 2015, Vol. 35
2015, Vol. 35文章信息
- 郑超超, 伊力塔, 张超, 余树全, 库伟鹏, 钱逸凡, 凌骅
- ZHENG Chaochao, YI Lita, ZHANG Chao, YU Shuquan, KU Weipeng, QIAN Yifan, LING Hua
- 浙江江山公益林物种种间关系及CCA排序
- Interspecific relationship and canonical correspondence analysis of the dominant species in ecological service forest of Jiangshan City in Zhejiang Province
- 生态学报, 2015, 35(22): 7511-7521
- Acta Ecologica Sinica, 2015, 35(22): 7511-7521
- http://dx.doi.org/10.5846/stxb201403240538
-
文章历史
- 收稿日期: 2014-03-24
- 网络出版日期: 2015-04-20






2. 国家林业局华东林业调查规划设计院, 杭州 310019
2. The East China Institute of Forest Inventory and Planning of the State Forestry Administration, Hangzhou 310019, China
The uniqueness of this study is introducing the species-environment relations (CCA) into the analyzing framework to solve the complex inter-species, intra-species, and plant-environment relationships to enrich niche theory. This study also demonstrated that importance values were directly correlated with the breadth of ecological niches. In general, the higher the niche overlap between two species, the more similar the environmental conditions in which they were found. In addition, the analysis showed that the distribution frequency was an important factor affecting the niche breadth and species niche overlap. We observed positive relationships between the distribution frequency and the niche overlap probabilities, namely, the higher the distribution frequency the higher the probability of species niche overlap. Furthermore, the interspecific relationships reflected the interconnectedness of different species in the spatial distribution and represented mutual attraction or repulsion between species and their environment. In our study area, most of the interspecific relationships between the dominant species showed a positive correlation. There were many factors, including local climate, soil types, topography, and landforms, that influenced the distribution of plant species in a given community.
In summary, this study found the following general patterns for interspecific relationships: if species have a narrow niche, they also have a narrower spatial distribution, and specialize on certain resources with less overlap with other species in the CCA sequencing diagram, characterized as a tendency of deviating from the high species distribution areas. However, if species have a wide niche, they also have a wider distribution range and greater overlaps with other species, and occupy a majority of the species distribution area on the CCA sequencing diagram. For a given species, if it does not have significant interactions with other species, the higher its niche overlap values are with another species, the closer the distances between the species on the CCA sequencing diagram. If there is a positive correlation, this correlation is significant and there are shorter distances among species on the CCA sequencing diagram; while if there is a negative correlation, this negative correlation is also significant and the distances are longer among species on the CCA sequencing diagram. Our study not only indicates the complex interspecific and species-environment relationships for constructing ecological public welfare forests in subtropical regions, especially in China, but also provides recommendations of how to use our research in practice, including providing feasible suggestions to protect species diversity and accelerate positive community succession.
植物种间关系是指不同物种在空间分布上的相互关联性,通常由于群落生境的差异影响了植物种分布而引起,是对各种物种在不同生境中相互影响、相互作用所形成的有机联系反映[1]。一直以来,植物种间关系、植物-环境的关系与环境对植物分布和构建的影响都是植物生态学研究的基础内容之一[2, 3]。
一个多世纪以来,植物生态学家们从未间断过对决定物种分布及多样性因子的探索[4]。生态学家尝试使用各种方法来了解植物种间关系,包括生态位理论、种间联结及植物分类排序等方法。其中,生态位理论和种间联结常用于研究种间关系,以往的研究通常利用这两种方法,基于物种对资源的利用能力来解释分析物种间的关系。研究表明,物种间的联结性与其生态位重叠之间有较大的相关性,种间正联结性越强,则生态位重叠值就越大,反之,种间的负联结性越强,其生态位重叠值就越小[5];生态位宽度较大的物种对资源利用能力较强,与其它种群间的生态位重叠一般较大,物种间的竞争较强[6]。
近年来,随着植被分类排序被广泛应用,这种方法已然成为植被生态学研究物种-环境关系的重要方法之一[7, 8, 9],运用该方法不仅可以把植物群落的分布格局与环境资料进行客观定量地分析,而且可以给出群落类型分布及其环境梯度的具体关系[10, 11],进而能充实生态位理论与种间联结方法对物种-环境关系分析的薄弱之处,具有重要的生态学研究意义。
本研究以浙江省江山市公益林监测数据为依托,选取群落优势种为研究对象,基于生态位理论、种间联接及CCA(Canonical Correspondence Analysis)排序3种分析方法研究种间关系及物种受环境因子的影响。利用这3种方法在分析过程中的相互补充解释,完善以往运用生态位宽度、种间联结方法分析关于植物种间关系的研究中缺少详细的物种-环境关系解释的难题,同时也为植物排序中无法得到相应环境因子解释的部分做详尽的种间关系分析,不但为江山市公益林建设与管理提供依据,而且为解释物种种间关系及植物-环境关系提供新的思考方向。
1 研究区概况浙江省江山市地处浙江西南部,地处东经118°22′37″至118°48′48″和北纬28°15′26″至28°53′27″之间,是浙闽赣三省交界。土地总面积为20.19×104 hm2,其中有林地面积14.72×104 hm2,森林覆盖率67.7%。江山市地理纬度较低,温暖湿润,四季分明,光照充足,降水丰沛,具明显的山地立体气候特征,属浙闽山丘甜槠木荷林植被区。
江山市共有公益林面积5.26×104 hm2,其中,国家级重点公益林面积达2.06×104 hm2,占全市公益林面积的39.16%,省级重点公益林3.20×104 hm2,占全市公益林面积的60.84%。根据森林类型大致可分为6类,即松林、杉木林、阔叶林、针阔混交林、毛竹林、灌木林,按群落类型划分如表1。
| 群落类型Forest types | 松林Pine forests | 杉木林Chinese fir forests | 阔叶林Broad-leaved forests | 针阔混交林Mixed coniferous and broad-leaved forests | 毛竹林Phyllostachys heterocycla forests | 灌木林Shrub forests | 合计Total |
| 面积Area/hm 2 | 12700 | 13620 | 17380 | 4630 | 3440 | 790 | 54560 |
| 面积所占百分比/%Area Percentage | 24.16 | 25.91 | 33.07 | 8.81 | 6.54 | 1.51 | 100 |
以公益林小班作为总体样本,按系统抽样抽取监测小班,选取典型地段设置面积为20 m×20 m的固定样地,详细记录样地信息,包括经纬度、海拔、坡度、坡向、坡位、土壤等环境因子[12];样地内乔木层(胸径大于5.0 cm)采用每木调查(包括树种名称、树高、胸径、枝下高等),同时在每块样地对角线上均匀设置3个2 m×2 m的灌草固定小样方(对角线两侧的小样方距样地边缘至少1 m),详细记录灌木种类、株数、盖度、高度、地径以及草本种类、数量、盖度、高度等相关指标[13]。
在江山市80个公益林样地具体群落类型如表2所示,6种群落类型所占比例与江山市公益林各群落所占比例大致相同,具有代表性。
| 群落类型Community types | 松林Pine forests | 杉木林Chinese fir forests | 阔叶林Broad-leaved forest | 针阔混交林Mixed coniferous and broad-leaved forests | 毛竹林Phyllostachys heterocycla forests | 灌木林Shrub forests | 合计Total |
| 样地数量/个The samples quantity | 21 | 24 | 20 | 6 | 7 | 2 | 80 |
| 百分比Percentage/% | 26.25 | 30.00 | 25.00 | 7.50 | 8.75 | 2.5 | 100 |
重要值表示物种的优势程度,本研究共选取213个物种进行了重要值计算。其公式为:
(1) 乔木层重要值 IV(%)=(相对高度+相对显著度+相对多度)/3
(2) 灌木层、草本层重要值 IV(%)=(相对盖度+相对多度)/2
2.2.2 生态位宽度生态位宽度(niche breadth)一般表示一个种群在一个群落中所利用的不同资源的综合,反映物种对资源利用的程度[14],本研究采用Levins公式中Shannon指数,公式为:
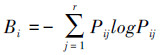
生态位重叠(niche overlap)是指两个或多个物种对统一资源因素(食物、营养成分、空间等)的共同利用程度,本研究中,生态位重叠采用Levins生态位重叠公式计算[15],公式如下:
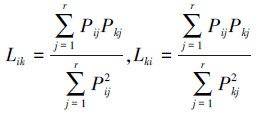
采用联结系数AC方法:
当ad≥bc时,AC=(ad-bc)/[(a+b)(b+d)];
当bc>ad时,且d≥a时,AC=(ad-bc)/[(a+b)(a+c)];
当bc>ad时,且d<a时,AC=(ad-bc)/[(b+d)(d+c)];
AC的值域为[-1,1];AC的值越接近1,表明物种间的正联结性越强;AC的值越接近-1,表明物种间的负联结性越强;AC的值为0,表明物种间完全独立[16]。
由于取样的非连续性,原始数据为事件存在与否的二元数据,因此非连续性数据的x 2值用Yates的连续校正公式计算:
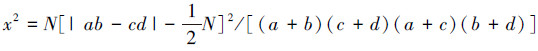
典范对应分析(CCA排序)是一种非线性多元直接梯度分析方法,它把分析与多元回归结合起来[18, 19]。本研究在重要值的基础上,选取乔、灌、草3个层中重要值大于1的物种进行排序分析,即乔木层13个物种、灌木层18个物种(6个物种为乔木幼苗)、草本层11个物种,共计研究对象42个。利用CANOCO 5.0软件,对42×80维重要值矩阵进行趋势对应分析(DCA排序),并结合6个环境数据(海拔、坡度、坡向、坡位、土壤厚度、腐殖质厚度)的6×80维环境矩阵进行CCA排序。
3 结果与分析 3.1 生态位 3.1.1 生态位宽度生态位宽度是度量植物种群对资源环境利用现状的尺度,种群生态位宽度越大,它对环境的适应能力就越强,相应的对资源利用也就越充分[20]。表3为研究区内优势物种的重要值及生态位宽度表,由表3分析可知,在乔木层中,杉木的生态位宽度最大(IV=30.30,Bi=1.504),湿地松(IV=3.62,Bi=0.705)与毛竹(IV=12.94,Bi=0.858)的生态位宽度最小;灌木层中,隔药柃(IV=8.07,Bi=1.678)的生态位宽度最大,而苦竹(IV=2.38,Bi=0.786)的生态位宽度最小;草本层中,葫芦藓(IV=12.50,Bi=1.757)的生态位宽度最大,三花悬钩子(IV=1.08,Bi=0.643)、野菊(IV=1.01,Bi=0.670)的生态位宽度最小。
| 层 Layer | 种号Number | 物种种名Species | 重要值Importance value (IV) | 生态位宽度Niche breath (Bi) |
| 乔木层 Arbor layer | 1 | 杉木 Cunninghamia lanceolata | 30.30 | 1.504 |
| 2 | 马尾松 Pinus massoniana | 14.53 | 1.444 | |
| 3 | 毛竹 Phyllostachys heterocycla | 12.94 | 0.858 | |
| 4 | 木荷 Schima superba | 6.54 | 1.327 | |
| 5 | 石栎 Lithocarpus glabra | 4.95 | 1.296 | |
| 6 | 湿地松 Pinus elliottii | 3.62 | 0.705 | |
| 7 | 甜槠 Castanopsis eyrei | 2.55 | 1.104 | |
| 8 | 青冈 Cyclobalanopsis glauca | 2.40 | 1.125 | |
| 9 | 苦槠 Castanopsis sclerophylla | 2.32 | 0.990 | |
| 10 | 短柄枹 Quercus glandulifera | 1.95 | 1.170 | |
| 11 | 白栎 Quercus fabri | 1.55 | 1.088 | |
| 12 | 山合欢 Albizia.kalkora | 1.30 | 1.162 | |
| 13 | 黄檀 Dalbergia hupeana | 1.05 | 0.891 | |
| 灌木层 Shrub layer | 14 | 檵木 Loropetalum chinensis | 12.13 | 1.571 |
| 15 | 杜鹃 Rhododendron simsii | 9.56 | 1.601 | |
| 16 | 隔药柃 Eurya muricata | 8.07 | 1.678 | |
| 17 | 杉木 Cunninghamia lanceolata | 7.32 | 1.465 | |
| 18 | 阔叶箬竹 Indocalamus latifolius | 5.78 | 1.047 | |
| 19 | 石栎 Lithocarpus glabra | 4.49 | 1.301 | |
| 20 | 白栎 Quercus fabri | 3.12 | 1.325 | |
| 21 | 油茶 Camellia oleifera | 2.76 | 1.266 | |
| 22 | 乌饭 Vaccinium bracteatum | 2.75 | 1.509 | |
| 23 | 菝葜 Smilax china | 2.48 | 1.447 | |
| 24 | 大血藤 Sargentodoxa cuneata | 2.42 | 1.366 | |
| 25 | 苦竹 Pleioblastus amarus | 2.38 | 0.786 | |
| 26 | 金樱子 Rosa laevigata | 1.62 | 1.181 | |
| 27 | 乌药 Lindera aggregata | 1.55 | 1.051 | |
| 28 | 山苍子 Litsea cubeba | 1.46 | 1.214 | |
| 29 | 青冈 Cyclobalanopsis glauca | 1.41 | 0.990 | |
| 30 | 木荷 Schima superb | 1.33 | 0.967 | |
| 31 | 山胡椒 Lindera glauca | 1.10 | 0.957 | |
| 草本层 Herb layer | 32 | 紫萁 Osmunda japonica | 17.49 | 1.438 |
| 33 | 蕨 Pteridium aquilinum | 17.42 | 1.621 | |
| 34 | 莎草 Cyperus rotundus | 14.01 | 1.635 | |
| 35 | 葫芦藓 Funaria hygrometrica | 12.50 | 1.757 | |
| 36 | 芒萁 Dicranopteris pedata | 5.60 | 1.350 | |
| 37 | 粗叶悬钩子 Rubus alceifolius | 5.41 | 0.922 | |
| 38 | 淡竹叶 Lophatherum gracile | 1.96 | 1.285 | |
| 39 | 海金沙 Lygodium japonicum | 1.65 | 0.818 | |
| 40 | 蛇莓 Duchesnea indica | 1.14 | 0.740 | |
| 41 | 三花悬钩子 Rubus trianthus | 1.08 | 0.643 | |
| 42 | 野菊 Dendranthema indica | 1.01 | 0.670 |
生态位重叠是两个物种在与生态因子联系上具相似性,当两个物种利用同一种资源或共同占有某一资源时,就会出现生态位重叠现象[6]。对江山市公益林各层优势种进行生态位重叠计算,共计572对种对,结果如表4—表7,其中,表7为不同层生态位重叠情况统计表。
| 物种Species | 生态位重叠 Niche overlap | |||||||||||||
| 1 | 2 | 3 | 4 | 5 | 6 | 7 | 8 | 9 | 10 | 11 | 12 | 13 | ||
| 生态位重叠Niche overlap | 1 | 0.101 | 0.004 | 0.080 | 0.041 | 0.016 | 0.044 | 0.043 | 0.022 | 0.139 | 0.087 | 0.054 | 0.044 | |
| 2 | 0.124 | 0.001 | 0.034 | 0.195 | 0.006 | 0.020 | 0.079 | 0.124 | 0.060 | 0.150 | 0.085 | 0.051 | ||
| 3 | 0.016 | 0.002 | 0.005 | 0.002 | 0.000 | 0.003 | 0.000 | 0.014 | 0.001 | 0.000 | 0.000 | 0.000 | ||
| 4 | 0.152 | 0.052 | 0.002 | 0.259 | 0.030 | 0.435 | 0.109 | 0.028 | 0.197 | 0.055 | 0.093 | 0.050 | ||
| 5 | 0.076 | 0.296 | 0.001 | 0.253 | 0.007 | 0.254 | 0.497 | 0.132 | 0.355 | 0.035 | 0.037 | 0.014 | ||
| 6 | 0.100 | 0.031 | 0.000 | 0.098 | 0.023 | 0.000 | 0.000 | 0.000 | 0.007 | 0.000 | 0.000 | 0.000 | ||
| 7 | 0.116 | 0.043 | 0.002 | 0.595 | 0.357 | 0.000 | 0.144 | 0.025 | 0.413 | 0.031 | 0.058 | 0.042 | ||
| 8 | 0.128 | 0.194 | 0.000 | 0.170 | 0.797 | 0.000 | 0.165 | 0.131 | 0.111 | 0.146 | 0.028 | 0.067 | ||
| 9 | 0.099 | 0.446 | 0.015 | 0.066 | 0.312 | 0.000 | 0.042 | 0.194 | 0.061 | 0.099 | 0.048 | 0.131 | ||
| 10 | 0.342 | 0.122 | 0.000 | 0.255 | 0.471 | 0.003 | 0.390 | 0.092 | 0.034 | 0.017 | 0.081 | 0.045 | ||
| 11 | 0.258 | 0.363 | 0.000 | 0.085 | 0.056 | 0.000 | 0.035 | 0.144 | 0.066 | 0.020 | 0.529 | 0.299 | ||
| 12 | 0.219 | 0.279 | 0.000 | 0.196 | 0.079 | 0.000 | 0.090 | 0.037 | 0.044 | 0.132 | 0.718 | 0.171 | ||
| 13 | 0.245 | 0.232 | 0.000 | 0.145 | 0.042 | 0.000 | 0.090 | 0.124 | 0.165 | 0.101 | 0.562 | 0.236 | ||
| 物种Species | 生态位重叠 Niche overlap | ||||||||||||||||||
| 14 | 15 | 16 | 17 | 18 | 19 | 20 | 21 | 22 | 23 | 24 | 25 | 26 | 27 | 28 | 29 | 30 | 31 | ||
| 生态位重叠Niche overlap | 14 | 0.382 | 0.335 | 0.174 | 0.018 | 0.174 | 0.379 | 0.213 | 0.431 | 0.166 | 0.183 | 0.029 | 0.205 | 0.105 | 0.120 | 0.055 | 0.019 | 0.079 | |
| 15 | 0.378 | 0.448 | 0.163 | 0.061 | 0.248 | 0.333 | 0.075 | 0.359 | 0.156 | 0.126 | 0.024 | 0.063 | 0.070 | 0.109 | 0.065 | 0.141 | 0.047 | ||
| 16 | 0.262 | 0.354 | 0.339 | 0.113 | 0.196 | 0.134 | 0.129 | 0.290 | 0.120 | 0.306 | 0.055 | 0.060 | 0.118 | 0.276 | 0.057 | 0.066 | 0.063 | ||
| 17 | 0.218 | 0.206 | 0.542 | 0.065 | 0.139 | 0.085 | 0.159 | 0.251 | 0.105 | 0.376 | 0.015 | 0.063 | 0.130 | 0.103 | 0.018 | 0.178 | 0.034 | ||
| 18 | 0.064 | 0.220 | 0.514 | 0.183 | 0.107 | 0.026 | 0.068 | 0.201 | 0.046 | 0.157 | 0.387 | 0.176 | 0.016 | 0.071 | 0.072 | 0.075 | 0.073 | ||
| 19 | 0.350 | 0.503 | 0.503 | 0.223 | 0.061 | 0.062 | 0.160 | 0.361 | 0.091 | 0.233 | 0.017 | 0.078 | 0.013 | 0.061 | 0.122 | 0.079 | 0.032 | ||
| 20 | 0.672 | 0.596 | 0.303 | 0.121 | 0.013 | 0.055 | 0.208 | 0.211 | 0.214 | 0.072 | 0.081 | 0.194 | 0.086 | 0.120 | 0.044 | 0.013 | 0.041 | ||
| 21 | 0.494 | 0.176 | 0.382 | 0.294 | 0.044 | 0.185 | 0.272 | 0.311 | 0.166 | 0.196 | 0.005 | 0.084 | 0.109 | 0.209 | 0.356 | 0.088 | 0.014 | ||
| 22 | 0.519 | 0.438 | 0.447 | 0.242 | 0.068 | 0.216 | 0.144 | 0.162 | 0.194 | 0.161 | 0.019 | 0.101 | 0.099 | 0.164 | 0.155 | 0.030 | 0.044 | ||
| 23 | 0.390 | 0.370 | 0.361 | 0.196 | 0.030 | 0.107 | 0.284 | 0.168 | 0.379 | 0.105 | 0.027 | 0.093 | 0.075 | 0.141 | 0.022 | 0.046 | 0.019 | ||
| 24 | 0.309 | 0.215 | 0.660 | 0.506 | 0.075 | 0.195 | 0.069 | 0.143 | 0.226 | 0.075 | 0.031 | 0.073 | 0.145 | 0.216 | 0.050 | 0.060 | 0.049 | ||
| 25 | 0.205 | 0.171 | 0.490 | 0.083 | 0.765 | 0.058 | 0.318 | 0.016 | 0.111 | 0.082 | 0.127 | 0.401 | 0.070 | 0.207 | 0.010 | 0.006 | 0.019 | ||
| 26 | 0.566 | 0.175 | 0.214 | 0.138 | 0.137 | 0.108 | 0.302 | 0.101 | 0.231 | 0.109 | 0.120 | 0.159 | 0.086 | 0.051 | 0.040 | 0.028 | 0.036 | ||
| 27 | 0.401 | 0.273 | 0.579 | 0.399 | 0.017 | 0.025 | 0.187 | 0.180 | 0.317 | 0.123 | 0.329 | 0.038 | 0.119 | 0.121 | 0.199 | 0.124 | 0.019 | ||
| 28 | 0.314 | 0.286 | 0.921 | 0.213 | 0.052 | 0.079 | 0.176 | 0.235 | 0.356 | 0.156 | 0.334 | 0.077 | 0.048 | 0.083 | 0.029 | 0.025 | 0.105 | ||
| 29 | 0.258 | 0.309 | 0.341 | 0.067 | 0.095 | 0.284 | 0.117 | 0.719 | 0.601 | 0.044 | 0.138 | 0.006 | 0.067 | 0.242 | 0.052 | 0.067 | 0.000 | ||
| 30 | 0.079 | 0.579 | 0.344 | 0.577 | 0.086 | 0.160 | 0.029 | 0.155 | 0.103 | 0.080 | 0.145 | 0.004 | 0.041 | 0.131 | 0.039 | 0.059 | 0.004 | ||
| 31 | 0.398 | 0.236 | 0.405 | 0.136 | 0.104 | 0.080 | 0.115 | 0.031 | 0.184 | 0.040 | 0.147 | 0.014 | 0.066 | 0.025 | 0.202 | 0.000 | 0.005 | ||
| 物种Species | 生态位重叠 Niche overlap | |||||||||||
| 32 | 33 | 34 | 35 | 36 | 37 | 38 | 39 | 40 | 41 | 42 | ||
| 生态位重叠Niche overlap | 32 | 0.321 | 0.200 | 0.396 | 0.241 | 0.015 | 0.241 | 0.017 | 0.011 | 0.030 | 0.001 | |
| 33 | 0.202 | 0.344 | 0.662 | 0.240 | 0.035 | 0.147 | 0.116 | 0.019 | 0.006 | 0.004 | ||
| 34 | 0.132 | 0.362 | 0.664 | 0.147 | 0.095 | 0.240 | 0.084 | 0.049 | 0.038 | 0.038 | ||
| 35 | 0.183 | 0.488 | 0.465 | 0.182 | 0.058 | 0.194 | 0.050 | 0.024 | 0.057 | 0.009 | ||
| 36 | 0.305 | 0.483 | 0.281 | 0.498 | 0.029 | 0.269 | 0.024 | 0.127 | 0.155 | 0.029 | ||
| 37 | 0.052 | 0.196 | 0.501 | 0.441 | 0.079 | 0.268 | 0.085 | 0.221 | 0.067 | 0.000 | ||
| 38 | 0.311 | 0.302 | 0.468 | 0.540 | 0.274 | 0.099 | 0.067 | 0.118 | 0.115 | 0.025 | ||
| 39 | 0.069 | 0.733 | 0.505 | 0.434 | 0.076 | 0.097 | 0.208 | 0.000 | 0.157 | 0.000 | ||
| 40 | 0.054 | 0.148 | 0.352 | 0.248 | 0.481 | 0.303 | 0.439 | 0.000 | 0.000 | 0.037 | ||
| 41 | 0.163 | 0.050 | 0.312 | 0.672 | 0.669 | 0.104 | 0.486 | 0.215 | 0.000 | 0.000 | ||
| 42 | 0.008 | 0.043 | 0.344 | 0.111 | 0.138 | 0.000 | 0.118 | 0.000 | 0.046 | 0.000 | ||
| 层Layer | 0≤ L<0.01 | 0.01≤ L<0.1 | 0.1≤ L<0.3 | 0.3≤ L<0.5 | 0.5≤ L<0.8 | 0.8≤ L | 合计Total |
| 乔木层Arbor layer | 37 | 60 | 43 | 11 | 5 | 0 | 156 |
| 灌木层Shrub layer | 8 | 115 | 123 | 43 | 16 | 1 | 306 |
| 草本层Herb layer | 15 | 33 | 33 | 21 | 8 | 0 | 110 |
结合表4—表7分析可知,乔木层生态位重叠数值的平均值为0.117,生态位重叠值主要集中分布于0.01与0.3之间,其中,有38.46%对介于0.01与0.1之间,生态位重叠指数值介于0.1与0.3之间的有27.57%;灌木层生态位重叠指数的平均值为0.175,主要集中分布于0.01与0.5之间,37.58%的生态位重叠指数值介于0.01与0.1之间,生态位重叠指数值介于0.1与0.3之间的有40.20%,生态位重叠指数值介于0.3与0.5之间的有14.05%;草本层生态位重叠数值的平均值为0.191,主要分布于0.01与0.5之间,其中30%的生态位重叠指数值介于0.01与0.1之间,生态位重叠指数值介于0.1与0.3之间的有30%,生态位重叠指数值介于0.3与0.5之间的有19.09%。
3.2 种间联结种间联结是对物种在不同生境中相互影响、相互作用所形成的有机联系的反映,它表示种间相互吸引或排斥的性质[1]。
3.2.1 乔木层乔木层13个优势种的联结系数AC半矩阵图如图1所示。联结系数AC≥0.26的有21对,占乔木层总对数的26.9%;联结系数AC≤-0.37的有22对,占总对数的28.2%;联结系数-0.37 < AC < 0.26的种对有35对,占总对数的44.9%。其中有8对呈极显著正关联,有2对为显著正关联。

|
| 图 1 乔木层13个优势种种间联结性半矩阵图 Fig.1 Semi-matrices of interspecific association for 13 dominant species in tree layer *表示3.841 < χ2 < 6.635(P < 0.05),**表示χ2>6.635 |
灌木层18个优势种的联结系数AC半矩阵图如图2所示。联结系数AC≥0.33的有19对,占乔木层总对数的12.4%;联结系数AC≤-0.33的有22对,占总对数的14.4%;联结系数-0.33 < AC < 0.33的种对有112对,占总对数的73.2%。其中有3对呈极显著正关联,有5对为显著正关联,有3对显著负关联。

|
| 图 2 灌木层18个优势种种间联结性半矩阵图 Fig.2 Semi-matrices of interspecific association for 18 dominant species in shrub layer |
草本层11个优势种的联结系数AC半矩阵图如图3所示。联结系数AC≥0.33的有12对,占乔木层总对数的21.8%;联结系数AC≤-0.33的有5对,占总对数的9.1%;联结系数-0.33 < AC < 0.33的种对有38对,占总对数的69.1%。其中有2对呈极显著正关联,有2对为显著正关联,有1对显著负关联。

|
| 图 3 灌木层11个优势种种间联结性半矩阵图 Fig.3 Semi-matrices of interspecific association for 11 dominant species in Herbaceous layer |
利用CANOCO 5.0对研究区内42个研究对象和环境数据做典范对应分析。首先,对42×80维重要值矩阵进行趋势对应分析(DCA排序),通过DCA排序可得Lengths of gradient的第一轴值为3.963,约为4,即可进行CCA排序。结合6×80维环境矩阵进行CCA排序,如表8所示,蒙特卡洛检验结果表明,所得的典范轴都是极显著的(P < 0.01)。
| 环境因子 Environmental factors | 第一轴Axis 1 | 第二轴Axis 2 | 第三轴Axis 3 | 第四轴Axis 4 |
| 坡位 Position,Pos | 0.1833 | -0.4121 | -0.0315 | -0.2470 |
| 坡度 Slope,Slo | -0.6314 | 0.1184 | -0.3605 | 0.0847 |
| 坡向 Aspect,Asp | 0.2979 | -0.4740 | -0.2450 | 0.1865 |
| 海拔 Elevation,Ele | -0.7944 | -0.1412 | 0.1235 | 0.0830 |
| 土壤厚度 Soil depth,Sod | 0.2361 | -0.0297 | 0.2474 | 0.3302 |
| 腐殖质厚度 Humus depth,Hud | -0.3802 | -0.2400 | 0.1866 | 0.1893 |
| CCA排序概要 Summary of CCA ordination | ||||
| 特征值 Eigenvalues | 0.228 | 0.161 | 0.080 | 0.061 |
| 物种-环境相关 Species-environment correlations | 0.807 | 0.638 | 0.610 | 0.626 |
| 物种环境关系的方差累积比例/%Cumulative percentage variance of species-environment | 38.84 | 67.01 | 80.77 | 89.44 |
| 所有典范轴的显著性检验Test of significance of all canonical axes | 0.002 |
由表8可知,CCA排序中4个轴累积解释了物种-环境关系的89.44%,其中前两轴的累积解释度达67.01%,说明CCA前两轴已能较好地反映出研究区群落物种与环境之间的关系。6个环境因子中与轴1相关性最高的环境因子为海拔,其次是坡度、腐殖质厚度、坡向;与第2轴相关性最高的环境因子为坡向,其次为坡位、腐殖质厚度。如图4所示,海拔、坡度沿CCA1轴由左向右逐渐降低,而土壤厚度沿第1轴从左往右增加逐渐增加。坡位、坡向沿CCA2轴从下向上逐渐减小,即坡位沿CCA第2轴由下向上依次为上坡位、中坡位与下坡位,坡向沿CCA第2轴自下向上依次为阳坡、半阳坡、半阴坡、阴坡。

|
| 图 4 物种和环境因子的CCA二维排序图 Fig.4 CCA biplot of species and environmental factors 图中仅显示了重要值在1以上的物种,共计42个(物种序号参见表3);Asp:坡向;Ele:海拔;Hud:腐殖质厚度;Pos:坡位; Slo:坡度;Sod:土壤厚度 |
不同物种对环境的要求存在差异,因而在CCA排序图中具有不同的分布差异。排序图位于纵轴左侧的物种分布主要受到海拔和坡度的影响,其中甜槠(物种7,主要分布于海拔600—1200 m,坡度38°—55°地区)、木荷(物种4,主要分布于海拔550—1200 m,坡度38°—50°地区)与木荷幼苗(物种30)、短柄枹(物种10,主要分布于海拔650—1200 m,坡度38°—55°地区)、三花悬钩子(物种41,主要分布于海拔620—980 m,坡度40°—45°地区)、粗叶悬钩子(物种37,主要分布于海拔550—1050 m,坡度38°—45°地区)在海拔较高、坡度较陡的区域分布较多;而位于纵轴右侧的物种则在海拔较低、坡度较为平缓的区域分布较广,其中以湿地松(物种6,主要分布于海拔120—280 m,坡度20°—30°地区)、苦竹(物种25,主要分布于海拔130—480 m,坡度23°—35°地区)、野菊(物种42,主要分布于海拔120—530 m,坡度28°—33°地区)最为明显。排序图位于横轴下方的物种分布主要受到坡位和坡向的影响,其中菝葜(物种23)、马尾松(物种6)、黄檀(13)主要分布在上坡位的半阳面和阳面;而位于横轴上方的物种则主要分布于下坡位的阴面或半阴面,其中毛竹(物种3)、蛇莓(物种40)、野菊(物种42)、湿地松(物种6)、苦竹(物种25)最为明显。
4 结论与讨论本研究通过对江山市公益林优势种的生态位、种间联结性及CCA排序进行分析,发现优势种群中杉木、檵木、紫萁的重要值分别在乔木层、灌木层、草本层中占有优势地位;群落各层物种间均存在不同程度的生态位重叠现象;并且通过CCA排序表明不同物种对环境的要求存在差异,优势种分布主要受海拔、腐殖质厚度、坡向及坡位的影响较为明显。
以往的研究常常将种间关系割裂单独分析,如生态位理论,种间联结性或CCA协变量矩阵偏典范对应分析等。特别是利用CCA排序分析环境因子中,常常会出现未能得到相应解释的部分,说明一些未知的环境因子的空间过程在种群空间结构的形成过程中起了重要作用。空间因子和环境因子未能解释的部分主要反映了群落自身的相互作用。因此,本研究充分利用种间关系研究方法来探究环境对物种与种间关系的影响能力。
本研究表明,重要值大的物种其生态位宽度大,反之亦然,如本研究中的马尾松、杉木、檵木、杜鹃等物种,此类物种对环境资源利用能力较强,与其他物种的生态位重叠程度高,在CCA排序图中大致分布于物种集中区域,反之亦然。如,甜槠、黄檀、三花悬钩子、野菊等物种,生态位重叠程度低,且在CCA排序图中分布在外围;然而也有部分植物种虽然重要值大,但生态位宽度值并不大,如本研究中的毛竹、湿地松等物种,这类物种与其他物种的生态位重叠程度比同等重要值的物种低,在CCA排序图中分布偏离物种集中区,且与其他物种的种间联结性大致呈不相关或负相关,这表明此类物种对环境因子的要求较为独特,与其他物种间竞争较弱,从而在某一特定环境下占据优势,导致该类物种重要值高但生态位宽度小;除此之外,有部分物种重要值不高,但其生态位宽度大,如山合欢、乌饭、山苍子、淡竹叶等,这类物种虽然重要值不高,但与其他物种的生态位重叠程度较同等重要值的物种来说高,在CCA排序图中大致分布于物种集中区域,且与其他物种的种间联结性大致呈正相关,说明其分布较广且与CCA排序图中距离较近的物种间关于环境资源的竞争强,但由于其本身对环境资源利用能力较弱,因而在广泛的分布范围内物种数量少,导致其重要值低但生态位重叠值却高。
影响植物种在群落中分布的因素较为复杂,而上述分析基于优势种重要值和生态位宽度的分类,通过结合生态位重叠值、种间联接及CCA排序3方面的综合分析对物种分布的现状进行分析,得到以下规律:生态位宽度较小的物种由于分布不够广泛、对资源利用专性较强,与其它物种间的生态位重叠较小,在CCA排序图中偏离物种集中区,如本研究中的湿地松、毛竹;相反,生态位宽度较大的物种则与其它物种的生态位重叠值较大,分布于CCA排序图中物种集中区。对于同一物种来说,若与其它物种间没有显著的联结性,则物种间的生态位重叠值越高,物种在CCA排序图上的距离越近;若存在较为显著的正联结性,则正联结性越显著,物种在CCA排序图上的距离越近;若存在较为显著的负联结性,则负联结性越显著,物种在CCA排序图上的距离越远。
| [1] | 王乃江, 张文辉, 陆元昌, 范少辉, 王勇. 陕西子午岭森林植物群落种间联结性. 生态学报, 2010, 30(1): 67-78. |
| [2] | Phillips O L, Vargas P N, Monteagudo A L, Cruz A P, Zans M E C, Sánchez W G, Yli-Halla M, Rose S. Habitat association among Amazonian tree species: a landscape-scale approach. Journal of Ecology, 2003, 91(5): 757-775. |
| [3] | 程瑞梅, 王瑞丽, 刘泽彬, 封晓辉, 王晓荣, 肖文发. 三峡库区栲属群落主要乔木种群的种间联结性. 林业科学, 2013, 49(5): 36-42. |
| [4] | Motzkin G, Eberhardt R, Hall B. Vegetation variation across Cape Cod, Massachusetts: environmental and historical determinants. Journal of Biogeography, 2002, 29(10-11): 1439-1454. |
| [5] | 史作民, 程瑞梅, 刘世荣. 宝天曼落叶阔叶林种群生态位特征. 应用生态学报, 1999, 10(3): 265-269. |
| [6] | 赵永华, 雷瑞德, 何兴元, 贾夏. 秦岭锐齿栎林种群生态位特征研究. 应用生态学报, 2004, 15(6): 913-918. |
| [7] | Zhang W, Li C H, Jia X P, Chen P M. Research on spatial Interpolation methods of Macrobenthic biomass. Marine Science Bulletin, 2010, 29(3): 351-356. |
| [8] | Hemandez-Stefanoni J L, Pineda J B, Valades-Valadez G. Comparing the use of indigenous knowledge with classification and ordination techniques for assessing the species composition and structure of vegetation in a Tropical Forest. Environmental Management, 2006, 37(5): 686-702. |
| [9] | 何惠琴, 李绍才, 孙海龙, 刘世昌, 熊文兰. 锦屏水电站植被数量分类与排序. 生态学报, 2008, 28(8): 3706-3712. |
| [10] | 许莎莎, 孙国钧, 刘慧明, 龚雪平, 郝媛媛, 张立勋. 黑河河岸植被与环境因子间的相互作用. 生态学报, 2011, 31(9): 2421-2429. |
| [11] | 胡贝娟, 张钦弟, 张玲, 毕润成. 山西太岳山连翘群落优势种种间关系. 生态学杂志, 2013, 32(4): 845-851. |
| [12] | 伊力塔, 严晓素, 余树全, 包春泉, 屠永海, 潘昌尧. 浙江省不同森林类型林分健康指标体系. 南京林业大学学报: 自然科学版, 2012, 36(1): 145-148. |
| [13] | 钱逸凡, 伊力塔, 张超, 余树全, 沈露, 彭冬琴, 郑超超. 浙江省中部地区公益林生物量与碳储量. 林业科学, 2013, 49(5): 17-23. |
| [14] | 余树全, 李翠环. 千岛湖水源涵养林优势树种生态位研究. 北京林业大学学报, 2003, 25(2): 18-23. |
| [15] | 巨天珍, 张彦文, 葛建团, 张宋智, 宋晓伟, 师贺雄, 米彩艳. 小陇山国家级自然保护区油松群落生态位研究. 干旱区资源与环境, 2013, 27(4): 174-178. |
| [16] | Hurbert H. A coefficient of interspecific association. Ecology, 1969, 50(1): 1-9. |
| [17] | 覃林. 统计生态学. 北京: 中国林业出版社, 2009: 84-97. |
| [18] | 栾青杉, 孙军, 宋书群, 沈志良, 俞志明. 长江口夏季浮游植物群落与环境因子的典范对应分析. 植物生态学报, 2007, 31(3): 445-450. |
| [19] | 孟睿, 何连生, 过龙根, 席北斗, 李中强, 舒俭民, 刁晓君, 李必才. 长江中下游草型湖泊浮游植物群落及其与环境因子的典范对应分析. 环境科学, 2013, 34(7): 2588-2596. |
| [20] | 钱逸凡, 伊力塔, 胡军飞, 张超, 余树全, 沈露, 彭东琴. 普陀山主要植物种生态位特征. 生态学杂志, 2012, 31(3): 561-568. |