 2015, Vol. 35
2015, Vol. 35文章信息
- 陈晓丽, 王根绪, 杨燕, 杨阳
- CHEN Xiaoli, WANG Genxu, YANG Yan, YANG Yang
- 山地森林表层土壤酶活性对短期增温及凋落物分解的响应
- Response of soil surface enzyme activities to short-term warming and litter decomposition in a mountain forest
- 生态学报, 2015, 35(21): 7071-7079
- Acta Ecologica Sinica, 2015, 35(21): 7071-7079
- http://dx.doi.org/10.5846/stxb201312182982
-
文章历史
- 收稿日期:2013-12-18
- 修订日期:2015-04-14






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
凋落物作为陆地生态系统生产力的构成部分,通过分解将物质和养分返还生态系统;其分解成为全球碳收支的一个重要组成部分和流通环节[1, 2, 3]。大量研究结果表明凋落物分解主要由气候条件、凋落物性质和土壤生物群落决定[1, 2, 4, 5, 6, 7]。全球变暖作为全球气候变化主要特征之一已经改变并继续改变着全球生态系统的结构和功能[8],而对凋落物分解的影响是其中的一个重要内容。凋落物性质对凋落物分解的影响很大,强烈控制着凋落物分解和养分释放过程[9]。土壤和凋落物分解之间的作用是相互的,目前这方面的大多数研究集中在凋落物分解对增温、施碳、施氮及其交互作用的响应[10, 11, 12]和凋落物分解对土壤性质、植-土系统的影响[13, 14];而有关增温和凋落物分解对表层土壤酶活性的研究还较少。
峨眉冷杉(Abies fabri (Mast.) Craib)林是青藏高原东南缘贡嘎山东坡暗针叶林的主体,是横断山系林区的典型森林类型[15],具有调节气候、保持水土的重要生态功能,对维持区域生态安全和促进区域可持续发展具有举足轻重的作用。土壤酶是土壤生态系统的重要组成部分及土壤新陈代谢的主要参与者,在地下生态系统中的作用十分重要;推动着物质转化和能量流动[16];其活性大小表征着土壤肥力高低及养分循环速率快慢[17]。温度是影响土壤酶活性的一个关键非生物因子,全球变暖会对土壤酶活性产生复杂而深刻的作用,如Allison等[18]在阿拉斯加北方森林研究发现,生长季内增温会增加土壤铵态氮和硝态氮有效性以及活跃真菌的总体多样性;Sardans等[19]通过长期(1999—2005年)增温和干旱对地中海灌丛土壤酶活性的影响进行为期一年的研究,发现增温会不同程度地增加土壤酶活性,且主要通过对土壤温度和土壤含水量的改变来实现,而不是改变土壤有机质数量和营养状况。多数研究表明短期增温(一个或多个生长季,或一年时间)情况下,土壤温度升高会增加土壤酶活性[18, 20, 21]。更长时期增温与定位监测研究就显得十分必要。此外,土壤酸碱性也会影响土壤酶活性,每一种酶都有相应的pH值适应范围及最适宜的pH值,本研究中也测定了土壤pH值。
本研究中取峨眉冷杉林和杜鹃荚蒾(Ericaeae-Viburnum)群落不同比例混合的凋落物于林下层地表进行原位分解,采用开顶式生长箱和加热电缆(OTC-cable)联合增温的方式长期增温,监测土壤中几个重要酶活性的变化情况,探究土壤酶对增温和不同类型凋落物分解的响应,旨在为未来气候变化条件下青藏高原东南缘地区亚高山针叶林的经营管理提供数据支撑和科学依据,以期初步了解全球变暖条件下陆地生态系统的反馈作用。
1 研究区域与研究方法 1.1 研究区域概况研究区域位于贡嘎山东陂的海螺沟干河坝(N29°34′32.97″,E101°59′40.77″),海拔为3042 m,在中国科学院贡嘎山高山生态系统观测试验站(3000 m)附近。年均降雨量在1940 mm左右,年际变化不显著,但季节分布集中且不均匀。夏季降雨量大且集中;冬季气候寒冷,相对干燥。年平均空气相对湿度高达90.2%。年均蒸发量1578.6 mm。年均气温4.0 ℃,最冷月(1月)和最热月(7月)的平均气温分别为-4.3 ℃和11.9 ℃。土壤类型为粗骨土[22]。
1.2 实验方法于2009年5月,选取峨眉冷杉中龄林具有代表性的样地进行野外分解实验,样地总面积约为100 m2,地面尽量平整,坡度小于10°。取地表新鲜不同类型凋落物(A:75%峨眉冷杉针叶+25%杜鹃荚蒾灌木叶凋落物,B:55%峨眉冷杉针叶+45%杜鹃荚蒾灌木叶凋落物)加入各试验样方内,其厚度约为223 g/m。设置增温处理(T)和自然对照(CK);采取开顶式生长箱(Open top camber,OTCs)和加热电缆联合方式进行增温。其中OTC为下底面积2.6 m2,上端面积1.27 m2,高0.52 m的六棱柱体;加热电缆的功率为288 W。共4个处理组合,每种处理组合重复3次,各小样方立地条件相似。同时,凋落物失重率则采用凋落袋法(网袋规格为15 cm×20 cm,网孔直径为2 mm)测定,直接将装好凋落物的凋落袋置于林下地表;每个重复均两袋凋落物,每袋凋落物风干重为80 g,记为m0。实验时限为4a。整个实验期间,通过空气和土壤温湿度传感器对增温处理和自然对照小区中的地表空气和表层土壤的温湿度进行自动连续的观测、记录,人工定期收集数据并更换电池。
1.3 分析方法2013年5月中旬收集凋落袋,将凋落物于自然状态下风干至恒重,称重,记为m1。土壤样品分两层(0—5 cm和5—10 cm)采集,每个样方分3个小样方取土再混合成一个样品,新鲜土样过筛,于-20 ℃条件下保存。本研究测定过氧化氢酶活性、多酚氧化酶及脲酶活性,分别采用高锰酸钾滴定法、邻苯二酚法、苯酚钠-次氯酸钠比色法,酶活性单位则根据冯瑞芳等的方法表示,一个酶活性单位(EU)分别用20 min后1 g土样消耗0.1 mol/L的体积毫升数表示、1 g土样3 h内产生的红紫棓精毫克数表示、1 g土样在38 ℃下24 h内产生的NH4+-N的毫克数表示。土壤pH值的测定:取制备好的风干土样10.0 g于50 ml烧杯中,加25 ml去离子水,用玻璃棒搅拌使得充分混合,静置30 min后取上清液用pH计测得。
凋落物失重率计算公式:
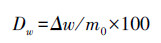
式中,Δw=m0-m1。
1.4 数据处理数据分析采用SAS8.2统计软件进行双因素方差分析(two-way ANOVA),利用Duncan检验法检验数据之间的显著性,检验增温、凋落物类型及交互作用对土壤过氧化氢酶、多酚氧化酶和脲酶活性的影响,显著水平为P < 0.05。
2 结果与分析 2.1 土壤和空气温湿度 2.1.1 土壤和空气温度的改变OTC-cable加热系统使得地表空气和表层土壤温度分别增加了2.84 ℃和 1.83 ℃ (图1),且具有统计显著性(P < 0.05)。该增温手段的增温效果明显。

|
| 图 1 自然对照和OTC-cable增温小区空气温度、土壤温度年变化 Fig.1 Annual air and soil temperature variations in the ambient plots (CK) and OTC-cable system warmed plots OTC-cble: 联合增温Open top chamber + cable warminga; CK: 自然对照 ambient control plot |
OTC-cable加热系统使得空气相对湿度和土壤含水量分别降低了5.27%和 1.55% (图2)。由图可以看出,增温和对照处理的空气相对湿度均在80%以上,即湿度仍维持在较高水平;12月至4月,增温处理的土壤含水量高于对照,这可能主要是由于研究区冬季常被厚雪,增温处理温度较高会融化一部分雪渗透入土壤中,从而使得土壤含水量升高。

|
| 图 2 自然对照和OTC-cable增温小区空气相对湿度、土壤含水量年变化 Fig.2 Annual relative air humidity (RH) and soil moisture content variations in the ambient plots (CK) and OTC-cable system warmed plots |
各处理凋落物失重率的大小为:T+B(76±3)>T+A(68±5)=CK+B(68±6)>CK+A(59±4)。即针叶比例较高凋落物失重率低于针叶比例较低凋落物失重率(P < 0.05),降幅为13%;增温处理使得不同类型凋落物的失重率均上升,均超过10%,但针叶比例较高的凋落物失重率增加幅度更大(图3)。结果表明,针叶比例高会抑制凋落物分解,增温促进凋落物分解且对针叶比例高的促进作用更加明显。

|
| 图 3 不同类型 (A和B)和气候条件(CK和T)凋落物失重率/% (平均值+标准误)P < 0.05 Fig.3 The litter weightlessness of two litter types in the ambient plots (CK) and warmed plots (T) (means +S.E.) A:75%峨眉冷杉针叶+25%杜鹃荚蒾灌木叶凋落物;B: 55%峨眉冷杉针叶+45%杜鹃荚蒾灌木叶凋落物 |
表层土壤pH值大小及相互关系如下:0—5 cm土壤: CK+B(4.91±0.10)>T+B(4.55±0.15)> T+A(4.33±0.17)>CK+A(4.18±0.07),5—10 cm土壤: CK+B(5.29±0.15)>T+B(5.12±0.17)>CK+A (4.39±0.07) >T+A (4.31±0.41) (表1和图4)。不同凋落物类型之间,凋落物针叶比例较低的土壤pH值较大,幅度在0.2—0.9之间。增温处理对土壤pH值影响并不显著;不同土层间pH值有所差异,随着土壤深度增加,土壤酸性减弱(P>0.05)。总体来看土壤pH值差异不是非常明显(P>0.05),表层土壤呈弱酸性。增温及凋落物分解不会改变土壤酸碱性以影响土壤酶活性。
| 因素 Factor | pH值 | 过氧化氢酶 Catalase | 多酚氧化酶 Polyphenol oxidase | 脲酶 Urease |
| W | 0.3842 | 0.0259 | 0.1397 | 0.3970 |
| LT | 0.0734 | 0.0044 | 0.0075 | < 0.0001 |
| W×LT | 0.2917 | 0.0017 | 0.0184 | 0.1178 |
| S | 0.0574 | 0.0002 | 0.0100 | < 0.0001 |
| W:增温 Warming; LT:凋落物类型 Litter type; S:土壤层 Soil layer | ||||

|
| 图 4 不同类型凋落物(A和B)和气候条件(CK和T)表层土壤0—5 cm和5—10 cm的pH值(平均值+标准误) Fig.4 The pH of surface soil 0—5 cm and 5—10 cm under two litter types in the ambient plots (CK) and warmed plots (T)(means + S.E.) A:75%峨眉冷杉针叶+25%杜鹃荚蒾灌木叶凋落物;B: 55%峨眉冷杉针叶+45%杜鹃荚蒾灌木叶凋落物; ns: P>0.05;* P < 0.05 |
增温对过氧化氢酶活性的效应显著(P < 0.05),而对多酚氧化酶、脲酶活性影响不明显(P>0.05);凋落物类型对3种土壤酶活性的作用明显(P < 0.05);增温与凋落物类型的交互作用对过氧化氢酶和多酚氧化酶活性的影响大(P < 0.05),而对脲酶活性未产生明显影响(P>0.05)。不同土层之间3种土壤酶活性差异明显(P < 0.05)。土壤pH值不受增温、凋落物类型及其交互作用和土壤深度的影响。增温处理和凋落物分解4a对不同土层土壤酶活性的影响方差分析显著性检验结果见表1。
2.4.1 土壤过氧化氢酶活性0—5 cm土壤过氧化氢酶活性:T+B>CK+B>CK+A>T+A。凋落物针叶比例较低的过氧化氢酶活性高于凋落物针叶比例较高的,增幅近60%;增温的效应而凋落物类型而异,降低了针叶比例高的过氧化氢酶活性34%,升高了针叶比例较低的过氧化氢酶活性约40%(P < 0.05)。5—10 cm土壤过氧化氢酶活性:CK+A>CK+B>T+B>T+A。针叶比例降低使得过氧化氢酶活性降低23%;增温降低土壤过氧化氢酶活性,幅度因凋落物类型而异,凋落物针叶比例较低的降幅大,高达66%(P < 0.05),而对针叶比例较高的凋落物无明显影响(表1和图5)。增温和凋落物类型间的交互作用明显。不同土层之间过氧化氢酶活性差异显著,大致规律:随着土壤深度加深,过氧化氢酶活性增大。

|
| 图 5 不同类型凋落物(A和B)和气候条件(CK和T)表层土壤0—5 cm和5—10 cm过氧化氢酶活性(平均值+标准误) Fig.5 The catalase activity of surface soil 0—5 cm and 5—10 cm under two litter types in the ambient plots (CK) and warmed plots (T) (means+S.E.) |
0—5 cm土壤多酚氧化酶活性:T+A>CK+A>T+B>CK+B。凋落物针叶比例较高,多酚氧化酶活性低;增温处理使得酶活性不同程度地升高,但差异均未达显著水平(P>0.05)。5—10 cm土壤多酚氧化酶活性:CK+A>T+B>CK+A>T+B。自然对照下凋落物针叶比例较高的土壤多酚氧化酶活性最高,且远高于其他3个处理;凋落物针叶比例较低降低多酚氧化酶活性多达84%(P < 0.05),而增温对土壤多酚氧化酶活性的作用因凋落物类型而表现出差异,明显降低凋落物针叶比例较高的多酚氧化酶活性近73%,增加凋落物针叶比例较低的多酚氧化酶活性近1倍(P < 0.05) (表1和图6)。随着土壤深度增加,多酚氧化酶活性大致增大(P < 0.05)。5—10 cm土壤 CK+A处理的土壤多酚氧化酶活性最大,与其他的相差一个数量级。

|
| 图 6 不同类型凋落物(A和B)和气候条件(CK和T)表层土壤0—5 cm和5—10 cm多酚氧化酶活性(平均值+标准误) Fig.6 The polyphenol oxidase(PPO) activity of surface soil 0—5 cm and 5—10 cm under two litter types in the ambient plots (CK) and warmed plots (T) (means +S.E.) |
0—5 cm土壤脲酶活性:T+B>CK+B>CK+A>T+A。自然条件下,凋落物针叶比例较低的,土壤脲酶活性高出38%;温度升高对脲酶活性的影响根据凋落物类型不同而异,降低凋落物针叶比例较高的土壤脲酶活性约30%,增大了凋落物针叶比例较低的土壤脲酶活性34%(P < 0.05)。5—10 cm土壤脲酶活性:CK+B>T+B>CK+A>T+B。凋落物针叶比例较高的,土壤脲酶活性低;温度升高会降低脲酶活性,降幅因凋落物类型而所有不同,针叶比例较低的降幅更大,为28%,差异达显著水平(表1和图7)。不同土层间土壤脲酶活性显著不同,随着土壤深度的增加,土壤脲酶活性降低(P < 0.05)。

|
| 图 7 不同类型凋落物(A和B)和气候条件(CK和T)表层土壤0—5 cm和5—10 cm脲酶活性(平均值+标准误) Fig.7 The urease activity of surface soil 0—5 cm and 5—10 cm under two litter types in the ambient plots (CK) and warmed plots (T) (means +S.E.) ns: P>0.05;* P < 0.05 |
本研究中利用OTCs-cable联合增温方式在贡嘎山东坡针叶林进行增温处理,该手段综合了OTC和加热电缆两种方式的优点,对环境的增温效果较为显著,且对其他环境因子影响较少[23, 24, 25]。本实验中土壤温度的增幅为1.83 ℃,高于Shaw和Harte[26]利用加热灯在亚高山干旱区的1.2 ℃和亚高山湿润区的0.1 ℃,Rinnan等[27]利用OTC在亚极地地区的1 ℃;接近Sjgersten和Wookey[28]利用OTC在高山树线、亚极地树线及极地树线的0.2—2 ℃上限,低于Rustad和Fernandez[23]在北方阔叶林利用加热电缆的4—5 ℃。该增温手段的增温效果达到了预期目标,和众多类似的研究相比效果显著。增温的效果除了与增温手段密切相关外,增温处理时间也有很大的影响,大多数研究均为短期增温尤其是国内的实验,大多为一个或多个生长季或1a时间增温[18, 19, 20, 21]。本实验增温时间为4a,属于中长期增温水平,增温效果明显。
徐振锋等[21]研究认为土壤温度升高利于土壤酶活性增加,将可能的机制概括为直接和间接两种途径。其中直接途径为温度通过酶动力学来影响土壤酶活性,间接途径则通过对土壤生物群落组成及其活动、植物根系和植物残体的活动、养分矿化速率的改变3种方式来影响土壤酶活性。本研究结果显示,增温对土壤酶活性的影响并不表现出一致地增加,因土壤酶的种类、凋落物类型及土壤深度而表现出差异。增温降低土壤过氧化氢酶和脲酶活性,增大多酚氧化酶活性。本研究结果中土壤过氧化氢酶活性随着土壤温度增加而降低,不同于徐振锋等[21]的结果没有明显影响,而与孙辉等[29]研究结果增温降低其活性一致;主要因为可能为土壤过氧化氢酶自身对高寒地区的低温环境较为适应,温度增加反而不利于该酶活动,进而降低其活性[30],此外增温方式和时间的差异也导致研究结果的不同。土壤脲酶活性也随着土壤温度的升高而降低,究其原因可能是增温降低了土壤含水量,在整个试验期间,土壤含水量处于较低水平,低于20%且不稳定,土壤某些微生物活动受到抑制,使得脲酶活性受影响而降低;这一结论与Steinweg等[31]的研究结果相一致。多酚氧化酶活性随土壤温度增加而增大,这一研究结果与徐振锋等[21]一致,说明增温能增加土壤多酚氧化酶活性,且不随增温时间的推移而改变。
大量研究表明,混合凋落物分解的不是简单的加合效应,不同类型凋落物之间存在着复杂的相互作用[5 ,32],具体表现为促进或抑制分解;混合分解以促进效应为主,即混合分解速率多增加,其中针阔树种混合分解的促进效应最为明显[33],也有研究表明混合分解对凋落物分解速率无显著效应[34]。本研究中凋落物分解实验采用混合凋落物,且将两个植物群落的凋落物按不同的比例进行混合,与多数凋落物分解研究采用单一种或少数几种混合凋落物有所不同。有研究显示,全球变暖使得灌木群落分布范围向高纬度或高海拔地区延伸,改变植物群落组成,从而也改变了凋落物的组成与质量[7, 10, 35]。而本研究所采用的不同类型凋落物为A:75%峨眉冷杉针叶+25%杜鹃荚蒾灌木叶凋落物和B:55%峨眉冷杉针叶+45%杜鹃荚蒾灌木叶凋落物。凋落物针叶比例升高会抑制凋落物分解,增温促进凋落物分解且对针叶比例高的促进作用更加明显。凋落物针叶比例降低,凋落物分解对3种土壤酶活性的影响大致趋势是增大。类型A接近当前气候条件下的凋落物分解状态;类型B提高了灌木凋落叶的比例,用于模拟未来全球变暖下凋落物的分解情况。自然条件下凋落物的存在几乎均为混合凋落物(人工纯林的情况较为特殊),因而本研究采用不同植物群落混合凋落物分解而得到的研究结果更接近凋落物分解的真实状态。
目前对土壤酶活性对短期增温的响应机制研究比较透彻[21, 24, 29],而关于土壤酶活性对更长时期增温的响应研究还不多。本研究针对该问题做了相应的探究。该研究仍存在一个问题,土壤酶活性具有季节动态,随着季节的不同而表现出差异,而在本试验中仅选取了一个时间点,没有做季节动态。但在时间尺度上可与其他研究进行对比,土壤酶活性对短期增温、中期增温、长期增温的响应,构成一个完整的序列。
致谢:感谢中国科学院贡嘎山高山森林生态系统观测站老师对本研究的支持。| [1] | Aerts R. Climate, leaf chemistry and leaf litter decomposition in terrestrial ecosystems: a triangular relationship. Oikos, 1997, 79(3): 439-449. |
| [2] | Robinson C H. Controls on decomposition and soil nitrogen availability at high latitudes. Plant and Soil, 2002, 242(1): 65-81. |
| [3] | Canadell J G, Le Quéré C, Raupach M R, Field C B, Buitenhuis E T, Ciais P, Conway T J, Gillett N P, Houghton R A, Marland G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. The National Academy of Sciences of the United States of America, 2007, 104(47): 18866-18870. |
| [4] | Garcia-Pausas J, Casals P, Romanyà J. Litter decomposition and faunal activity in Mediterranean forest soils: effects of N content and the moss layer. Soil Biology and Biochemistry, 2004, 36(6): 989-997. |
| [5] | Gartner T B, Cardon Z G. Decomposition dynamics in mixed species leaf litter. Oikos, 2004, 104(2): 230-246. |
| [6] | Hobbie S E, Gough L. Litter decomposition in moist acidic and non-acidic tundra with different glacial histories. Oecologia, 2004, 140(1): 113-124. |
| [7] | 徐振锋, 尹华军, 赵春章, 曹刚, 万名利, 刘庆. 陆地生态系统凋落物分解对全球气候变暖的响应. 植物生态学报, 2009, 33(6): 1208-1219. |
| [8] | Oreskes N. The scientific consensus on climate change. Science, 2004, 306(5702): 1686-1686. |
| [9] | Heal O W, Anderson J M, Swift M J. Plant litter quality and decomposition: an historical overview. Wallingford, UK: CAB International, 1997: 3-30. |
| [10] | Aerts R. The freezer defrosting: global warming and litter decomposition rates in cold biomes. Journal of Ecology, 2006, 94(4): 713-724. |
| [11] | 方华, 莫江明. 氮沉降对森林凋落物分解的影响. 生态学报, 2006, 26(9): 3127-3136. |
| [12] | 王晶苑, 张心昱, 温学发, 王绍强, 王辉民. 氮沉降对森林土壤有机质和凋落物分解的影响及其微生物学机制. 生态学报, 2013, 33(5): 1337-1346. |
| [13] | Keeler B L, Hobbie S E, Kellogg L E. Effects of long-term nitrogen addition on microbial enzyme activity in eight forested and grassland sites: implications for litter and soil organic matter decomposition. Ecosystems, 2009, 12(1): 1-15. |
| [14] | Wang C Y, Feng X G, Guo P, Han G M, Tian X J. Response of degradative enzymes to N fertilization during litter decomposition in a subtropical forest through a microcosm experiment. Ecological Research, 2010, 25(6): 1121-1128. |
| [15] | 钟祥浩, 张文敬, 罗辑. 贡嘎山地区山地生态系统与环境特征. AMBIO-人类环境杂志, 1999, 28(8): 648-654. |
| [16] | 吴秀臣, 孙辉, 杨万勤. 土壤酶活性对温度和CO2浓度升高的响应研究. 土壤, 2007, 39(3): 358-363. |
| [17] | 林先贵. 土壤微生物研究原理与方法. 北京: 高等教育出版社, 2010: 246-251. |
| [18] | Allison S D, Treseder K K. Warming and drying suppress microbial activity and carbon cycling in boreal forest soils. Global Change Biology, 2008, 14(12): 2898-2909. |
| [19] | Sardans J, Peñuelas J, Estiarte M. Changes in soil enzymes related to C and N cycle and in soil C and N content under prolonged warming and drought in a Mediterranean shrubland. Applied Soil Biology, 2008, 39(2): 223-235. |
| [20] | 冯瑞芳, 杨万勤, 张健, 邓仁菊, 简毅, 林静. 模拟大气CO2浓度和温度升高对亚高山冷杉(Abies faxoniana)林土壤酶活性的影响. 生态学报, 2007, 27(10): 4019-4026. |
| [21] | 徐振锋, 唐正, 万川, 熊沛, 曹刚, 刘庆. 模拟增温对川西亚高山两类针叶林土壤酶活性的影响. 应用生态学报, 2010, 21(11): 2727-2733. |
| [22] | 张保华, 何毓蓉, 苗国增, 丁存才. 贡嘎山东坡亚高山林区土壤结构综合评价. 山地学报, 2006, 24(4): 504-509. |
| [23] | Rustad L E, Fernandez I J. Soil warming: consequences for foliar litter deacy in a spruce-fir forest in Maine, USA. Soil Science Society of America Journal, 1998, 62(4): 1072-1080. |
| [24] | Aronson E L, McNulty S G. Appropriate experimental ecosystem warming methods by ecosystem, objective, and practically. Agricultural and Forest Meteorology, 2009, 149(11): 1791-1799. |
| [25] | Xu Z, Hu T X, Wang K Y, Zhang Y B, Xian J R. Short-term responses of phenology, leaf traits and shoot growth of four alpine shrubs in timberline ecotone to simulated global warming, Eastern Tibetan Plateau, China. Plant Species Biology, 2001, 24(1): 27-34. |
| [26] | Shaw M R, Harte J. Control of litter decomposition in a subalpine meadow-sagebrush steppe ecotone under climate change. Ecological Applications, 2001, 11: 1206-1223. |
| [27] | Rinnan R, Michelsen A, Jonasson S. Effects of litter addition and warming on soil carbon, nutrient pools and microbial communities in a subarctic heath ecosystem. Applied Soil Ecology, 2008, 39(3): 271-281. |
| [28] | Sjögersten S, Wookey P A. Decomposition of mountain birch leaf litter at the forest-tundra ecotone in the Fennoscandian mountains in relation to climate and soil conditions. Plant and Soil, 2004, 262(1/2): 215-227. |
| [29] | 孙辉, 吴秀臣, 秦纪洪, 杨万勤. 川西亚高山森林土壤过氧化氢酶活性对升高温度和CO2 浓度的响应. 土壤通报, 2007, 38(5): 891-895. |
| [30] | 谭波, 吴福忠, 杨万勤, 余胜, 刘利, 王奥, 杨玉莲. 川西亚高山/高山森林土壤氧化还原酶活性及其对季节性冻融的响应. 生态学报, 2012, 32(21): 6670-6678. |
| [31] | Steinweg J M, Dukes J S, Wallenstein M D. Modeling the effects of temperature and moisture on soil enzyme activity: Linking laboratory assays to continuous field data. Soil Biology & Biochemistry, 2012, 55: 85-92. |
| [32] | Hättenschwiler S, Tiunov A V, Scheu S. Biodiversity and Litter Decomposition in Terrestrial Ecosystems. Annual Review of Ecology, Evolution, and Systematics, 2005, 36(1): 191-218. |
| [33] | 许晓静, 张凯, 刘波, 兰长春, 徐小牛. 森林凋落物分解研究进展. 中国水土保持科学, 2007, 5(4): 108-114. |
| [34] | 陈瑾, 李扬, 黄建辉. 内蒙古典型草原4种优势植物凋落物的混合分解研究. 植物生态学报, 2011, 35(1): 9-16. |
| [35] | Hollister R D, Webber P J, Tweedie C E. The response of Alaskan arctic tundra to experimental warming: differences between short-and long-term responses. Global Change Biology, 2005, 11(4): 525-536. |