 2015, Vol. 35
2015, Vol. 35文章信息
- 张雪妮, 吕光辉, 王庭权, 马玉, 阿布里孜·阿不都热合, 赵晓英, 郭振洁, 朱修逸
- ZHANG Xueni, Lü Guanghui, WANG Tingquan, ZHAO Xiaoying, GUO Zhenjie, ZHU Xiuyi, GUO ZhenJie, ZHU XiuYI
- 荒漠区垂直河岸带植物多样性格局及其成因
- Patterns and mechanisms influencing plant diversity in an arid desert region riparian zone of the Aqikesu River, Xinjiang, China
- 生态学报, 2015, 35(18): 5966-5974
- Acta Ecologica Sinica, 2015, 35(18): 5966-5974
- http://dx.doi.org/10.5846/stxb201401160128
-
文章历史
- 收稿日期:2014-01-16
- 网络出版日期:2014-11-19






2. 绿洲生态教育部重点实验室, 乌鲁木齐 830046;
3. 新疆大学干旱生态环境研究所, 乌鲁木齐 830046
2. Key Laboratory of Oasis Ecology, Education Ministry, Urumqi 830046, China;
3. Institute of Arid Ecology and Environment, Xinjiang University, Urumqi 830046, China
植物多样性是区域生态系统功能和稳定的基础[1, 2],其分布格局和成因一直是群落生态学研究的重点问题之一[3, 4]。目前,国内关于陆生植物种多样性的研究多集中在山地、草原及交错带生态系统,较少关注河流廊道体系[5, 6],对干旱区内陆河垂直河岸带的植物多样性格局研究亦鲜见报道。河流及河岸带对于生物多样性保护、生态恢复和群落稳定性维持等十分重要[7, 8, 9, 10],在以缺水为显著特征的干旱、半干旱地区,河流及河岸带的生态地位不仅与荒漠河岸林的发生和演替息息相关,而且在连结生境斑块,稳定和维持区域生态系统安全过程中具有特殊意义[11, 12, 13]。荒漠河岸林是干旱区内陆河流域河流廊道植被类型的主体,在生态结构、功能及植被景观格局中占主导地位[11],因此,对不同区域或尺度下河岸带植被结构、多样性动态变化规律及其影响机制的研究应予以重视[5, 7]。
阿其克苏河位于新疆艾比湖国家级湿地自然保护区内,河岸分布着保护区的大部分胡杨,以胡杨为建群种的河岸植被群落形成了一条天然的防护带[14],对该区域的植物多样性保护、荒漠化防治和脆弱生态系统的维持等方面具有至关重要的作用。有研究表明,在干旱区随着离河道距离增加,植物种类分布和更替表现出一定的响应规律[15, 16],河流是维系荒漠河岸林天然植被的最主要因子[17]。然而,目前有关干旱区内陆河河岸植被的研究主要为河流断面上植被结构或多样性特征[9, 12, 18],以及多样性动态与单一因素如地下水或水因子间的关系等[15, 17, 19, 20],对垂直河流样带上的植物多样性格局及其土壤水盐和营养的交互影响机制关注较少。在干旱区,垂直河岸带不同距离区域的植物多样性格局如何?土壤水分梯度及其与土壤盐分和营养水平间对植物多样性的作用机制又将如何变化?上述问题目前尚需解答。基于此以阿其克苏河为例,验证和回答以下假设和问题:沿离河距离梯度(1)植物α、β多样性是否随旱胁迫加剧而递减?(2)不同距离梯度上土壤水分、盐碱性和C、P营养与多样性间有何关系,如何影响植物多样性的格局?最后基于对(1)、(2)问题的解答和河岸带、缓冲带宽度的建议,(3)提出阿其克苏河河岸带的具体管理措施,以期为充分发挥干旱区河岸带在植物多样性保护、荒漠化防治以及生态系统稳定性维持中的作用提供理论参考,同时也为河岸带和缓冲区设计与管理、河流的保护等提供理论依据[5, 21]。
1 材料和方法 1.1 研究区概况艾比湖湿地国家级自然保护区(44°30′—45°09′ N,82°36′—83°50′ E)位于新疆精河县西北、准噶尔盆地西南,是准噶尔盆地西南缘最低洼地和水盐汇集中心。本区为典型大陆性气候,干燥少雨多风,冬、夏季漫长,春、秋季短暂,年平均气温5 ℃。降水量年内分配不均匀,夏多冬少,多年平均降水量为105.17 mm,蒸发量为1315 mm。艾比湖特殊的湿地生态环境有其独特的生物资源多样性,主要植物种类包括近河岸的胡杨(Populus euphratica Oliv.)、柽柳(Tamarix ramosissima Ldb.)、梭梭(Haloxylon ammodendron (C.A.Mey.)Bunge)、盐豆木(Halimodendron halodendron (Pall.)Voss.)和芦苇(Phragmites australis(Cav.)Trin.exSteud.),湖滨盐沼地的盐穗木(Halostachys caspica C.A.Mey.exSchrenk)、盐节木(Halocnemum strobilaceum(Pall)M.Bieb)、碱蓬(Suaeda glauca (Bunge)Bunge)和盐爪爪(Kalidium foliatum(Pall.)Moq.),平原低地还分布有甘草(Glycyrrhiza uralensis Fisch.)、黑果枸杞(Lycium ruthenicum Murr.)、小獐毛(Aeluropus pungens (M.Bieb)C.Koch.),山前冲积洪扇有琵琶柴(Reaumuria soongorica (Pall.)Maxim.)等[22, 23]。阿其克苏河位于保护区内的湖区东侧,是艾比湖的水源之一,河岸及近处的古河道潜水溢出带分布有大量胡杨、柽柳、梭梭、盐豆木等典型植物,近年来,由于人类活动和气候变化的影响,阿其克苏河几近干涸,河岸植被面临退化威胁。
1.2 植被调查、采样与样品分析在保护区内东大桥管护站以南和以北,垂直阿其克苏河距离为0.1—0.2 km、0.2—0.4 km、0.9—1.5 km和2.0—3.0 km区域设置样带,分别以距离终点为下标,记为T0.2、T0.4、T1.5和T3.0。各样带土壤(0—15 cm)属性和海拔见表 1,分析表明各样带间土壤水分含量(SWC)、土壤有机质(SOM)和土壤碳磷比(C/P)有显著差异(P<0.05)。根据群落特征(乔木10 m×10 m,灌木5 m×5 m,草本1 m×1 m)分别在T0.2样带调查样方9个(乔、灌木和草本分别3、4和2个),T0.4样带6个(乔、灌木和草本各2个),T1.5样带8个(乔木和草本2个,灌木4个),T3.0样带8个(乔木2个,灌木和草本各3个),样方间距离大于20 m。现场鉴定样方中的物种类别,并采集标本带回标本室进行重复鉴定,记录种数、个体数、胸径、高度、冠幅等植物特征,以及各样地的海拔、经纬度、群落微环境和地貌特征,同时在调查样方内利用四分法取0—15 cm土壤作为土壤样品。土壤指标及测定方法分别为:土壤水分采用烘干法,全盐采用残渣烘干法(土水比1 ∶ 5),pH值采用酸度计法,土壤有机质采用重铬酸钾容量法,土壤全磷采用HClO4-H2SO4钼锑抗比色法[24]。
| 指标 Index | 样带Transect | |||
| T0.2 | T0.4 | T1.5 | T3.0 | |
| 同一指标的不同字母表示样带间差异显著,无标示即无显著差异(P<0.05) | ||||
| 土壤水分含量Soil water content/% | 17.55a | 7.82b | 7.50b | 7.01b |
| 全盐Total salt/% | 1.12 | 0.56 | 0.89 | 0.54 |
| pH值pH value | 8.67 | 8.73 | 8.69 | 8.42 |
| 土壤有机质Soil organic matter/% | 2.33a | 1.02b | 1.51ab | 1.98ab |
| 土壤全磷Phosphorus/(g/kg) | 0.33 | 0.37 | 0.35 | 0.39 |
| 碳磷比C/P ratio | 44.37a | 15.77b | 26.07ab | 30.16ab |
| 海拔Elevation/m | 281.67 | 268.50 | 266.13 | 296.50 |
物种累积曲线(Species Accumulation Curves,SAC)显示了连续抽样下新物种出现的速率,是分析调查样地物种组成和预测物种丰富度的有效工具,被广泛用于抽样量充分性、合理性的判断以及物种丰富度的估计[25, 26];SAC和物种丰富度期望值(Abundance-based Coverage Estimator,ACE;100次随机化物种增加顺序)在EstimateS 9中完成。
多样性指数利用R语言的vegan程序包计算;方差分析(GLM过程,Duncan法多重比较)、主成分分析在SAS8.0中完成。群落本质多样性排序利用目前较推崇的“右尾和”曲线分析[27, 28],其数据计算公式为:
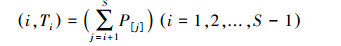
式中,(i,Ti)为种秩及其对应的本质多样性值,S为物种数,P[j]为按照降序排列的各物种相对多度;以i为横坐标,Ti为纵坐标绘图,即可得到各样带本质多样性。
多样性的影响因素分析采取多元回归和通径分析相结合的方法解释,不仅起到互补作用,还可更全面地剖析研究资料的内含信息[29]。其中,直接通径系数表明各影响因子对因变量的直接影响程度,代表了因变量在自变量方向上的标准变化速率;间接通径系数表明单因子通过其他因子对因变量的影响程度;决策系数既反映了单因子对因变量的直接决定作用,又反映了与单因子有关的通径对因变量的决定作用,将决策系数值由大到小排序,可以反映出各单因子对因变量的综合作用大小[30]。
2 结果与分析 2.1 各样带物种累积曲线及植物生活型组成由各样带物种(估计值)累积曲线可知(图 1),随着抽样数增加,样带中物种数均趋于平缓,并且实际调查的物种数占ACE的比例较高,达到88.7%以上(T0.2—T3.0依次为19/20.7、12/13.4、23/25.0和23/23.9),说明抽样能够代表各样带的物种组成,样地中的大多数物种被采集到,效果较好。

|
| 图 1 各样带的物种累计曲线 Fig. 1 Species cumulative curve of each transect 虚线表示物种数的95%置信区间 |
各样带共调查到30个植物种,T0.2—T3.0分别为19、12、23和23种(T3.0忽略1种藤本),其中大部分为(强)盐、旱生或盐旱中生类,T0.2—T3.0的比例分别达到84.2% (16/19)、100.0% (12/12)、91.3% (21/23)和91.3% (21/23),其余为盐生湿生或湿生类;统计各样带植物生活型比例可以看出(图 2),除T0.4外,各样带乔木、灌木和草本植物比例变化均较小,相较于T0.2,T0.4的乔木、灌木比例增加和草本比例降低幅度均较大,表现出一定的过渡带性质,这可能说明离河距离大于0.2 km时植物生长和组成基本不受河流(地下水)影响;并且表 1中土壤属性在T0.2到T0.4间的差异也反映了T0.4的过渡性特征。

|
| 图 2 各样带植物生活型比例 (种数) Fig. 2 Plant life form proportion of each transect 图中数据为相应生活型的比例,括号内为对应的种数 |
群落本质多样性曲线是一种与指数无关的多样性比较方法,只有本质多样性高(多样性曲线不相交)的群落才具有高的多样性[31]。由各样带群落多样性曲线可以看出(图 3),群落本质多样性排序为T3.0>T1.5>T0.2 >T0.4,说明离河较远区域的群落多样性本质上高于近河区域;T1.5和T3.0样带的曲线末端有部分重叠,表明样带间α多样性指数的排序会存在差别。

|
| 图 3 各样带“右尾和”多样性曲线 Fig. 3 Right tail-sum diversity curve of each transect |
对于α多样性指数(表 2),Shannon-Wiener指数、Simpson指数、Pielou指数和Margalef指数在样带间均有显著或近乎显著的差异(P<0.05或P<0.1),且多样性排序均为T3.0≥T1.5>T0.2>T0.4;样带间参数α指数存在显著差异(P<0.05),但排序为T1.5>T3.0>T0.2>T0.4,这与T1.5和T3.0本质多样性曲线有部分重合相符。
| 多样性指数 Diversity Index | 样带 Transect | ||||
| T0.2 | T0.4 | T1.5 | T3.0 | ||
| 下划线字母表示在P<0.1水平有差异 | |||||
| α多样性 | Shannon-Wiener | 1.05ab | 0.71b | 1.17ab | 1.22a |
| α Diversity | Simpson | 0.52ab | 0.39b | 0.59a | 0.59a |
| α | 1.36ab | 0.81b | 1.68a | 1.59ab | |
| Pielou | 0.63ab | 0.50b | 0.68a | 0.68a | |
| Margalef | 1.02ab | 0.64b | 1.18a | 1.19a | |
| β多样性 | Cody | 3.00ab | 1.80b | 4.00a | 3.57ab |
| β Diversity | Wilson | 0.52ab | 0.36b | 0.67a | 0.48ab |
| Jaccard | 0.33ab | 0.50a | 0.21b | 0.39ab | |
| Srenson | 0.48ab | 0.64a | 0.33b | 0.52ab | |
β多样性的Cody指数和Wilson指数反映环境/生境梯度上物种的替代速率[4, 32, 33]。T0.2至T3.0的物种替代率具有波动性,样带间差异显著(P<0.05),其中T0.4与T1.5间差异最大,随着离河距离增加,物种的更替速率先降后升(表 2)。Jaccard指数和Srenson指数表征不同群落-生境的物种相似性,T0.4物种相似性显著高于其他样带(P<0.05),T1.5最低。β多样性的上述特征可能说明,T0.4样带是受河流潜在影响(地下水)的边缘带,而T1.5已过渡至不受河流影响的区域,植物种变化较大,因此物种相似性降低。
2.3 影响植物多样性的土壤因素样带变化为便于分析多样性的影响机制,对5个α多样性指数进行主成分分析以综合其信息(表 3),第一主成分Z1(特征值为3.94)的贡献率达78.79%,与各多样性特征向量均为极显著相关(P<0.01),相关性较高,因此能够代表多样性的信息。以α多样性指数的第一主成分Z1为因变量,6个土壤属性数据(即SWC、TS、pH、SOM、P和C/P)为自变量,通过逐步回归分析确定影响多样性的土壤因子(P<0.05),并计算其直接通径系数、间接通径系数和决策系数,分析选入因子对多样性的直接和间接影响,以及评价各因子的作用大小(表 4)。
| 项目 Item | 特征值 Eigenvalue | 特征向量Eigenvector | ||||
| Shannon-Wiener | Simpson | Margalef | α | Pielou | ||
| 第一主成分First principal component | 3.94(78.79%) | 0.50 | 0.48 | 0.45 | 0.44 | 0.35 |
| 相关系数Correlation coefficient(P<0.0001) | 0.98 | 0.95 | 0.89 | 0.88** | 0.70 | |
| 样带及影响因素 Transects and factors | 直接系数 Direct coefficient | 间接通径系数Indirect coefficient | 决策系数 Decision coefficien | ||||
| * *P<0.01,*P<0.05,下划线数字P<0.1 | |||||||
| T0.2 | SWC | C/P | TS | pH | SOM | ||
| SWC | -1.43** | — | 0.48 | 0.65 | 0.38 | -0.39 | -1.15 |
| C/P | 2.74** | -0.25 | — | 0.31 | -0.62 | -2.05 | -6.79 |
| TS | 1.92* | -0.49 | 0.44 | — | -1.83 | -0.63 | -5.98 |
| pH | -2.33** | 0.23 | 0.72 | 1.51 | — | -0.67 | -2.95 |
| SOM | -2.18* | -0.26 | 2.58 | 0.56 | -0.72 | — | -4.66 |
| T0.4 | pH | P | |||||
| pH | 1.34* | — | -1.53 | — | — | — | -2.30 |
| P | 1.82* | -1.13 | — | — | — | — | -0.79 |
| T1.5 | TS | P | pH | ||||
| TS | -0.63 | — | 0.83 | 0.01 | — | — | -0.66 |
| P | 0.97 | -0.54 | — | -0.01 | — | — | -0.13 |
| pH | 0.13 | -0.03 | -0.08 | — | — | — | -0.01 |
| T3.0 | SWC | P | |||||
| SWC | 0.66* | — | -0.01 | — | — | — | 0.42 |
| P | 0.20 | -0.02 | — | — | — | — | 0.04 |
通径分析表明(表 4),影响T0.2植物多样性的土壤因素有SWC、C/P、TS、pH和SOM,且各因素间交互影响途径复杂;各因素对多样性的直接作用为C/P>pH>SOM>TS>SWC,间接作用主要有SOM→C/P和pH→TS及其反向途径(绝对值>1),其间接通径大多较接近或大于其直接通径,说明各因素及其交互影响对植物多样性的作用机制同等重要;由决策系数(绝对值)可知C/P、TS和SOM是影响该样带植物多样性的主要因素,pH和SWC为次要因素;上述说明,土壤贫营养和高盐分是影响T0.2植物多样性的主要原因,而SWC对该样带多样性的决定作用较不明显。
P与pH是直接影响T0.4植物多样性的土壤因素,pH→P强于P→pH,且pH→P的间接作用大于其直接通径,因此pH对该样带多样性的决定性较P更大。P、TS与pH是依次影响T1.5植物多样性的土壤因素,TS→P对多样性有较强的间接作用,反之亦然,因此二者是影响该样带多样性的主要因素。影响T3.0的主要因素为SWC和P,二者相互的间接通径均较弱,由决策系数可知,SWC是决定该样带植物多样性的重要因素。由T0.4—T3.0样带土壤属性对植物多样性的影响地位和途径可知,土壤P含量、盐分(pH、TS)和土壤水分在距河较远区域对植物多样性的决定性增强,其中土壤盐分水平(pH、TS)对T0.4—T1.5的P供应状况影响较大,因此在研究该区营养元素特别是P对植物多样性的影响方面,需要对土壤的盐分特征及其间接影响加以重视。
3 结论与讨论 3.1 植物多样性及其影响因素变化与河岸带宽度的确定研究区离河不同距离样带上的植物多样性格局显示,距河较近样带上(T0.2)植物多样性并非最高,无论其本质多样性亦或测度富集种或稀有种的多样性和均匀度的各α多样性指数[34, 35]均表明距河0.9—1.5 km(T1.5)和2.0—3.0 km(T3.0)区域的植物多样性普遍高于近河区域(T0.2);土壤水分在干旱区生态过程中有特殊重要性,理论上在一定范围内距河道越近,干旱胁迫越弱,植物多样性应越高[36],但上述显然与预期不同,这可能与高地下水位下土壤盐分的动态有关。在近河区域(T0.2),地下水埋深约1.63—2.67 m[14],水位相对较高,这对深根系、水分需求高的植物如胡杨、柽柳等的生长较有利[37],但高地下水位加之强烈的地表蒸发又导致近河岸带浅层土盐分的集聚,使得一些植物面临盐胁迫[38],特别是对浅根系、不耐盐碱植物(草本或小型灌木等)较不利,T0.2样带土壤水分较高,且盐分平均达到1.12%(表 1),属于中—重度盐渍化[39],部分植物生长受限,这与该样带较低的(强)盐、旱生或盐旱中生类植物比例相符(84.2%);T0.2样带土壤水分的直接限制和通过盐分的间接正向作用也反映了该样带土壤水盐条件对植物多样性的上述影响,同时该样带植物多样性还受C/P、pH和SOM的直接和复杂的交互影响,这与近河样带群落生境较高的异质性和复杂性有关[16]。
距河0.2—0.4 km区域(T0.4)的植物多样性本质上最低,α多样性指数也表明无论富集种、稀疏种多样性亦或物种均匀度均显著低于其他区域,同时β多样性表明该区域物种相异性最低,相似性最高;该样带较低的多样性与塔河绿洲—荒漠过渡带相似[40, 41],这主要与该样带土壤水盐和有机质较T0.2显著或明显降低、土壤性质变异大有关[42, 43];此外T0.4样带植被无湿生类型,且该样带草本比例较T0.2大幅降低也与相关研究一致[15, 44];影响因素方面,该样带对多样性具显著影响的土壤因素也由T0.2的水分、盐分和营养5个指标转变为仅含土壤pH值和P含量;综合上述,T0.4无论在植被构成、多样性还是土壤属性及其影响方面均表现出较明显的缓冲或过渡带特征,这可能说明在距河0.2 km以外的区域上,阿其克苏河对研究区植被已无影响,因此可将距河0.2—0.4 km区域(T0.4)作为确定河岸带和过渡带宽度的参考样带,这对于明确河岸带林分宽度、合理保护和管理河岸带生态系统以及促进河岸林的更新具有重要意义[45]。
T1.5—T3.0样带植物主要以(强)盐、旱生或盐旱中生类为主(91.3%),其本质多样性和α多样性普遍高于近河区域,同时T1.5物种更替速率和相似性与T0.4间的差异较大,说明随着离河距离由0.2—0.4 km范围增加至0.9—1.5 km区域,物种组成差异增大,这与该生境下的土壤环境密切相关;T1.5、T3.0土壤水分均较低(7.0%—7.5%),有机营养较T0.4升高(C/P分别为26.07和30.16),盐分略升但与T0.4相同均低于1.0%,土壤属性在上述范围内可能有助于维持或提高植物多样性;土壤P与盐分是影响T1.5植物多样性的最主要因素,随着离河距离持续增加,土壤水分和土壤P对植物多样性的影响逐渐显著(T3.0),反映了旱胁迫加剧时土壤水分和营养的重要性增强。
从土壤因素对植物多样性的作用来看,随着离河距离增加,影响T0.4—T3.0样带植物多样性的因素减少,但均包括土壤P,说明在水分限制强烈的生境下,营养元素P的限制更加突出,同时土壤盐分和水分随着距离增加逐渐成为影响植物多样性的主导因素,包括通过P产生的间接影响,这在保护区今后的植被和土地资源管理和保护中应予以考虑。
3.2 对河岸带管理和生物多样性保护措施的建议鉴于河岸带位置的特殊性及其在涵养水源,过滤和截留沉积物及营养物质,以及维持生物多样性和生态平衡等方面的重要功能[46, 47],对干旱区河岸带的管理和生物多样性的保护应受到有关部门的重视。首先,阿其克苏河河岸带宽度可能在距河0.2 km范围,保护区在确定河岸带宽度、建设(如道路建设)和管理河岸带过程中可将0.2 km作为参考。其次,在河流影响范围外的区域(0.2—0.4 km或更远),土壤P养分和水分对植被动态的决定作用更突出,因此在植被恢复以及利用植物资源修复盐渍化土壤实践中应结合土壤水分条件选取植物种类,在旱胁迫严重的生境下(如土壤水分接近7.8%)应选择耐旱且具有较强P吸收能力或P需求较低的物种。再次,土壤水分、盐分动态及其与盐分的交互作用是影响该地区植物多样性的主要因素,且随着离河距离增加其影响更显著,因此对河岸带的植被保护应综合考虑水分、盐分和营养间互作对生物多样性的影响机制;根据垂直河岸样带多样性格局的特征,土壤水分在约7.0—7.5%之间,土壤盐分低于1.0%,C/P在26.1—30.2之间时植物多样性较高,因此从管理角度出发的受损生态系统的恢复与重建,可重点考虑上述主要环境因子及其范围,进而制定有效的局部尺度(河岸带)生物多样性保护管理对策。
| [1] | Maestre F T, Quero J L, Gotelli N J, Escudero A, Ochoa V, Baquerizo M D, Gómez M G, Bowker M A, Soliveres S, Escolar C, Palacios P G, Berdugo M, Valencia E, Gozalo B, Gallardo A, Aguilera L, Arredondo T, Blones J, Boeken B, Bran D, Conceição A A, Cabrera O, Chaieb M, Derak M, Eldridge D J, Espinosa C I, Florentino A, Gaitán J, Gatica M G, Ghiloufi W, González S G, Gutiérrez J R, Hernández R M, Huang X W, Sannwald E H, Jankju M, Miriti M, Monerris J, Mau R L, Morici E, Naseri K, Ospina A, Polo V, Prina A, Pucheta E, Collantes D A R, Romão R, Tighe M, Díaz C T, Val J, Veiga J P, Wang D L, Zaady E. Plant species richness and ecosystem multifunctionality in global drylands. Science, 2012, 335(6065):214-218. |
| [2] | Loïc M T, Sean R C. Understanding diversity-stability relationships: towards a unified model of portfolio effects. Ecology Letters,2013,16:140-150. |
| [3] | 李旭华,邓永利,张峰,董刚,李世广.山西庞泉沟自然保护区森林群落物种多样性.生态学杂志,2013,32(7):1667-1673. |
| [4] | 杨洁,余华光,徐凤洁,马明睿,由文辉.崇明东滩围垦区草本植物群落组成及物种多样性.生态学杂志,2013,32(7):1748-1755. |
| [5] | 邓红兵,王青春,王庆礼,吴文春,邵国凡.河岸植被缓冲带与河岸带管理.应用生态学报, 2001,12(6):951-954. |
| [6] | 赵鸣飞,康慕谊,刘全儒,邢开雄,王菁兰,孟世勇.东江干流河岸带植物多样性分布规律及影响因素.资源科学,2013,35(3):488-495. |
| [7] | 郭二辉,孙然好,陈利顶.河岸植被缓冲带主要生态服务功能研究的现状与展望.生态学杂志,2011,30(8):1830-1837. |
| [8] | 修晨, 欧阳志云,郑华,王大尚.北京永定河-海河干流河岸带植物的区系分析.生态学报, 2014,34(6):1535-1547. |
| [9] | 高润梅,郭晋平,郭跃东,张东旭.文峪河上游河岸林的群落结构与多样性特征.林业科学研究,2011,24(1):74-81. |
| [10] | Soykan C U, Brand L A, Ries L, Stromberg J C, Hass C, Simmons D A, Patterson W J D, Sabo J L. Multitaxonomic diversity patterns along a desert riparian-upland gradient. PLoS ONE,2012,7(1): e28235. |
| [11] | 赵良菊,肖洪浪,程国栋,宋耀选,赵亮,李彩芝,杨秋.黑河下游河岸林植物水分来源初步研究.地球学报,2008,29(6):709-718. |
| [12] | 郭春秀,刘世增,杨自辉,刘虎俊,李银科,刘淑娟,张莹花,韩富贵.石羊河中下游河岸植被的结构及其功能研究.中国农学通报,2011,27(28):115-120. |
| [13] | 尹力,赵良菊,阮云峰,肖洪浪,程国栋,周茅先,王芳,李彩芝.黑河下游典型生态系统水分补给源及优势植物水分来源研究.冰川冻土,2012,34(6):1478-1486. |
| [14] | 赵枫,金海龙.艾比湖湿地自然保护区阿其克苏河地下水特征与胡杨生长关系研究.干旱区资源与环境,2011,25(8):156-160. |
| [15] | 陈亚宁,张宏锋,李卫红,陈亚鹏.新疆塔里木河下游物种多样性变化与地下水位的关系.地球科学进展,2005,20(2):158-165. |
| [16] | 刘加珍,李卫红,吴纯渊,陈永金.荒漠河岸生态系统退化与物种多样性恢复研究.干旱区研究,2009,26(2):212-220. |
| [17] | 李卫红,郝兴明,覃新闻,陈亚宁,黄湘.干旱区内陆河流域荒漠河岸林群落生态过程与水文机制研究.中国沙漠,2008,28(6):1113-1117. |
| [18] | 卢磊,赵振勇,乔木,张科,周生斌,唐金.塔里木河中游河岸带植物区系及物种多样性初探.水土保持通报,2011,31(1):16-21. |
| [19] | 席海洋,冯起,司建华,常宗强,苏永红.黑河下游绿洲NDVI对地下水位变化的响应研究.中国沙漠,2013,33(2):574-582. |
| [20] | 陈永金,刘加珍,陈亚宁,李新功,朱海勇.输水前后塔里木河下游物种多样性与水因子的关系.生态学报,2013,33(7):2212-2224. |
| [21] | 王青春,邓红兵,王庆礼.基于生物多样性保护的河岸带植被管理对策-以长白山二道白河为例.生态学杂志,2006,25(6):682-685. |
| [22] | 杨晓东,吕光辉,田幼华,杨军,张雪梅.新疆艾比湖湿地自然保护区植物的生态分组.生态学杂志,2009,28(12):2489-2494. |
| [23] | 米吉提·胡达拜尔地,徐建国.新疆高等植物检索表.乌鲁木齐:新疆大学出版社,2000. |
| [24] | 南京农业大学.土壤农化分析.北京:农业出版社,1996:126-128. |
| [25] | 孙志强,张星耀,朱彦鹏,梁军,于善栋,张英军,杨晓燕.应用物种指示值法解析昆嵛山植物群落类型和植物多样性.生态学报,2011,31(11):3120-3132. |
| [26] | 孙志强,朱彦鹏,张星耀,林琳,梁军,车吉明.采用Rényi多样性曲线法评价昆嵛山林下灌草层多样性排序效果.南京林业大学学报:自然科学版,2012,36(1):73-78. |
| [27] | 刘灿然,马克平.多样性排序:方法与实例.植物生态学报,2002,26(增刊):63-67. |
| [28] | Liu C R, Whittaker R J, Ma K P, Malcolm J R. Unifying and distinguishing diversity ordering methods for comparing communities. Population Ecology,2007, 49:89-100. |
| [29] | 张永泉,白庆利,郭文学,丰程程,尹家胜.洛氏鱥形态性状对体质量的影响.生态学杂志, 2013,32(11):3063-3068. |
| [30] | 姚勇哲,李建明,张荣,孙三杰,陈凯利.温室番茄蒸腾量与其影响因子的相关分析及模型模拟.应用生态学报,2012,23(7):1869-1874. |
| [31] | 雷相东,张会儒,李冬兰,梁毓照.东北过伐林区四种森林类型的物种多样性比较研究.生态学杂志, 2003,22(5):47-50. |
| [32] | 马克平,刘灿然,刘玉明.生物群落多样性的测度方法Ⅱ.β多样性的测度方法.生物多样性,1995,3(1):38-43. |
| [33] | 何艳华,闫明,张钦弟,张直峰,苗艳明,毕润成.五鹿山国家级自然保护区物种多样性海拔格局.生态学报,2013,33(8):2452-2462. |
| [34] | 马克平.生物群落多样性的测度方法Ⅰ.α多样性的测度方法(上).生物多样性,1994, 2(3):162-168. |
| [35] | 马克平,刘玉明.生物群落多样性的测度方法Ⅰ.α多样性的测度方法(下).生物多样性,1994,2(4):231-239. |
| [36] | 李新荣,何明珠,贾荣亮.黑河中下游荒漠区植物多样性分布对土壤水分变化的响应.地球科学进展,2008,23(7):685-691. |
| [37] | 赵文智,刘鹄.荒漠区植被对地下水埋深响应研究进展.生态学报,2006,26(8):2702-2708. |
| [38] | 郑丹,李卫红,陈亚鹏,刘加珍.干旱区地下水与天然植被关系研究综述.资源科学,2005,27 (4):160-167. |
| [39] | 张雪妮,吕光辉,杨晓东,贡璐,秦璐,何学敏,刘昊奇.基于盐分梯度的荒漠植物多样性与群落、种间联接响应.生态学报,2013,33(18):5714-5722. |
| [40] | 郑田,李卫红,李建贵,湾疆辉.塔里木河下游绿洲荒漠过渡带群落多样性特征分析.中国沙漠, 2009, 29(2): 241-247. |
| [41] | 周洪华,陈亚宁,李卫红.塔里木河下游绿洲-荒漠过渡带植物多样性特征及优势种群分布格局.中国沙漠,2009,29(4): 688-696. |
| [42] | 李朝生,慈龙骏,于春堂,杨晓晖,王忠,白飞.沙漠--河岸过渡带土壤养分与盐分的空间变异.农业环境科学学报,2006,24(4):643-647. |
| [43] | 李银科,刘世增,刘虎俊,魏怀东,杨自辉,张莹花,刘淑娟.石羊河流域中下游沙漠-河岸过渡带土壤特征研究.水土保持研究,2012,19(6):29-35. |
| [44] | 白元,徐海量,张鹏,赵新风,傅荩仪.塔里木河下游荒漠植物群落物种多样性及其结构特征分析.生态与农村环境学报,2012,28(5):486-492. |
| [45] | 席庆,尚杨娟,李明阳.下李林场河岸林林分过渡带宽度划分方法.林业调查规划,2009, 33(6):47-50. |
| [46] | 张建春,彭补拙.河岸带研究及其退化生态系统的恢复与重建.生态学报,2003,23(1):56-63. |
| [47] | 岳隽,王仰麟.国内外河岸带研究的进展与展望.地理科学进展,2005,24(5):33-40. |