.008 | 0.975 | 0.336 | 0.147 |
| 化学需氧量 COD | 0.172 | 0.469 | 0.394 | 0.086 | 0.466* | 0.038 | 0.229 | 0.332 |
| 盐度 Salinity | 0.484* | 0.031 | 0.437 | 0.054 | -0.226 | 0.338 | 0.223 | 0.344 |
| 叶绿素a Chl a | 0.221 | 0.349 | 0.182 | 0.444 | -0.105 | 0.659 | 0.494* | 0.027 |
| 透明度 Transparency | 0.249 | 0.289 | 0.062 | 0.795 | 0.374 | 0.104 | 0.063 | 0.791 |
| 总氮 TN | -0.129 | 0.588 | -0.196 | 0.407 | -0.014 | 0.954 | 0.088 | 0.711 |
| 总磷 TP | -0.078 | 0.745 | 0.194 | 0.413 | -0.010 | 0.966 | 0.247 | 0.294 |
为了进一步探讨环境因子与浮游动物群落结构的关系,对其进行CCA分析。在 CCA 排序图中,箭头表示环境因子,箭头连线的长短表示物种分布与环境因子相关性的大小。蒙特卡罗检验(Monte Carlo test) 表明,第 1 轴和全部轴均呈极显著差异(P < 0.01),故 CCA 分析的排序结果是可信的。通过图4可以看出影响浮游动物群落结构最主要的因素是水温,其次是盐度,影响最小的是透明度和Chla。这与表3所示的结果相吻合。不同浮游动物对环境的要求不一样,某一个环境条件对某些种类比较适宜,而对其它某些种类则不一定适宜。从图4中可知,桡足类幼体、海胆长腕幼虫、短尾类幼体、帚毛类幼体等大多数浮游幼虫都分布在水温和盐度较高的6、7月份,与水温、盐度呈一定的正相关性;背针胸刺水蚤、强额拟哲水蚤、太平洋纺锤水蚤、强额拟哲水蚤等大部分的桡足类以及水螅水母类都分布在水温较高且总磷含量较高的8月、9月,与水温、总磷呈现较好的正相关性;而钩虾、麦秆虫等端足类则多分布在11、12月份,与水温、盐度呈现较强的负相关,与总氮呈现较强的正相关。

|
| 图 4 浮游动物群落分布与环境因子典范对应分析 Fig.4 CCA of the zooplankton and associated environmental factors |
本次调查共鉴定浮游动物75种,分属14大类,其中桡足类无论是丰度还是种类均占有绝对的优势。毕洪生[13]等分析1959年全国海洋普查渤海浮游动物样品时发现桡足类是渤海浮游动物的主要构成者,张武昌[14]等在调查1998—1999年渤海浮游动物群落构成时也报道过桡足类的优势地位,这与本次浮游动物调查结果相一致。本次调查发现浮游动物的种类与丰度分布均呈现比较明显的季节变化特征。在3—5月份由于表层水温升高,光度增强,营养盐类丰富,浮游植物主要是硅藻迅速繁殖,数量激增,而浮游动物由于水温适宜和食料丰富也出现大量繁殖[15],浮游动物的丰度在5月份达到最高值,种类数也在6、7月份达到峰值。在本次调查中,浮游动物的种类变化与多样性指数变化没有很好的吻合(图1),如2009年8月份浮游动物的种类数较高(35种),而多样性指数反而仅为1.16,仔细观察发现2009年8月份样品中强壮箭虫的丰度比较高,而其他浮游动物的丰度较低,即8月份样品的物种均匀度较低(图1),最终导致浮游动物多样性指数较低。因此在判断浮游动物的群落组成时不能单单通过一两个指标,而要综合考虑多种因素才能更加真实地反映群落组成情况。
莱州湾金城海域浮游动物主要优势种包括强壮箭虫、中华哲水蚤、墨氏胸刺水蚤以及洪氏纺锤水蚤。4种优势种在5—6月份数量均比较多,基本达到年度最高峰,这与此时期水温比较适宜,食料丰富不无关系。中华哲水蚤、墨氏胸刺水蚤和洪氏纺锤水蚤这3种优势种,在5—6月份达到高峰以后迅速下降到极低水平,在8—12月份近于绝迹,具有明显的季节分布特性。张武昌[14]分析过1998—1999年春秋两季渤海浮游动物样品时发现,小拟哲水蚤和拟长腹剑水蚤是秋季优势种,而洪氏纺锤水蚤和墨氏胸刺水蚤是春季优势种,优势种随季节交替产生明显的变化。本研究较1998—1999年数据相比,优势种产生了较大的变化,且优势种的丰度大幅降低,产生这种差异的原因可能与整体的海洋环境变化有关,孙松报道[16],近年来胶州湾浮游动物丰度较本世纪初有明显下降;近50年来,受暖流增强的影响,南黄海浮游动物物种数量增加,优势种组成种类多样化,主要优势种中华哲水蚤和强壮箭虫比例增加,小型桡足类和暖水性种类数量增加[17],与本文研究相符。
由于气候的季节变化,海洋中水文条件也有季节变化的现象,因此,在季节更替中,经常有新的种类出现和旧的种类消失,浮游动物的群落结构也相应地发生变化[15]。2009年与2010年浮游动物群落组成的变化趋势较为相似,从聚类分析结果看,浮游动物的群落结构分为明显地春、夏、秋、冬四大季节类型。Fabrizio[18]等在对亚得里亚海进行长达30a的调查研究后发现,浮游动物的季节分布特性具有较好的稳定性。本次调查结果也证实了这一点。不同年份间同一季节,甚至同一月份的浮游动物群落结构具有高度的相似性。如2009年7月份与2010年7份样品以及2009年8月与2010年8月,其群落结构组成相似度均在60%以上。
3.2 环境因子对浮游动物的影响浮游动物的分布受着各种环境因子综合作用的影响,其中温度、盐度是影响浮游动物分布的重要因子[19, 20, 21]。由于各种浮游动物对温度与盐度变化的忍受力不同,因此在数量和种类组成上都随着温度、盐度的变化,而在分布上表现出差异[15]。温度直接影响生物有机体的体温,体温的高低又决定了浮游动物新陈代谢过程的强度和特点、有机体的生长和发育速度、繁殖、行为等,从而影响浮游动物的生物量和分布[22, 23, 24]。金琼贝等[25]在室内实验中探讨了 20—50 ℃范围内浮游动物群落结构与温度的关系。实验结果表明,种类数在 30 ℃左右最多,38 ℃开始明显减少;枝角类、桡足类、无节幼体数量在 30 ℃以下随温度的升高而增多,而后逐渐下降,40 ℃时数量锐减。莱州湾金城海域全年水温波动范围在2.2—27.4 ℃之间,在此范围内浮游动物的种类与水温呈极显著的正相关性(P < 0.01),较好的印证了已有研究[25]。
典型对应分析结果显示,盐度是继水温影响海洋浮游动物群落结构的第二环境因素,浮游动物丰度随盐度升高而显著增加(P < 0.05)。徐兆礼[26, 27]和Gao等[28]对长江口及邻近水域浮游动物群落研究表明,盐度是影响长江口浮游动物群落特征最重要的环境因子。李开枝等[29, 30]对珠江口及邻近海域以及 Mouny等[31]对英吉利海峡塞纳河口(Seine estuary)的研究进一步表明,在河口水域盐度是影响浮游动物分布最重要的环境因子。浮游动物适应盐度的变化与渗透压和离子调节机制密切相关。在正常环境中,浮游动物体内的体液与环境保持渗透压和离子浓度的平衡;当盐度改变时,浮游动物可以通过呼吸和排泄器官的调节机制来适应变化了的渗透压和离子浓度,以维持正常的生理活动[32]。
除温度和盐度外,总磷含量对桡足类的影响也比较大,如图4所示,大部分桡足类都分布在总磷含量较高的区域。总磷对桡足类的影响可能是通过对浮游植物数量的影响来间接实现。值得注意的是,CCA结果显示本文所选环境因子对浮游动物群落分布的累计贡献率仅为49.8%,说明,除所选环境因子外,浮游动物群落结构还受浮游植物,鱼类捕食,以及水产养殖等人类活动等其他多方面的影响,要想进一步深入研究影响浮游动物群落结构的因素,不仅要从时间与空间上进行大尺度调查,更需从影响因素上进行进一步的筛选与整合。
| [1] | 鲍献文, 万修全, 高郭平, 吴德星. 渤海、黄海、东海 AVHRR海表温度场的季节变化特征. 海洋学报, 2002, 24(5): 125-133. |
| [2] | Lin C L, Su J L, Xu B R, Tang Q S. Long-term variations of temperature and salinity of the Bohai Sea and their influence on its ecosystem. Progress in Oceanography, 2001, 49(1-4): 7-19. |
| [3] | 张莹, 刘元进, 张英, 徐炳庆, 吕振波. 莱州湾多毛类底栖动物生态特征及其对环境变化的响应. 生态学杂志, 2012, 31(4): 888-895. |
| [4] | 金显仕, 邓景耀. 莱州湾渔业资源群落结构和生物多样性的变化. 生物多样性, 2000, 8(1): 65-72. |
| [5] | Zhou S C, Jin B S, Guo L, Qin H M, Chu T J, Wu J H. Spatial distribution of zooplankton in the intertidal marsh creeks of the Yangtze River Estuary, China. Estuarine, Coastal and Shelf Science, 2009, 85(3): 399-406. |
| [6] | Mazumder D, Saintilan N, Williams R J. Trophic relationships between itinerant fish and crab larvae in a temperate Australian saltmarsh. Marine and Freshwater Research, 2006, 57(2): 193-199. |
| [7] | David V, Sautour B, Chardy P, Leconte M. Long-term changes of the zooplankton variability in a turbid environment: the Gironde estuary (France). Estuarine, Coastal and Shelf Science, 2005, 64(2-3): 171-184. |
| [8] | Giraudel J L, Lek S. A comparison of self-organizing map algorithm and some conventional statistical methods for ecological community ordination. Ecological Modelling, 2001, 146(1-3): 329-339. |
| [9] | Huang C J, Lin X P, Lin J D, Du H, Dong Q X. Population dynamics of Pseudo-nitzschia pungens in Zhelin Bay, China. Journal of the Marine Biological Association of the United Kingdom, 2009, 89(4): 663-668. |
| [10] | 刘亚林, 姚炜民, 张树刚, 陈雷. 2008年夏季飞云江口浮游植物分布特征及与环境因子的相关性. 海洋环境科学, 2009, 28(S1): 31-41. |
| [11] | 国家质检总局, 国家标准化委员会. 海洋调查规范第6部分: 海洋生物调查. 北京: 中国标准出版社, 2007. |
| [12] | 国家质检总局, 国家标准化委员会. 海洋监测规范第4部分: 海水分析. 北京: 中国标准出版社, 2007. |
| [13] | 毕洪生, 孙松, 高尚武, 张光涛. 渤海浮游动物群落生态特点 Ⅱ. 桡足类数量分布及变动. 生态学报, 2001, 21(2): 177-185. |
| [14] | 张武昌, 王克, 高尚武, 王荣. 渤海春季和秋季的浮游动物. 海洋与湖沼, 2002, 33(6): 630-639. |
| [15] | 郑重, 李少菁, 许振祖. 海洋浮游生物学. 北京: 海洋出版社, 1984: 139-491. |
| [16] | 孙松, 周克, 杨波, 张永山, 吉鹏. 胶州湾浮游动物生态学研究 Ⅰ. 种类组成. 海洋与湖沼, 2008, 39(1): 1-7. |
| [17] | 左涛, 王荣, 陈亚瞿, 高尚武, 王克. 春季和秋季东、黄海陆架区大型网采浮游动物群落划分. 生态学报, 2005, 25(7): 1531-1540. |
| [18] | Fabrizio B A, Gianpiero C, Francesco A, Mauro B, Franco B, Elisa C, Amelia D L, Alessandra P, Cosimo S, Giorgio S. Plankton communities in the northern Adriatic Sea: Patterns and changes over the last 30 years. Estuarine, Coastal and Shelf Science, 2012, 115:125-137. |
| [19] | Jin B S, Fu C Z, Zhong J S, Li B, Chen J K, Wu J H. Fish utilization of a salt marsh intertidal creek in the Yangtze River estuary, China. Estuarine, Coastal and Shelf Science, 2007, 73(3-4): 844-852. |
| [20] | Froneman P W. Zooplankton community structure and bio-mass in a southern African temporarily open/closed estuary. Estuarine, Coastal and Shelf Science, 2004, 60(1): 125-132. |
| [21] | Marques S C, Azeiteiro U M, Marques J C, Neto J M, Pardal M A. Zooplankton and ichthyoplankton communities in a temperate estuary: spatial and temporal patterns. Journal of Plankton Research, 2006, 28(3): 297-312. |
| [22] | Elliott J A. A comparison of thermal polygons for British freshwater teleosts. Freshwater Forum, 1995, 5(3): 178-184. |
| [23] | Devreker D, Souissi S, Seuront L. Effects of chloropHyll concentration and temperature variation on the reproduction and survival of Temora longicornis (Copepoda, Calanoida) in the Eastern English Channel. Journal of Experimental Marine Biology and Ecology, 2005, 318(2): 145-162. |
| [24] | Lenz P H, Hower A E, Hartline D K. Temperature compensation in the escape response of a marine copepod, Calanus finmarchicus (Crustacea). The Biological Bulletin, 2005, 209(1): 75- 85. |
| [25] | 金琼贝, 盛连喜, 张然. 温度对浮游动物群落的影响. 东北师范大学学报: 自然科学版, 1991, (4): 103-111. |
| [26] | 徐兆礼. 长江口邻近水域浮游动物群落特征及变动趋势. 生态学杂志, 2005, 24(7): 780-784. |
| [27] | 徐兆礼. 中国海洋浮游动物研究的新进展. 厦门大学学报: 自然科学版, 2006, 45(Z2): 16-23. |
| [28] | Gao Q, Xu Z L, Zhuang P. The relation between distribution of zooplankton and salinity in the Changjiang Estuary. Chinese Journal of Oceanology and Limnology, 2008, 26(2): 178-185. |
| [29] | Li K Z, Yin J Q, Huang L M, Tan Y H. Spatial and temporal variations of mesozooplankton in the Pearl River estuary, China. Estuarine, Coastal and Shelf Science, 2006, 67(4): 543-552. |
| [30] | 李开枝, 尹健强, 黄良民, 谭烨辉, 许战洲. 珠江口浮游动物的群落动态及数量变化. 热带海洋学报, 2005, 24(5): 60-68. |
| [31] | Mouny P, Dauvin J C. Environmental control of mesozooplankton community structure in the Seine estuary (English Channel). Oceanologica Acta, 2002, 25(1): 13-22. |
| [32] | 黄加祺, 郑重. 九龙江口桡足类和盐度关系的初步研究. 厦门大学学报: 自然科学版, 1984, 23(4): 497-505. |