 2015, Vol. 35
2015, Vol. 35文章信息
- 马志良, 高顺, 杨万勤, 吴福忠, 谭波, 张玺涛
- MA Zhiliang, GAO Shun, YANG Wanqin, WU Fuzhong, TAN Bo, ZHANG Xitao
- 亚热带常绿阔叶林6个常见树种凋落叶在不同降雨期的分解特征
- Litter decomposition of six common tree species at different rainy periods in the subtropical region
- 生态学报, 2015, 35(22): 7553-7561
- Acta Ecologica Sinica, 2015, 35(22): 7553-7561
- http://dx.doi.org/10.5846/stxb201403180470
-
文章历史
- 收稿日期: 2014-03-18
- 网络出版日期: 2015-04-20






凋落物是森林生态系统物质和能量流动的关键环节之一,在维持森林生态系统生产力、净碳储量、土壤有机质的形成、森林植被群落演替等方面具有不可替代的作用和地位,其分解由降水淋溶、动物的咀嚼和啃食、土壤干湿交替和冻融循环和生物代谢等相互联系的物理、化学和生物作用共同完成[1]。我国亚热带常绿阔叶林区具有雨热同季的气候特点,明显的季节性干湿交替可通过增加凋落物的物理破碎[2]和调控生物分解者活动[3]等影响凋落物分解。亚热带常绿阔叶林凋落物大量发生在秋末冬初[4, 5],新鲜凋落物虽然具有较高的易分解组分含量[6],但由于冬季降水少、气温低,分解者的活性较低,造成凋落物分解相对缓慢[7]。随着凋落物分解的进行,季节性降雨期的来临使得水热条件得到明显改善,分解者活性和淋溶强度显著提高,在一定程度上促进了凋落物分解[8],但经历一个旱季分解的凋落物,基质质量随着大量易分解组分的损失明显下降,一定程度上又限制了凋落物分解进程[9]。可见,季节性干湿交替环境下凋落物分解过程并不清晰。然而,近年来开展的相关研究多关注亚热带常绿阔叶林凋落物产量[10]、总体分解特征如年分解速率[11]和养分释放[12]、以及分解者活动[4, 13]等方面,有关季节性降雨对凋落物分解的影响研究相对较少。更为重要的是,气候变化情景下季节性降雨格局的改变可能使季节性降雨对凋落物分解的影响更为重要。
四川盆地亚热带常绿阔叶林夏季高温多雨,季节性降雨期长达5—6个月。由于相对较好的水热环境,凋落物在季节性降雨期间可能具有更为明显的分解特征,并受到不同树种凋落物质量的影响,但相关过程缺乏必要关注。因此,本文以四川盆地亚热带常绿阔叶林区6种常见树种马尾松(Pinus massoniana)、柳杉(Cryptomeria fortunei)、杉木(Cunninghamia lanceolata)、香樟(Cinnamomum camphora)、红椿(Toona ciliata)、麻栎(Quercus acutissima)凋落叶为研究对象,根据降雨特征,采用凋落物袋法研究了季节性降雨期间凋落叶分解过程及其对季节性干湿交替的响应,以期为更好的认识亚热带常绿阔叶林区凋落物分解过程提供基础理论依据。
1 材料与方法 1.1 研究区概况凋落物分解实验在四川省崇州市桤泉镇四川农业大学现代农业研发基地进行。该基地坐落于四川省川西平原西部,地理坐标103°49'E,30°55'N,海拔516 m,属四川盆地亚热带湿润季风气候。年平均气温16.0 ℃,年平均降雨量1015.2 mm,降雨主要集中在5—9月,降雪稀少。年平均日照时数为1161.5 h,平均无霜期为283 d,土壤类型为老冲积黄壤。研究区域内无乔木、灌木等木本植物,草本植物主要有青蒿(Artemisia annua)、空心莲子草(Alternanthera philoxeroides)、狗尾草(Setaria viridis)、稗草(Echinochloa crusgalli)、虮子草(Leptochloa panicea)、牛繁缕(Malachium aquaticum)、藜(Chenopodium album)、野苋菜(Amaranthus viridis)等。
1.2 试验方法 1.2.1 试验设计和样品处理2012年10月底在四川盆地典型生态系统用尼龙网收集马尾松、柳杉、杉木、香樟、红椿、麻栎的新鲜凋落叶。收集好的6种凋落叶带回实验室,经自然风干后,每个树种准确称取10 g装入凋落物袋内(规格为20 cm × 20 cm,网孔大小为底部0.5 mm,表面层1 mm)并标记,每种凋落叶分别装15袋,共计90袋。另外分别称取相同质量(10 g)的6种树种凋落叶各3份,在65 ℃下烘干至恒重,测定含水量,用于推算放置在凋落物袋内凋落叶的初始干质量(马尾松8.71 g、柳杉8.89 g、杉木8.89 g、香樟8.78 g、红椿8.78 g、麻栎8.78 g)。烘干的凋落叶样品粉碎后用于初始全C、N、P、木质素和纤维素含量的测定。其中,全C采用重铬酸钾氧化-外加热法测定(LY/T 1237—1999);样品全N和全P待测液用浓H2SO4-H2O2消煮法制备(NY/T 2017—2011),全N采用半微量凯氏定氮法测定(LY/T 1228—1999),全P采用钼锑抗比色法测定(LY/T 1270—1999);木质素和纤维素含量采用酸性洗涤纤维法测定[14]。6种凋落叶分解前基质特征见表1。
| 物种 Species | C/ (g/kg) | N/ (g/kg) | P/ (g/kg) | C/N | C/P | N/P | 木质素 Lignin/% | 纤维素 Cellulose/% | 木质素/N Lignin/N |
| 表中同列不同的小写字母表示不同类型凋落物之间差异显著, P < 0.05 | |||||||||
| 马尾松 P. massoniana | 441.99±10.83a | 10.72±0.33abc | 0.81±0.03c | 41.23±0.88a | 543.52±29.41a | 13.19±0.83a | 34.87±1.10b | 20.03±1.26cd | 32.53±1.30bc |
| 柳杉 C. lanceolata | 425.12±5.51a | 11.99±0.36ab | 0.87±0.06bc | 35.48±0.65a | 489.87±38.73a | 13.82±1.33a | 36.23±0.98b | 25.03±1.33a | 30.26±1.68c |
| 杉木 C. fortunei | 393.95±2.05b | 10.19±1.19c | 0.76±0.06c | 39.25±7.54a | 518.16±60.46a | 13.36±1.58a | 38.92±0.58a | 24.39±2.69ab | 38.51±4.03a |
| 香樟 C. camphora | 379.11±9.61bc | 10.51±0.87bc | 1.23±0.12a | 36.19±2.18a | 308.07±30.08b | 8.54±1.08b | 38.38±1.02a | 22.09±1.87abc | 37.02±3.13ab |
| 红椿 T. ciliata | 350.36±2.52c | 12.23±1.49a | 0.98±0.13b | 28.95±3.73b | 363.29±49.52b | 12.78±2.95a | 28.75±0.95c | 17.65±1.65d | 23.84±3.92d |
| 麻栎 Q. acutissima | 428.59±12.50a | 11.06±0.66abc | 0.78±0.06c | 38.78±1.20a | 552.24±50.29a | 14.26±1.63a | 38.80±0.87a | 20.35±0.67bcd | 35.14±1.37abc |
设置3块样地作为重复,于2013年1月18日去除样地土壤表面的植物和凋落物,将上述凋落物袋平铺于地表,每种凋落物袋在每块样地内放置5袋。样品埋设后,在试验地地表凋落物袋内设置一个纽扣式温度记录器(iButton DS1923-F5,Maxim/Dallas Semiconductor,Sunnyvale,USA),设定为每120 min读取1次温度数据,自动记录试验期间地表的温度变化,试验期间的日平均温度变化特征如图1。根据崇州市多年降雨资料,将试验时间划分为微量降雨期(MRS: 2013年1月18日—2013年2月19日)、春季少雨期(SRS: 2013年2月19日—2013年4月22日)、雨季前期(ERS: 2013年4月22日—2013年8月19日)、雨季后期(LRS: 2013年8月19日—2013年10月22日)和冬季少雨期(WRS: 2013年10月22日—2014年1月18日)5个时期。试验期间的降水特征见图1和表2。其中,2013年1月18日—4月22日及2013年10月22日—2014年1月18日降雨不集中、降雨量小(总降雨量为98 mm),占试验期间总降雨量的8.13%;试验地降雨主要发生在2013年4月22日—10月22日(总降雨量为1106.9 mm),占试验期间总降雨量的91.87%,表现为明显的雨季和旱季。具体采样时间为2013年2月19日、4月22日、8月19日、10月22日和2014年1月18日,每次从每块样地内采集6种凋落物袋各1袋,带回实验室。小心去除表面泥土和新长入的细根,于65 ℃烘干至恒重,称重后用于质量损失数据的计算。

|
| 图 1 试验期间地表温度动态和月实际降水量 Fig.1 Dynamics of average surface temperature and month actual precipitation during the experiment 降雨量资料从http://sc.weather.com.cn/qxfw/index.shtml中获得 |
|
采样时期 Sampling time | 小雨 Light rain | 中雨 Moderate rain | 大雨 Heavy rain | 暴雨 Rainstorm | 大暴雨 Downpour | 降雨量 Precipitation/mm |
| 小雨:24h降水量小于10 mm;中雨:24h降雨量在10.1—25 mm之间;大雨:24h降雨量在25.1—50 mm之间;暴雨:24h降雨量在50.1—100 mm之间;大暴雨:24h降雨量在100.1—200 mm之间; MRS: 微量降雨期Micro rainy stage;SRS: 春季少雨期Spring rainy stage;ERS: 雨季前期Early stage of rainy season;LRS: 雨季后期Late stage of rainy season;WRS: 冬季少雨期Winter rainy stage, | ||||||
| MRS | 3 | 0 | 0 | 0 | 0 | 2.3 |
| SRS | 14 | 2 | 0 | 0 | 0 | 58.8 |
| ERS | 29 | 9 | 1 | 5 | 1 | 925.2 |
| LRS | 19 | 6 | 3 | 0 | 0 | 181.7 |
| WRS | 22 | 0 | 0 | 0 | 0 | 36.9 |
凋落物质量残留率:

各时期凋落物质量损失贡献率:
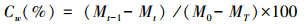
各时期凋落物质量损失速率(以天计):
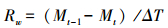
各时期凋落物质量损失:
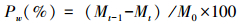
式中,M0为凋落物分解袋埋置前烘干凋落物质量(g);Mt为各采样时间凋落物内凋落物的残留量(g);(Mt-1-Mt)为相邻两次采集凋落物分解袋的残留量差(g);ΔT为相邻两次采样时间的间隔天数(d);MT为最后一次采样时凋落物袋内凋落物残留量(g)。
1.2.3 数据统计与分析试验数据统计与分析采用Excel 2003和SPSS 20.0完成。使用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)比较6种凋落叶初始基质质量与各采样时期的质量损失和质量损失速率;采用Pearson相关分析方法分析凋落物质量损失与初始基质质量之间的关系;采用线性回归方法分析凋落物质量损失与降雨量和温度的之间的关系。制图使用Origin 9.0 完成。
2 结果与分析 2.1 凋落叶质量损失6种凋落叶质量损失如图2和图3。6种凋落叶分解过程质量残留规律一致,均表现为随时间的进行凋落叶干物质残留率不断减少。经历1a的分解,6种凋落叶质量残留率大小顺序依次为:红椿(27.90%) < 柳杉(41.39%) < 杉木(48.93%) < 麻栎(49.62) < 马尾松(68.82%) < 香樟(72.23%),红椿凋落叶质量残留率均显著(P < 0.01)低于其它5种凋落叶(图2)。

|
| 图 2 6个树种凋落叶分解过程中质量残留率的动态变化 Fig.2 Dynamics of mass remaining of six species leaf litter decomposing 图中横线表示标准误差;**表示不同树种之间差异显著(P <0.01); PM:马尾松 Pinus massoniana;CF: 柳杉Cryptomeria fortunei;CL: 杉木Cunninghamia lanceolata;CC: 香樟Cinnamomum camphora;TC: 红椿Toona ciliate;QA: 麻栎Quercus acutissima |

|
| 图 3 各采样时期6个树种凋落叶质量损失的贡献率 Fig.3 Contribution rates of six species litter mass loss during different sampling period MRS: 微量降雨期 Micro rainy stage;SRS: 春季少雨期 Spring rainy stage;ERS: 雨季前期 early stage of rainy season;LRS: 雨季后期 late stage of rainy season;WRS: 冬季少雨期 Winter rainy stage |
总体来看,在1a的分解中,整个雨季(ERS和LRS)对6种凋落叶质量损失的贡献率(69.73%—89.68%)均明显大于旱季(MRS、SRS和WRS)的贡献率(10.32%—30.27%)(图3)。值得注意的是,阔叶树种香樟、红椿、麻栎凋落叶在旱季也有较大的质量损失(分别占总质量损失17.96%、18.88%、30.27%),均高于针叶树种马尾松、柳杉、杉木凋落叶在旱季的质量损失(分别占总质量损失的12.07%、13.42%、10.32%)。
2.2 凋落叶质量损失速率由图4可见,6种凋落叶在不同分解关键时期质量损失速率差异差异显著(P < 0.05)。总体来看,凋落叶在雨季(ERS和LRS)的质量损失速率显著(P < 0.05)均高于旱季(MRS、SRS和WRS)。其中,马尾松和香樟凋落叶在各时期中质量损失速率表现为ERS > LRS > WRS > MRS > SRS;柳杉和杉木凋落叶表现为LRS > ERS > WRS > MRS > SRS;红椿凋落叶表现为ERS > LRS > MRS > SRS > WRS;麻栎凋落叶表现为LRS > ERS > WRS > SRS > MRS。

|
| 图 4 各分解时期6个树种凋落叶质量损失速率 Fig.4 Leaf litter mass loss rates of six species during different sampling period |
凋落叶各分解时期质量损失及速率与初始基质质量的相关系数于表3中列出。由表3可见,凋落叶质量损失与初始C、木质素含量及C/N、木质素/N极显著(P < 0.01)负相关,与N含量极显著(P < 0.01)正相关。凋落叶前期分解过程(MRS、SRS和ERS)质量损失速率与初始C、木质素、纤维素含量及C/N、木质素/N极显著(P < 0.01)负相关,与N含量极显著(P < 0.01)正相关。
|
项目 Item | 采样时间 Sampling time | C/ (g/kg) | N/ (g/kg) | P/ (g/kg) | C/N | C/P | N/P | 木质素 Lignin/% | 纤维素 Cellulose/% | 木质素/N Lignin/N |
| *表示存在显著相关性 (P < 0.05);**表示存在极显著相关性(P < 0.01) | ||||||||||
| 质量损失 | 2013-02-19 | -0.719** | 0.665** | 0.224 | -0.866** | -0.480* | 0.013 | -0.877** | -0.745** | -0.815** |
| Mass lossPw/% | 2013-04-22 | -0.644** | 0.662** | 0.150 | -0.816** | -0.389 | 0.095 | -0.819** | -0.793** | -0.783** |
| 2013-08-19 | -0.546* | 0.855** | 0.008 | -0.874** | -0.292 | 0.276 | -0.884** | -0.447 | -0.909** | |
| 2013-10-22 | -0.494* | 0.750** | -0.222 | -0.752** | -0.090 | 0.464 | -0.686** | -0.191 | -0.729** | |
| 2014-01-18 | -0.402 | 0.754** | -0.295 | -0.699** | 0.005 | 0.547* | -0.584* | -0.156 | -0.672** | |
| MRS | -0.719** | 0.665** | 0.224 | -0.866** | -0.480** | 0.013 | -0.877** | -0.744** | -0.816** | |
| 质量损失速率 | SRS | -0.501* | 0.612* | 0.042 | 0.695** | -0.243 | 0.195 | -0.686** | -0.801** | -0.686** |
| Mass loss rates | ERS | -0.494* | 0.861** | -0.045 | -0.829** | -0.234 | 0.324 | -0.833** | -0.276 | -0.882** |
| Rw/(mg/d) | LRS | -0.065 | 0.102 | -0.573* | -0.070 | 0.381 | 0.586* | 0.120 | 0.459 | 0.065 |
| WRS | 0.358 | 0.109 | -0.357 | 0.154 | 0.420 | 0.435 | 0.385 | 0.140 | 0.176 | |
凋落叶质量损失及其速率均与降雨量极显著(P < 0.01)正相关(R2分别为0.65和0.32),说明6种凋落叶分解过程受降雨量影响极显著(P < 0.01)(图5)。同时,凋落叶分解过程受平均温度的影响,在凋落物分解过程中质量损失及其速率均与平均温度极显著(P < 0.001)相关(R2分别为0.49和0.41)(图6)。

|
| 图 5 凋落叶各分解时期质量损失和质量损失速率与采样期间降雨量的相关关系 Fig.5 Correlation among mass loss, mass loss rates of each decomposition stage and precipitation during each sampling period |

|
| 图 6 凋落叶各分解时期质量损失和质量损失速率与采样期间平均温度的相关关系 Fig.6 Correlation among mass loss, mass loss rates of each decomposition stage and average temperature during each sampling period |
普遍认为,凋落物质量损失随着降雨量的增加而增加[15, 16, 17]。本项研究表明,亚热带常绿阔叶林6种常见树种第一年的凋落物质量损失为27.77%—72.10%,其中,69.73%—89.68%发生在雨季(RES和LRS),即雨热同季的4月底—10月中旬。这与Anaya等对热带森林生态系统凋落物分解的研究结果一致[15]。可能的原因包括两方面:一方面,雨季的降雨可促进凋落物化学成分淋溶,降雨量越大,表层凋落物淋溶越快[15],降雨对亚热带森林生态系统凋落物分解有直接的正效应;另一方面,在亚热带湿润气候区,雨热同季的气候变化有利于分解者(土壤动物和微生物)活动[8],森林地表凋落物层具有强大的持水能力[18],雨季更为频繁的降雨和较大的降雨量所提供良好的土壤水分条件持续时间更久[19],这将更有利于生物分解者的繁殖、生长和采食活动,从而增加凋落物的分解速率[20]。此外,凋落叶质量损失及其速率与降雨量及温度的相关分析结果表明,凋落叶质量损失及其速率均与降雨量和温度呈极显著(P < 0.01)正相关关系。中国长期凋落物分解实验研究结果显示,气候是控制凋落物分解的主要因子,年均降雨量对凋落物分解影响显著[21]。我国中东部不同气候带森林凋落物分解速率也表现为随温度和降雨量的增加而增大[10]。本研究结果与此一致。
经历3个月旱季分解后,6种物种凋落叶在雨季的质量损失急速加快,原因之一可能是凋落叶在经过初期旱季(MRS和SRS)分解,化学成分发生变化,同时积累了一定的可溶性物质,在随后的季节性降雨期通过淋溶作用的方式爆发式释放[2]。雨季强大的降雨脉冲对凋落物表面造成的物理破碎也可能是加速其分解的另一个原因[22]。而经历一个季节性降雨期分解的凋落叶可能只剩余更加难分解的组分(如木质素和纤维素等),导致WRS时期的凋落叶质量损失和质量损失速率均显著(P < 0.01)降低(图4)。然而,不同树种的凋落物质量损失对降雨格局的响应存在很大差异。
3.2 基质质量与不同关键时期凋落物质量损失的关系与树种相关的凋落物基质质量决定着生态系统水平的凋落物分解[23]。初始基质质量中的C、N、木质素浓度及C/N、木质素/N能够作为凋落物分解速率的预测指标[16]。已有的多数研究表明,凋落物初始N含量高会加速分解,而木质素含量高将延缓分解[24, 25]。本研究中,红椿凋落叶的N含量最高,木质素含量最低,因而分解最快,历经1年分解后,质量残留率仅为27.90%,显著(P < 0.05)低于其它5种凋落叶。而香樟和麻栎凋落叶N含量较低,木质素含量较高,且叶表面蜡质程度较高,分解较慢。相关分析结果表明,凋落叶质量损失与初始C、木质素含量及C/N、木质素/N极显著(P < 0.01)负相关,与初始N含量极显著(P < 0.01)正相关。凋落叶初期分解过程(MRS、SRS和ERS)质量损失速率与初始C、木质素、纤维素含量及C/N、木质素/N极显著(P < 0.01)负相关,与初始N含量极显著(P < 0.01)正相关(表3)。这也验证了初始基质质量中N含量、C/N、木质素/N等是预测凋落物在亚热带常绿阔叶林区早期分解过程的良好指标这一结论[25]。
本研究还发现,不同树种凋落叶在不同关键时期质量损失及其速率具有显著差异(图2和图4)。凋落物自身特性(易分解和难分解成分)和降雨量交互作用可能是控制凋落物在不同关键时期分解的主要因子[26]。不同树种凋落叶的组成成分存在显著差异(表1),造成在不同分解阶段对降雨的响应不同。凋落物在分解初期主要向环境中释放可溶性物质和矿质养分,可溶性物质(易分解成分)含量高的凋落物在雨季更易受降雨淋溶作用的促进而具有更大的质量损失速率[15, 27],而纤维素、木质素等难分解物质含量高的凋落叶在分解初期由于受到细胞壁上木质素-纤维素复合体的保护[28],降雨不能直接作用于凋落物易分解成分,从而造成质量损失在时间上的后延。值得注意的是,在旱季少量降雨情景下,阔叶树种凋落叶的质量损失(17.96%—30.27%)明显高于针叶树种凋落叶(10.32%—13.42%) (图3),这说明具有较大叶面积的阔叶树种凋落叶分解对旱季少量降雨的响应比针叶树种更为敏感。
综上所述,四川盆地亚热带常绿阔叶林区凋落叶分解主要发生在雨季,温湿度对凋落叶分解过程影响显著。在未来全球气候变化过程中,亚热带常绿阔叶林区夏季气温升高、降雨量增加[29],季节性降雨期间具有较快的凋落物分解速率,提高了林地养分归还速度,这将能够为正处于生长季的植物提供更多的有效养分,促进森林生态系统的物质循环。
| [1] | 杨万勤, 邓仁菊, 张健. 森林凋落物分解及其对全球气候变化的响应. 应用生态学报, 2007, 18(12): 2889-2895. |
| [2] | 赵红梅, 黄刚, 马健, 李彦, 周丽. 荒漠区地表凋落物分解对季节性降水增加的响应. 植物生态学报, 2012, 36(6): 471-482. |
| [3] | 武海涛, 吕宪国, 杨青, 姜明. 土壤动物主要生态特征与生态功能研究进展. 土壤学报, 2006, 43(2): 314-323. |
| [4] | 王文君, 杨万勤, 谭波, 刘瑞龙, 吴福忠. 四川盆地亚热带常绿阔叶林土壤动物对几种典型凋落物分解的影响. 生态环境学报, 2013, 22(9): 1488-1495. |
| [5] | 张德强, 叶万辉, 余清发, 孔国辉, 张佑昌. 鼎湖山演替系列中代表性森林凋落物的研究. 生态学报, 2000, 20(6): 938-944. |
| [6] | Fröberg M, Berggren Kleja D, Hagedorn F. The contribution of fresh litter to dissolved organic carbon leached from a coniferous forest floor. European Journal of Soil Science, 2007, 58(1): 108-114. |
| [7] | Austin A T, Vivanco L. Plant litter decomposition in a semi-arid ecosystem controlled by photodegradation. Nature, 2006, 442(7102): 555-558. |
| [8] | Anaya C A, Jaramillo V J, Martínez-Yrízar A, García-Oliva F. Large rainfall pulses control litter decomposition in a tropical dry forest: evidence from an 8-year study. Ecosystems, 2012, 15(4): 652-663. |
| [9] | Prescott C E. Litter decomposition: what controls it and how can we alter it to sequester more carbon in forest soils? Biogeochemistry, 2010, 101(1-3): 133-149. |
| [10] | 王健健, 王永吉, 来利明, 赵学春, 王飞, 申国珍, 赖江山, 鲁洪斌, 赵春强, 郑元润. 我国中东部不同气候带成熟林凋落物生产和分解及其与环境因子的关系. 生态学报, 2013, 33(15): 4818-4825. |
| [11] | 季晓燕, 江洪, 洪江华, 马元丹. 亚热带3种树种凋落叶厚度对其分解速率及酶活性的影响. 生态学报, 2013, 33(6): 1731-1739. |
| [12] | 仲米财, 王清奎, 高洪, 于小军. 中亚热带主要树种凋落叶在杉木人工林中分解及氮磷释放过程. 生态学杂志, 2013, 32(7): 1653-1659. |
| [13] | 王文君, 杨万勤, 谭波, 刘瑞龙, 吴福忠. 四川盆地亚热带常绿阔叶林不同物候期凋落物分解与土壤动物群落结构的关系. 生态学报, 2013, 33(18): 5737-5750. |
| [14] | Rowland A P, Roberts J D. Lignin and cellulose fractionation in decomposition studies using acid-detergent fibre methods. Communications in Soil Science and Plant Analysis, 1994, 25(3-4): 269-277. |
| [15] | Anaya C A, García-Oliva F, Jaramillo V J. Rainfall and labile carbon availability control litter nitrogen dynamics in a tropical dry forest. Oecologia, 2007, 150(4): 602-610. |
| [16] | Swift M J, Heal O W, Anderson J M, Anderson D J, Greig-Smith P, Pitelka F A. Decomposition in Terrestrial Ecosystems. Berkly: University of California Press, 1979: 108-109. |
| [17] | 唐仕姗, 杨万勤, 殷睿, 熊莉, 王海鹏, 王滨, 张艳, 彭艳君, 陈青松, 徐振峰. 中国森林生态系统凋落叶分解速率分解速率的分布特征及其控制因子. 植物生态学报, 2014, 38(6): 529-539. |
| [18] | Pérez-Suárez M, Arredondo-Moreno J T, Huber-Sannwald E. Early stage of single and mixed leaf-litter decomposition in semiarid forest pine-oak: the role of rainfall and microsite. Biogeochemistry, 2012, 108(1-3): 245-258. |
| [19] | Knapp A K, Fay P A, Blair J M, Collins S L, Smith M D, Carlisle J D, Harper C W, Danner B T, Lett M S, McCarron J K. Rainfall variability, carbon cycling, and plant species diversity in a mesic grassland. Science, 2002, 298(5601): 2202-2205. |
| [20] | Cardenas R E, Dangles O. Do canopy herbivores mechanically facilitate subsequent litter decomposition in soil? A pilot study from a Neotropical cloud forest. Ecological Research, 2012, 27(5): 975-981. |
| [21] | Zhou G Y, Guan L L, Wei X H, Tang X L, Liu S G, Liu J X, Zhang D Q, Yan J H. Factors influencing leaf litter decomposition: an intersite decomposition experiment across China. Plant and Soil, 2008, 311(1-2): 61-72. |
| [22] | Brandt L A, King J Y, Hobbie S E, Milchunas D G, Sinsabaugh R L. The role of photodegradation in surface litter decomposition across a grassland ecosystem precipitation gradient. Ecosystems, 2010, 13(5): 765-781. |
| [23] | Cornwell W K, Cornelissen J H C, Amatangelo K, Dorrepaal E, Eviner V T, Godoy O, Hobbie S E, Hoorens B, Kurokawa H, Pérez-Harguindeguy N, Quested H M, Santiago L S, Wardle D A, Wright I J, Aerts R, Allison S D, van Bodegom P, Brovkin V, Chatain A, Callaghan T V, Díaz S, Garnier E, Gurvich D E, Kazakou E, Klein J A, Read J, Reich P B, Soudzilovskaia N A, Vaieretti M V, Westoby M. Plant species traits are the predominant control on litter decomposition rates within biomes worldwide. Ecology Letters, 2008, 11(10): 1065-1071. |
| [24] | Hobbie S E, Reich P B, Oleksyn J, Ogdahl M, Zytkowiak R, Hale C, Karolewski P. Tree species effects on decomposition and forest floor dynamics in a common garden. Ecology, 2006, 87(9): 2288-2297. |
| [25] | 窦荣鹏, 江洪, 余树全, 马元丹, 郭培培. 柳杉凋落物在中国亚热带和热带的分解. 生态学报, 2010, 30(7): 1758-1763. |
| [26] | Berg B, Berg M P, Bottner P, Box E, Breymeyer A, Ca de Anta R, Couteaux M, Escudero A, Gallardo A, Kratz W, Madeira M, Mälkönen E, McClaugherty C, Meentemeyer V, Muñoz F, Piussi P, Remacle J, Vi de Santo A. Litter mass loss rates in pine forests of Europe and Eastern United States: some relationships with climate and litter quality. Biogeochemistry, 1993, 20(3): 127-159. |
| [27] | Heaney A, Proctor J. Chemical elements in litter in forests on Volcan Barva, Costa Rica. Mineral Nutrients in Tropical Forest and Savanna Ecosystems, Oxford: Blackwell Scientific, 1989: 225-271. |
| [28] | Godoy O, Castro-Díez P, Van Logtestijn R S P, Cornelissen J H C, Valladares F. Leaf litter traits of invasive species slow down decomposition compared to Spanish natives: a broad phylogenetic comparison. Oecologia, 2010, 162(3): 781-790. |
| [29] | Intergovernmental Panel on Climate Change. Climate Change in 2007-The Physical Science Basis. Cambridge, UK: Cambridge University Press, 2007. |