 2015, Vol. 35
2015, Vol. 35文章信息
- 麻秋云, 韩东燕, 刘贺, 薛莹, 纪毓鹏, 任一平
- MA Qiuyun, HAN Dongyan, LIU He, XUE Ying, JI Yupeng, REN Yiping
- 应用稳定同位素技术构建胶州湾食物网的连续营养谱
- Construction of a continuous trophic spectrum for the food web in jiaozhou bay using stable isotope analyses
- 生态学报, 2015, 35(21): 7207-7218
- Acta Ecologica Sinica, 2015, 35(21): 7207-7218
- http://dx.doi.org/10.5846/stxb201310312632
-
文章历史
- 收稿日期: 2013-10-31
- 修订日期: 2015-04-14






营养级是海洋食物网研究的重要内容之一。大量研究表明,食物网中氮稳定同位素比值会随着营养层次的升高出现稳定富集的现象,因此生物在食物网中的营养层次可以用氮稳定同位素比值来表征[1, 2, 3]。国内外学者应用稳定同位素技术对水域生态系统食物网开展了一系列相关的研究。例如:Vizzini等应用碳氮稳定同位素技术研究了地中海保护区鱼类的摄食和营养级[4];Kaehler等基于碳氮稳定同位素分析,研究了Prince Edward岛海洋食物网的营养结构[5];蔡德陵等运用稳定同位素先后研究了崂山湾[6, 7]、渤海[8]和黄东海[9]生态系统的食物网结构;全为民对长江口盐沼湿地食物网进行了稳定同位素的初步研究[10];徐军应用碳、氮稳定同位素探讨了巢湖等4个淡水湖泊的食物网结构和营养级关系[11]。稳定同位素技术已经成为研究水域生态系统食物网结构、营养关系及其动态变化的一种重要方法[12]。
胶州湾位于黄海之滨,是一个典型的半封闭型海湾,湾内水域生产力高,饵料生物丰富,是多种经济鱼、虾、蟹类繁殖、育幼和索饵的场所[13]。在20世纪80年代,杨伟祥等使用传统的胃含物分析方法对胶州湾食物网进行了研究[14],但尚未见应用稳定同位素技术对胶州湾食物网进行研究的报道。本文应用稳定同位素技术对胶州湾食物网进行研究,探讨主要生物种类的碳氮稳定同位素特征,进而分析其营养级并构建胶州湾食物网的连续营养谱,旨在为深入研究胶州湾生物群落的营养结构以及食物网的物质循环和能量流动提供基础资料。
1 材料与方法 1.1 样品采集样品采自2011年春季(5月)和秋季(11月)在胶州湾海域(35°59′—36°07′N,120°13′—120°23′E)进行的底拖网调查。本次调查采用国际上通用的分层随机取样(stratified random sampling)的方法设置调查站位[15, 16, 17],每个航次分别在胶州湾的湾内、湾口和湾外随机设置5个、3个和4个调查站位(每1′×1′的方格内设置一个站位),共12个调查站位(图1)。调查船为44 kW左右的单拖渔船,调查网具网口目数为900目,囊网网目10 mm,平均拖速2 kn,每站拖网时间0.5 h左右。所有渔获样品冷冻保存。在进行拖网调查的同时,使用0.05m2箱式采泥器采集底栖生物,泥样经0.5mm套筛冲洗,滤出样品冷冻保存。在每个站位分别用浅水Ⅰ型和Ⅲ型浮游生物网采集浮游动物和浮游植物样品,都是自水底至水表垂直拖网采样,样品冷藏带回实验室立即处理。

|
| 图 1 2011年春季和秋季胶州湾底拖网调查站位 Fig.1 Sampling stations by bottom trawl survey in Jiaozhou Bay in spring and autumn 2011 |
在实验室中,对渔获物进行种类鉴定,并按照《海洋调查规范》[18]进行生物学测定,包括体长(mm)和体重(g)等生物学参数。根据每个站位不同种类的体长组成,选取不同个体大小的样品以备分析,将采自湾内、湾口和湾外的样品分别合并为一个样品进行同位素分析。对于3种优势鱼类——普氏栉虾虎鱼Acentrogobius pflaumii、六丝钝尾虾虎鱼Amblychaeturichthys hexanema和方氏云鳚Pholis fangi,则分别以5mm或10mm体长为间隔进行取样,每个体长组选取1条鱼留样进行同位素分析。其中鱼类和无脊椎动物取肌肉组织作为分析样品,浮游植物和浮游动物取生物个体的全体。
对于浮游植物样品,先用180 μm 筛绢网过滤掉浮游动物,再用35 μm 生物网采集浮游植物,加蒸馏水于塑料瓶中静置;取上清液用燃烧过的玻璃纤维滤膜(GF/C)(450℃下灼烧6 h)过滤。对于浮游动物样品,首先在冰箱冷藏柜4℃保存24 h,以去除胃中的残留食物[19],用浮游动物网过滤后,在解剖镜下挑除杂质,然后使用不同粒径的筛绢网对浮游动物进行分级(>900 μm,500—900 μm,300—500 μm和100—300 μm)。在进行下一步分析前,所有样品于-20℃的冰柜中保存。
将样品放于烘箱中(DHG-9070A)60℃下烘干至恒重,使用玻璃研钵充分研磨,然后用1 mol/L盐酸酸化以去除碳酸盐的影响[20],并用脱脂溶液(甲醇∶氯仿∶水= 2∶1∶0.8)进行脱脂处理[21]。干燥后再次研磨,放入玻璃瓶中,干燥保存。
1.3 同位素分析实验样品的稳定同位素分析在中国科学院海洋研究所进行,运用英国GV公司的IsoPrime稳定同位素质谱仪测定样品的碳氮稳定同位素比值。
碳、氮稳定同位素比值用国际通用的δ值表示,分别以VPDB (Vienna Pee Dee Belemnite)国际标准和大气氮作为参考标准。δ13C、δ15N分别按以下公式算出:
式中,13C/12CVPDB为国际标准物VPDB的碳同位素比值,15N/14Nair 为标准大气氮同位素比值。为保持实验结果的准确性和仪器的稳定性,每测试5个样品后加测1个标准样,个别样品则进行2—3次复测。δ13C值的分析精度为±0.15‰,δ15N值的分析精度为±0.18‰。 1.4 聚类分析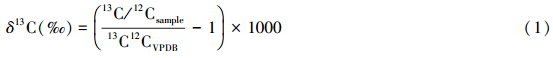

根据各种类δ13C值与δ15N值的标准化欧氏距离(Normalized Euclidean distance),采用多元统计分析软件PRIMER v5中的等级聚类分析(hierarchical cluster analysis)划分胶州湾食物网主要生物种类的营养组群。
1.5 营养级计算生物种类营养级(Trophic Level)的计算公式[2]如下:
式中,δ15Nsample 表示渔获生物氮稳定同位素比值;δ15Nbaseline 表示基准生物氮稳定同位素平均比值,本研究取中型浮游动物(即样品中粒径为300 μm—900 μm的浮游动物)[22]的平均值7.2‰;Δδ15N表示一个营养级的氮富集度,平均值为3.4‰[1];2为基准生物(初级消费者)的营养级。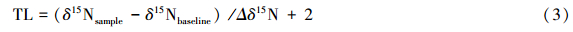
本研究选择海洋食物网营养层次1—5级的划分标准[23],并对引文中使用营养层次0—4级的划分标准进行修正,即每种生物的营养级加1。
2 结果 2.1 胶州湾主要生物种类的同位素特征在本次调查中,除浮游植物和4种粒径范围的浮游动物外,共分析了63个生物种类的稳定同位素值,其中鱼类最多(34种),其次是虾类13种,蟹类和头足类各有5种,其它生物种类6种。
胶州湾食物网δ13C值范围为-25.63‰—-17.16‰,总跨度为8.47‰,平均值为(-19.42±1.80)‰;δ15N值的范围为4.15‰—14.11‰,总跨度为9.96‰,略大于δ13C,平均值为(11.98±1.77)‰。
由图2可以看出,胶州湾各种生物种类的碳、氮稳定同位素比值差异较大。其中,鱼类碳氮稳定同位素比值的范围较大:δ13C值最小值为(-22.49±0.61)‰,最大值为(-17.26±2.54)‰,平均值为-19.29‰,跨度为5.23‰;而δ15N值的范围是9.68‰—14.11‰,平均值为12.60‰,跨度为4.43‰。

|
| 图 2 胶州湾食物网主要生物种类的碳、氮稳定同位素比值(平均值±标准差) Fig.2 δ13C and δ15N values (average ± SD) of major species in the food web of Jiaozhou Bay |
头足类δ13C值的范围是-19.16‰—-17.81‰,平均值为-18.60‰;而δ15N值的范围是12.05‰—13.55‰,平均值为12.87‰。虾类的δ13C值最小为-20.17‰,最大为(-17.75±1.12)‰,差值为2.42‰;而δ15N值最小值为(10.68±0.57)‰,最大为13.76‰,相差3.08‰。虾类的δ13C平均值为-18.96‰,δ15N平均值为12.03‰。蟹类δ13C值的范围是-19.50‰—-17.16‰,平均值为-18.47‰;而δ15N值的范围是11.14‰—13.30‰,平均值为11.98‰。
浮游生物的碳、氮稳定同位素比值是所有生物种类中最小的。其中大于 900 μm浮游动物的δ13C和δ15N值都是浮游生物的最高值,浮游植物的δ15N值最低(4.15‰),而300—500 μm浮游动物的δ13C值最小(-25.63‰)。浮游动物δ13C平均值为(-24.74±0.89)‰,跨度为1.67‰;δ15N平均值为(7.49±1.00)‰,跨度略大,为2.15‰。
2.2 聚类分析基于δ13C和δ15N值的欧几里得距离对胶州湾食物网的主要生物种类进行分类,即将具有相近最终营养来源和营养级的生物种类划为一类。根据聚类分析结果,在1.5的欧氏距离上可将胶州湾食物网的主要生物种类划分为4个营养组群:第一组群为浮游植物,属于初级生产者;第二组群为浮游动物,属于食物网中的初级消费者;第三组群包括经氏壳蛞蝓(Philine kinglippini)、玉筋鱼(Ammodytes personatus)和尖海龙(Syngnathus acus),属于食物网中的次级消费者;第四组群则为其它无脊椎动物和鱼类(图3)。

|
| 图 3 胶州湾食物网主要生物种类碳氮稳定同位素比值的聚类分析 Fig.3 Cluster analyses based on Normalized Euclidean distances of δ13C and δ15N for species in the food web of Jizozhou Bay |
经计算,胶州湾食物网主要生物种类的营养级范围为1.10(浮游植物)—4.03(红狼牙虾虎鱼Odontamblyopus rubicundus),营养层次长度为4级(表1)。其中,鱼类营养级的范围为2.73至4.03。有两种鱼类(尖海龙和玉筋鱼)的营养级低于3;营养级高于4的只有红狼牙虾虎鱼;大部分鱼类(82%)集中在3.40—3.90的营养级范围内(表1)。
| 种类 Species | n | SL/(mm)Range | TL NAverage ± SD | TL D | ΔTL |
| 浮游植物 Phytoplankton | 1 | 1.10 | |||
| 500—900μm浮游动物 Zooplankton | 3 | 1.83 ± 0.09 | |||
| 100—300μm浮游动物 Zooplankton | 3 | 1.88 ± 0.46 | |||
| 300—500μm浮游动物 Zooplankton | 3 | 2.18 ± 0.28 | |||
| > 900μm浮游动物 Zooplankton | 3 | 2.46 ± 0.27 | |||
| 尖海龙 Syngnathus acus | 2 | 135—202 | 2.73 ± 0.01 | ||
| 经氏壳蛞蝓 Philine kinglippini | 2 | 2.96 ± 0.03 | |||
| 玉筋鱼 Ammodytes personatus | 1 | 102—140 | 2.99 | 3.3[24] | -0.31 |
| 栉江珧 Atrina pectinata | 1 | 3.02 | |||
| 海蜇虾 Latreutes anoplonyx | 3 | 12—25 | 3.02 ± 0.17 | ||
| 葛氏长臂虾 Palaemon gravieri | 1 | 56 | 3.08 | 3.32[25] | -0.24 |
| 双斑蟳 Charybdis bimaculata | 3 | 7—30 *** | 3.16 ± 0.27 | 3.37[25] | -0.21 |
| 多毛类 Polychaeta | 1 | 3.19 | |||
| 疣背宽额虾 Latreutes planirostris | 3 | 11—34 | 3.19 ± 0.12 | ||
| 中颌棱鯷 Thryssa mystax | 1 | 83—87 | 3.23 | 3.4[26] | -0.17 |
| 鹰爪虾 Trachysalambria curvirostris | 1 | 55—63 | 3.27 | 3.33[25] | -0.06 |
| 绿鳍鱼 Chelidonichthys kumu | 1 | 168—172 | 3.28 | 3.65[27] | -0.37 |
| 戴氏赤虾 Metapenaeopsis dalei | 2 | 25—52 | 3.31 ± 0.27 | 3.37[25] | -0.06 |
| 日本鼓虾 Alpheus japonicus | 3 | 25—55 | 3.33 ± 0.03 | 3.5[28] | -0.17 |
| 栉孔扇贝 Chlamys farreri | 1 | 3.39 | |||
| 石鲽 Kareius bicoloratus | 1 | 140—160 | 3.40 | 4.1[24] | -0.7 |
| 鲬 Platycephalus indicus | 1 | 170—240 | 3.40 | 4[24] | -0.6 |
| 日本蟳 Charybdis japonica | 1 | 12—50 *** | 3.41 | 3.9[28] | -0.49 |
| 纹缟虾虎鱼 Tridentiger trigonocephalus | 1 | 37—66 | 3.43 | 3.1[26] | 0.33 |
| 双喙耳乌贼 Sepiola birostrata | 3 | 9—22 ** | 3.43 ± 0.28 | 3.7[28] | -0.27 |
| 褐菖鲉 Sebastiscus marmoratus | 1 | 42—44 | 3.45 | 3.6[26] | -0.15 |
| 细巧仿对虾 Parapenaeopsis tenella | 3 | 28—62 | 3.48 ± 0.15 | ||
| 鲜明鼓虾 Alpheus distinguendus | 1 | 29—60 | 3.49 | 3.3[28] | 0.19 |
| 长丝虾虎鱼 Myersina filifer | 2 | 58—81 | 3.49 ± 0.06 | ||
| 长足七腕虾 Heptacarpus futilirostris | 1 | 11—35 | 3.50 | ||
| 短鳍 Callionymus sagitta | 1 | 48—107 | 3.51 | 3.4[29] | 0.11 |
| 皮氏叫姑鱼 Johnius belengerii | 2 | 70—98 | 3.51 ± 0.19 | 3.73[27] | -0.22 |
| 细纹狮子鱼 Liparis tanakae | 2 | 45—130 | 3.52 ± 0.13 | 4.3[24],3.9[27] | -0.78,-0.38 |
| 寄居蟹 Pagurus sp. | 1 | 3.53 | |||
| 许氏平鲉 Sebastes schlegelii | 3 | 76—117 | 3.53 ± 0.02 | 4.7[24] | -1.17 |
| 铠平鲉 Sebastes hubbsi | 2 | 46—105 | 3.53 ± 0.27 | ||
| 钟馗虾虎鱼 Tridentiger barbatus | 1 | 43—88 | 3.54 | ||
| 普氏栉虾虎鱼 Acentrogobius pflaumii | 49 | 25—52 | 3.55 ± 0.34 | ||
| 方氏云鳚 Pholis fangi | 47 | 99—155 | 3.55 ± 0.53 | 4.6[24] | -1.05 |
| 中华安乐虾 Eualus sinensis | 1 | 20—26 | 3.58 | ||
| 多鳞鱚 Sillago sihama | 2 | 67—120 | 3.58 ± 0.19 | 3.4[24] | 0.18 |
| 繸鳚 Chirolophis japonicus | 1 | 102—194 | 3.60 | ||
| 细螯虾 Leptochela gracilis | 1 | 24 | 3.61 | ||
| 日本关公蟹 Dorippe japonica | 1 | 20 *** | 3.63 | ||
| 白姑鱼 Pennahia argentata | 3 | 62—108 | 3.63 ± 0.46 | 3.9[24] | -0.27 |
| 短蛸 Octopus ocellatus | 2 | 47—55 ** | 3.64 ± 0.05 | ||
| 李氏 Callionymus richardsonii | 1 | 70—91 | 3.66 | 3.3[30] | 0.36 |
| 六丝钝尾虾虎鱼 Amblychaeturichthys hexanema | 50 | 42—128 | 3.66 ± 0.36 | 3.3[24] | 0.36 |
| 脊腹褐虾 Crangon affinis | 3 | 20—71 | 3.67 ± 0.10 | 3.3[25] | 0.37 |
| 长蛇鲻 Saurida elongata | 2 | 154—195 | 3.67 ± 0.16 | 4.3[27] | -0.63 |
| 细条天竺鲷 Apogon lineatus | 1 | 48—49 | 3.68 | 3.43[27] | 0.25 |
| 矛尾虾虎鱼 Chaeturichthys stigmatias | 3 | 61—157 | 3.68 ± 0.36 | 4[24],3.7[27] | -0.32,-0.02 |
| 蓝无壳侧鳃海牛 Pleurobranchaea novaezealandiae | 2 | 3.70 ± 0.02 | |||
| 四盘耳乌贼 Euprymna morsei | 1 | 22 ** | 3.72 | ||
| 斑尾刺虾虎鱼 Synechogobius ommaturus | 2 | 141—293 | 3.76 ± 0.53 | 4[24] | -0.24 |
| 口虾蛄 Oratosquilla oratoria | 1 | 48—203 | 3.77 | 3.38[25] | 0.39 |
| 长蛸 Octopus variabilis | 3 | 58—158 ** | 3.78 ± 0.04 | ||
| 三疣梭子蟹 Portunus trituberculatus | 1 | 48—68 *** | 3.79 | 3.37[25] | 0.42 |
| 星康吉鳗 Conger myriaster | 3 | 212—352 * | 3.81 ± 0.40 | 4.8[24] | -0.99 |
| 大泷六线鱼 Hexagrammos otakii | 3 | 55—228 | 3.86 ± 0.14 | 4.2[24] | -0.34 |
| 黄鮟鱇 Lophius litulon | 1 | 270 | 3.87 | 4.5[24],4.4[27] | -0.63,-0.53 |
| 小头栉孔虾虎鱼 Paratrypauchen microcephalus | 1 | 65—69 | 3.87 | ||
| 枪乌贼 Loliolus sp. | 2 | 25—89 ** | 3.87 ± 0.14 | 3.65[25] | 0.22 |
| 短吻红舌鳎 Cynoglossus joyneri | 1 | 117—127 * | 3.89 | 3.9[24] | -0.01 |
| 小黄鱼 Larimichthys polyactis | 2 | 116—148 | 3.90 ± 0.12 | 3.57[27] | 0.33 |
| 凡纳滨对虾 Litopenaeus vannamei | 1 | 133—155 | 3.93 | ||
| 长吻红舌鳎 Cynoglossus lighti | 1 | 71—168 * | 3.97 | ||
| 红狼牙虾虎鱼 Odontamblyopus rubicundus | 1 | 142 | 4.03 | ||
| ΔTL=TLW-TLD; *全长;** 胴长; ** * 头胸甲长;n 样品数;SL 体长;TLN通过氮稳定同位素比值所得营养级;TLD 文献中胃含物分析所得营养级;ΔTL 营养级差值;文献[24]、[25]和[30]中的营养级已修正 | |||||
头足类和虾蟹类均属于第Ⅲ营养级(3—4)。在头足类中,营养级最高的为枪乌贼Loliolus sp.(3.87±0.10),其次为长蛸Octopus variabilis(3.78),双喙耳乌贼Sepiola birostrata的营养级最低,为3.43±0.16。在种类较多的虾类中,营养级最高值为凡纳滨对虾Litopenaeus vannamei的3.93,最低值为海蜇虾Latreutes anoplonyx的3.02。在蟹类中,营养级最高的是三疣梭子蟹(Portunus trituberculatus)(3.79),双斑蟳Charybdis bimaculata的营养级最低,为3.16。另外,经氏壳蛞蝓的营养级为2.96,是除浮游动物外无脊椎动物中最小的营养级(表1)。
在浮游生物中,浮游植物的营养级为1.10,作为初级生产者,是胶州湾食物网中营养级最低的种类。浮游动物的营养级范围是1.83—2.46,属于初级消费者(表1)。
2.4 营养谱根据营养级的计算结果,构建了胶州湾食物网的连续营养谱(图4)。位于1.0—1.5营养级上的生物种类只有浮游植物;位于1.5—2.0营养级上的生物种类有两类,分别为粒径100—300μm和500—900μm的浮游动物;位于2.0—2.5营养级上的生物种类也有两类,分别为粒径300—500μm和>900μm的浮游动物;位于2.5—3.0营养级上的生物种类有3种,即尖海龙、玉筋鱼和经氏壳蛞蝓;位于3.0—3.5营养级上的生物种类有21种,包括7种鱼类、8种虾类、2种蟹类、1种头足类、2种贝类和沙蚕;位于3.5—4.0营养级上的生物种类有38种,包括24种鱼类、5种虾类、3种蟹类、4种头足类以及口虾蛄Oratosquilla oratoria和蓝无壳侧鳃海牛Pleurobranchaea novaezealandiae;营养级高于4.0的种类只有红狼牙虾虎鱼一种。

|
| 图 4 胶州湾食物网的连续营养谱 Fig.4 The continuous trophic spectrum for the food web of Jiaozhou Bay |
本次胶州湾春、秋季节调查,用于同位素分析生物种类的生物量之和占总生物量的95%,基本涵盖了胶州湾周年出现的主要生物种类[31, 32, 33, 34],因此本研究结果可以反映胶州湾食物网的基本情况。
研究发现,胶州湾食物网中各生物类别δ13C和δ15N的数值范围存在较大差异,其中鱼类种类数最多(34种),其δ13C和δ15N跨度都是最大的,分别为5.23‰和4.43‰。鱼类较大的δ13C和δ15N跨度不仅与种类数较多有关,也与个别种有关,即尖海龙和玉筋鱼,其δ13C和δ15N值均偏低。除此之外的32种鱼类,其δ13C值主要集中在-20.57‰—-17.26‰,跨度为3.31‰;δ15N值主要集中在11.39‰—14.11‰,跨度为2.72‰。
本研究中,共有17个生物种类与2005年蔡德陵在东黄海进行的同位素研究[9]相同。通过比较发现,这17个相同种类在本研究中的δ15N值均较大,差值为0.33‰—3.35‰,其中小黄鱼Larimichthys polyactis差值最大,其余16个种类的差值均小于3.00‰。而与2010年李忠义对南黄海渔获物进行的同位素研究[35]相比,胶州湾11种相同种类的δ15N值依然较大,只有双斑蟳的略小(差值为0.38‰),其中差值较大的有细螯虾Leptochela gracilis(7.00‰)、大泷六线鱼Hexagrammos otakii(5.73‰)和双喙耳乌贼(4.07‰),其余种类的差值均小于3.00‰。造成这种差别原因可能是多方面的。首先,由于研究海域和调查时间不同,捕食者的基础饵料生物不同,从而造成了同种生物δ15N值的差异。另一方面可能与体长差异有关,例如:本研究中大泷六线鱼体长范围55—225mm,δ15N值为(13.54±0.47)‰,而南黄海大泷六线鱼的体长范围为46—55mm,相应的δ15N值也较小,为7.81‰[35]。
胶州湾春季浮游动物δ15N和δ13C的数值范围分别是6.61‰—8.76‰和-25.63‰—-23.96‰,跨度分别为2.15‰和1.67‰。由于在分析时将浮游动物按照粒径进行区分,造成不同食性浮游动物的分离,因此浮游动物样品氮同位素的数值范围较大,营养级跨度也较大。浮游动物的粒径越大,其营养级就越高,δ15N值随营养层次升高会出现稳定富集现象[1],而δ13C值富集度极小[36]。因此浮游动物的δ15N值整体上随着粒径的增大而逐渐增大。本研究结果显示,胶州湾春季大型浮游动物优势种主要为中华哲水蚤Calanus sinicus、夜光虫Noctiluca scientillans和强壮箭虫Sagitta crassa等,均为大于500μm的浮游动物,δ15N和δ13C平均值分别为(7.69±1.33)‰和(-23.97±0.95)‰,均与蔡德陵的研究结果相近[8]。
胶州湾2011年春季浮游植物δ15N值为4.15‰,δ13C平均值为-25.03 ‰± 0.51‰,都比南黄海北部春季浮游植物的δ15N、δ13C值[9]要小,两地浮游植物优势种类的不同可能是造成δ15N、δ13C差异的主要原因[37, 38]。
3.2 营养级的比较分析本研究按照海洋食物网营养层次1—5级的划分标准,计算得出胶州湾食物网主要生物种类的营养层次为4级:浮游植物为1.10级,高级肉食性鱼类红狼牙虾虎鱼为4.03级。同样基于氮稳定同位素计算生物营养级,本次调查的胶州湾食物网的营养层次范围明显大于黄东海(1.43—3.62)[9],这主要是因为本研究的生物种类包括了位于食物网底层的初级生产者(浮游植物)和初级消费者(浮游动物),因此营养级的跨度较大。
Vander Zanden等[39]应用稳定同位素技术和胃含物分析的方法计算了Ontario和Quebec 36个湖泊8种鱼类的营养级,发现两种方法所计算的结果相似。本研究检索了我国近年来的相关研究,在涉及胶州湾食物网各个种类的胃含物分析营养级报道中,有文献可查的种类有37种,包括25种鱼类、7种虾类、3种蟹类和2种头足类(表1)。通过比较发现,两者分析的营养级结果具有很好的一致性,约78%的种类(29种)在0.5个营养级的误差范围之内是一致的,这与国内学者蔡德陵[9]、李忠义[35]和万炜[40]的研究结果相似。因此可以认为,氮稳定同位素法是一种研究水生生态系统食物网营养位置的有效方法。
在本研究中,有8种鱼类δ15N分析的营养级明显低于1992年韦晟运用胃含物分析的黄海同种鱼类的营养级[24]。差异最大的是许氏平鲉Sebastes schlegelii(差值为1.17);其次是方氏云鳚,差值为1.05;其余6种鱼类(星康吉鳗Conger myriaster、细纹狮子鱼Liparis tanakae、石鲽Kareius bicoloratus、长蛇鲻Saurida elongata、黄鮟鱇Lophius litulon和鲬Platycephalus indicus)差值在0.60至0.99之间。
与稳定同位素分析相比,胃含物分析只能反映鱼类短时间内的摄食情况,且分析时容易忽略一些小型、易消化的饵料种类,因此可能造成营养级的计算结果较实际值偏高[4]。此外,这些鱼类营养级降低还与其食物组成中饵料生物种类的变化有关。通过分析发现,这些鱼类的主要饵料生物普遍由小型鱼类等高营养级的饵料生物转换为营养级较低的饵料生物,饵料生物营养级的下降可能是导致捕食鱼类营养级降低的主要原因[41],这种现象在其它海域也有过报道[41, 42, 43, 44]。另一方面,鱼类个体的体长组成也会对营养级的计算结果造成影响,例如:本研究中采集的许氏平鲉体长范围是76—117mm,细纹狮子鱼体长范围是45—130mm,个体均较小,因此导致其营养级偏低。
3.3 胶州湾食物网连续营养谱的特征根据营养级计算的结果,本研究构建了胶州湾食物网的连续营养谱。胶州湾34种鱼类的营养级范围为2.73—4.03,主要集中于3.0—4.0之间。只有尖海龙和玉筋鱼的营养级低于3.0,同时也只有红狼牙虾虎鱼的营养级高于4.0,为4.03。在营养谱中,鱼类主要属于中级和顶级消费者。20世纪80年代杨伟祥对胶州湾53种鱼类的胃含物分析表明,66%(35种)的种类以浮游动物和底栖生物为食,属于3.0— 4.0的第Ⅲ营养级[14],本文的研究结果与之相似。在胶州湾食物网中,浮游植物和浮游动物位于营养谱的底层,而其它无脊椎动物主要位于第Ⅲ营养级,营养级范围为2.96—3.93,29个种类中只有经氏壳蛞蝓的营养级低于3.0。
本研究中34种鱼类,营养级范围为2.73—4.03,跨度为1.30。这是由于近年来胶州湾鱼类趋向小型化、低龄化[31, 34],本次调查所采集的鱼类主要为杂食性鱼类,而高级肉食性鱼类,如星康吉鳗又由于个体较小的原因,其营养级仅为3.8±0.4,一些历史资料中记录的胶州湾植食性和碎屑食性的鱼类(营养级2.0—2.9)[14]在本次调查中均未采集到,如鮻鱼Liza haematocheilus和鲻鱼Mugil cephalus等。这也从另一方面说明了胶州湾鱼类群落结构的衰退。
致谢:中国科学院海洋研究所于心科研究员和南青云老师对样品测定给予帮助,中国海洋大学水产学院刘群教授给予支持,特此致谢。
| [1] | Minagawa M, Wada E. Stepwise enrichment of 15N along food chains: Further evidence and the relation between δ15N and animal age. Geochimica et Cosmochimica Acta, 1984, 48(5): 1135-1140. |
| [2] | Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology, 2002, 83(3): 703-718. |
| [3] | Deniro M J, Epstein S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta, 1981, 45(3): 341-351. |
| [4] | Vizzini S, Mazzola A. Stable isotopes and trophic positions of littoral fishes from a Mediterranean marine protected area. Environmental Biology of Fishes, 2009, 84(1): 13-25. |
| [5] | Kaehler S, Pakhomov E A, McQuaid C D. Trophic structure of the marine food web at the Prince Edward Islands (Southern Ocean) determined by δ13C and δ15N analysis. Marine Ecology Progress Series, 2000, 208: 13-20. |
| [6] | 蔡德陵, 孟凡, 韩贻兵, 高素兰. 13C/12C 比值作为海洋生态系统食物网示踪剂的研究--崂山湾水体生物食物网的营养关系. 海洋与湖沼, 1999, 30(6): 671-678. |
| [7] | 蔡德陵, 洪旭光, 毛兴华, 张淑芳, 韩贻兵, 高素兰. 崂山湾潮间带食物网结构的碳稳定同位素初步研究. 海洋学报, 2001, 23(4): 41-47. |
| [8] | Cai D L, Wang R, Bi H S. Trophic relationships in the Bohai ecosystem: preliminary investigation from δ13C analysis. Acta Ecologica Sinica, 2001, 21(8): 1354-1359. |
| [9] | 蔡德陵, 李红燕, 唐启升, 孙耀. 黄东海生态系统食物网连续营养谱的建立: 来自碳氮稳定同位素方法的结果. 中国科学C辑: 生命科学, 2005, 35(2): 123-130. |
| [10] | 全为民. 长江口盐沼湿地食物网的初步研究: 稳定同位素分析[D]. 上海: 复旦大学, 2007. |
| [11] | 徐军. 应用碳、氮稳定性同位素探讨淡水湖泊的食物网结构和营养级关系[D]. 武汉: 中国科学院研究生院(水生生物研究所), 2005. |
| [12] | Michener R H, Kaufman L. Stable isotope ratios as tracers in marine aquatic food webs: An update // Lajtha K, Michener R H, eds. Stable Isotopes in Ecology and Environmental Science. Oxford: Blackwell, 2007: 238-270. |
| [13] | 刘瑞玉. 胶州湾自然环境特点 // 刘瑞玉. 胶州湾生态学和生物资源. 北京: 科学出版社, 1992: 2-3. |
| [14] | 杨伟祥, 杨纪明, 吴鹤洲. 鱼类资源--饵料生物和鱼类摄食习性 // 刘瑞玉. 胶州湾生态学和生物资源. 北京: 科学出版社, 1992: 413-423. |
| [15] | Cochran W G. Sampling Techniques. New York: John Wiley and Sons, 1977: 900. |
| [16] | Miller T J, Skalski J R, Ianelli J N. Optimizing a stratified sampling design when faced with multiple objectives. ICES Journal of Marine Science, 2007, 64(1): 97-109. |
| [17] | Manly B F, Akroyd J A M, Walshe K A. Two-phase stratified random surveys on multiple populations at multiple locations. New Zealand Journal of Marine and Freshwater Research, 2002, 36(3): 581-591. |
| [18] | 中华人民共和国国家质量监督检验检疫总局, 中国国家标准化管理委员会. GB/T 12763. 6-2007海洋调查规范第6部分: 海洋生物调查. 北京: 中国标准出版社, 2007. |
| [19] | Junger M, Planas D. Quantitative use of stable carbon isotope analysis to determine the trophic base of invertebrate communities in a boreal forest lotic system. Canadian Journal of Fisheries and Aquatic Sciences, 1994, 51(1): 52-61. |
| [20] | Jacob U, Mintenbeck K, Brey T, Knust R, Beyer K. Stable isotope food web studies: a case for standardized sample treatment. Marine Ecology Progress Series, 2005, 287: 251-253. |
| [21] | Bligh E G, Dyer W J. A rapid method of total lipid extraction and purification. Canadian Journal of Biochemistry and Physiology, 1959, 37(8): 911-917. |
| [22] | Tamelander T, Renaud P E, Hop H, Carroll M L, Ambrose W G, Hobson K A. Trophic relationships and pelagic-benthic coupling during summer in the Barents Sea Marginal Ice Zone, revealed by stable carbon and nitrogen isotope measurements. Marine Ecology Progress Series, 2006, 310: 33-46. |
| [23] | Jiming Y. A Tentative Analysis of the Trophic Levels of North Sea Fish. Marine Ecology Progress Series, 1982, 7: 247-252. |
| [24] | 韦晟, 姜卫民. 黄海鱼类食物网的研究. 海洋与湖沼, 1992, 23(2): 182-192. |
| [25] | 程济生, 朱金声. 黄海主要经济无脊椎动物摄食特征及其营养层次的研究. 海洋学报, 1997, 19(6): 102-108. |
| [26] | 黄良敏, 张雅芝, 潘佳佳, 崔裕侠, 吴亚娟. 厦门东海域鱼类食物网研究. 台湾海峡, 2008, 27(1): 64-73. |
| [27] | 薛莹. 黄海中南部主要鱼种摄食生态和鱼类食物网研究[D]. 青岛: 中国海洋大学, 2005. |
| [28] | 杨纪明. 渤海无脊椎动物的食性和营养级研究. 现代渔业信息, 2001, 16(9): 8-16. |
| [29] | 杨纪明. 渤海鱼类的食性和营养级研究. 现代渔业信息, 2001, 16(10): 10-19. |
| [30] | 张雅芝, 李福振, 刘向阳, 蓝晞文, 陈锦坤. 东山湾鱼类食物网研究. 台湾海峡, 1994, 13(1): 52-61. |
| [31] | 曾晓起, 朴成华, 姜伟, 刘群. 胶州湾及其邻近水域渔业生物多样性的调查研究. 中国海洋大学学报, 2004, 34(6): 977-982. |
| [32] | 陈晓娟, 薛莹, 徐宾铎, 任一平, 昝肖肖. 胶州湾中部海域秋、冬季大型无脊椎动物群落结构及多样性研究. 中国海洋大学学报, 2010, 40(3): 78-84. |
| [33] | 梅春, 徐宾铎, 薛莹, 任一平, 昝肖肖. 胶州湾中部海域秋、冬季鱼类群落结构及其多样性研究. 中国水产科学, 2010, 17(1): 110-118. |
| [34] | 徐宾铎, 张帆, 梅春, 任一平, 纪毓鹏, 薛莹. 胶州湾中部海域春、夏季鱼类群落结构特征. 应用生态学报, 2010, 21(6): 1558-1564. |
| [35] | 李忠义, 左涛, 戴芳群, 金显仕. 运用稳定同位素技术研究长江口及南黄海水域春季拖网渔获物的营养级. 中国水产科学, 2010, 17(1): 103-109. |
| [36] | DeNiro M J, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimica Acta, 1978, 42(5): 495-506. |
| [37] | 李广玉, 鲁静, 何拥军. 胶州湾浮游植物多样性及其与环境因子的关系. 海洋地质动态, 2005, 21(4): 10-13, 38-38. |
| [38] | 宋书群. 黄、东海浮游植物功能群研究[D]. 青岛: 中国科学院研究生院(海洋研究所), 2010. |
| [39] | Vander Zanden M J, Cabana G, Rasmussen J B. Comparing trophic position of freshwater fish calculated using stable nitrogen isotope ratios (δ15N) and literature dietary data. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(5): 1142-1158. |
| [40] | 万祎, 胡建英, 安立会, 安伟, 杨敏, 伊藤光明, 服部达也, 陶澍. 利用稳定氮和碳同位素分析渤海湾食物网主要生物种的营养层次. 科学通报, 2005, 50(7): 708-712. |
| [41] | 张波, 唐启升. 渤、黄、东海高营养层次重要生物资源种类的营养级研究. 海洋科学进展, 2004, 22(4): 393-404. |
| [42] | Pauly D, Christensen V, Dalsgaard J, Froese R, Torres F Jr. Fishing down marine food webs. Science, 1998, 279(5352): 860-863. |
| [43] | Pauly D, Palomares M L, Froese R, Sa-a P, Vakily M, Preikshot D, Wallace S. Fishing down Canadian aquatic food webs. Canadian Journal of Fisheries and Aquatic Sciences, 2001, 58(1): 51-62. |
| [44] | Tang Q S, Jin X S, Wang J, Zhuang Z M, Cui Y, Meng T X. Decadal-scale variations of ecosystem productivity and control mechanisms in the Bohai Sea. Fisheries Oceanography, 2003, 12(4/5): 223-233. |