 2015, Vol. 35
2015, Vol. 35文章信息
- 陈刚, 涂利华, 彭勇, 胡红玲, 胡庭兴
- CHEN Gang, TU Lihua, PENG Yong, HU Hongling, HU Tingxing
- 瓦屋山扁刺栲-中华木荷常绿阔叶次生林土壤有机碳组分特征
- Characteristics of soil organic carbon components in a secondary Castanopsis platyacantha-Schima sinensis evergreen broad-leaved forest, Wawushan Mountain
- 生态学报, 2015, 35(18): 6100-6109
- Acta Ecologica Sinica, 2015, 35(18): 6100-6109
- http://dx.doi.org/10.5846/stxb201401010005
-
文章历史
- 收稿日期: 2014-01-01
- 网络出版日期: 2014-11-19






次生林是原始森林由于人类活动被完全清除后,自然状态下自主生长的植物群落[1, 2]。我国现有森林面积为2.07亿hm2,其中原生林面积为0.12亿hm2,次生林面积为1.18亿hm2,分别占森林总面积的6%和57%[3],次生林已成为我国现今森林资源的主体。次生林是全球重要碳汇,约为0.35—0.6 Gt C/a[4](1 Gt=1015 g),并且是陆地生物圈近几十年来净碳汇增加的重要原因[5]。常绿阔叶林作为我国亚热带地区的顶级群落,其森林结构复杂、生产力高、生物多样性丰富,但由于人类活动的干扰和破坏,目前我国亚热带天然常绿阔叶林已接近消失,取而代之的是大面积的次生林[6]。常绿阔叶次生林植物物种丰富,在生物多样性保护,水土保持和碳固定方面具有重要作用,并且,我国绝大多数亚热带次生林尚处于较年轻的阶段,具有很大的生长和固碳潜力[6]。
土壤是全球最大的有机碳库,最新研究结果显示全球土壤有机碳库大于3300 Gt,其微小的变化可能引起大气CO2浓度的较大波动,进而影响温室效应和全球气候变化[7, 8]。因此,近年来,许多生态学家都致力于各类陆地生态系统土壤有机碳贮存和分配的研究[9, 10, 11, 12],以期为区域性土壤有机碳库的估算提供基础数据。尽管如此,由于地域性研究对象复杂而多样,基础数据并不全面、详细、可靠,为我国乃至全球范围内的有机碳库估算带来很大局限性[13]。
由于土壤碳库基数和自然变异大,碳库微小的变化很难被发现[14]。Biederbeck等[15]研究指出,土壤有机碳的短暂波动主要来源于易氧化、易分解部分的变化,并选择了易氧化碳、微生物生物量碳和可矿化碳作为土壤活性有机碳的指示因子。虽然在土壤有机碳含量中土壤活性有机碳只占很小比例,但它作为土壤生物化学转换的直接碳源,并且能够在土壤全碳变化之前反映土壤的微小变化[14, 16]。因此,活性碳组分研究在土壤有机碳研究中具有重要意义。针对各地域性土壤开展详尽的有机碳贮存及活性有机碳分配特征的研究意义重大。
本研究以瓦屋山中山段典型扁刺栲(扁刺锥)(Castanopsis platyacantha)-中华木荷(Schima sinensis)常绿阔叶次生林地为对象,探讨土壤有机碳及其活性组分的含量、储量和在土层中的垂直分布,为区域碳循环研究提供基础数据。
1 材料与方法 1.1 试验地概况位于四川盆地向川西高原过渡的四川盆地西部边缘地区,由于其高降水量和中亚热带湿润气候,被称为“华西雨屏”,东西宽 50—70 km,南北长 400—450 km,总面积约2.5万km2 [17]。试验地位于“华西雨屏”中心地带的四川洪雅县瓦屋山国家森林公园,广泛分布着中亚热带常绿阔叶树种,其中山段的次生林是1956年由原生的扁刺栲-中华木荷群落遭砍伐后自然恢复而成[18, 19]。试验地设置在瓦屋山中山区的木姜杠林班(29°32′35″N,103°15′41″E),海拔约1600m。该地段属中亚热带季风性湿润气候,温和湿润,日照短,雨水多,云雾多、湿度大。年均温10—14℃,年日照不足800 h,年降水日199—226 d,降水量达2398mm,年均相对空气湿度达85%—90%[19]。土壤类型为山地黄壤。该中山地段在破坏前均为顶极地带性常绿阔叶林扁刺栲-中华木荷群落。1956年遭到砍伐破坏,遗留下一些幼树幼苗,后基本上无进一步的干扰破坏而得以自然恢复,因此该林分树种林龄约为56a。目前已形成次生常绿阔叶成林,优势种为扁刺栲和中华木荷[18],草本层十分稀疏。
1.2 研究方法 1.2.1 凋落物及土壤样品的采集、处理和测定2 012年9月,根据典型性和代表性原则在木姜杠林班设置10块样方(20m×20m)。各样地调查基本情况如表 1所示。同年11月28日,采集凋落物和土壤样品。在每个样方内随机选择3个样点,量取 50cm×50cm的采样点,共计36个采样点,分别收集采样点内所有凋落物。在每个样方内随机挖取3个土壤剖面,共计36个土壤剖面。由于土壤腐殖质厚度约为10cm,故按0—10cm,10—40cm,40—70cm,70—100cm等4个层次采集各土层土壤样品,同时用环刀分层采样,每个剖面每个土层取3个环刀土样。
| 样地号 Quadrat Number | 坡度坡向 Slope and Aspect | 郁闭度 Canopy Density | 扁刺栲 株数 CL | 扁刺栲 平均胸径 Mean DBH of CL/cm | 中华 木荷株数 SS | 中华木荷 平均胸径 Mean DBH of SS/cm | 其他树种株数 Other Tree Spicies | 合计株数 Total |
| CL:Castanopsisp latyacantha 扁刺栲;SS:Schima sinense 中华木荷;DBH: Diameter at breast height 胸径; -:无数据 | ||||||||
| 1 | NE,9° | 0.8 | 19 | 18.0 | 24 | 25.1 | 3 | 46 |
| 2 | W,14° | 0.7 | 9 | 25.2 | 10 | 25.6 | 4 | 23 |
| 3 | NE,16° | 0.7 | 9 | 26.3 | 8 | 29.7 | 9 | 26 |
| 4 | NE,9° | 0.8 | 6 | 18.2 | 21 | 22.4 | 19 | 46 |
| 5 | WE,11° | 0.7 | 5 | 25.3 | 14 | 28.1 | 5 | 24 |
| 6 | WE,12° | 0.7 | 12 | 26.4 | 1 | 38.9 | 8 | 21 |
| 7 | N,9° | 0.8 | 18 | 23.0 | 0 | 10 | 28 | |
| 8 | NE,12° | 0.7 | 12 | 27.0 | 1 | 33.7 | 17 | 30 |
| 9 | N,5° | 0.8 | 8 | 15.6 | 9 | 21.8 | 17 | 34 |
| 10 | N,13° | 0.6 | 2 | 27.4 | 1 | 29.3 | 9 | 12 |
收集的凋落物样品,去除可见石砾及表面泥土,于65℃烘干至恒重并称量记录。各凋落物烘干样品按四分法分取一部分,粉碎过2mm筛后装袋,供化学分析使用。土样取小部分用于测量自然含水率,其余用四分法分出一半挑出可见根及石砾,过2mm筛,于4℃条件下保存待测,另一半土样自然风干,去除可见根系和石砾后研磨粉碎,分别过2mm和0.25mm筛,装袋待测。
测定项目有土壤pH值、土壤田间持水量、土壤总孔隙度、土壤容重、凋落物有机碳(LOC)、土壤有机碳(SOC)、土壤微生物生物量碳(MBC)、土壤可浸提溶解性有机碳(EDOC)和土壤易氧化碳(ROC)。土壤pH值采用氯化钾溶液浸提法进行测定(LY/T 1239—1999)。土壤田间持水量、总孔隙度和容重采用环刀法测定(LY/T 1215—1999)。凋落物和土壤有机碳采用重铬酸钾外加热法(LY/T 1237—1999)测定。土壤MBC采用氯仿熏蒸、0.5mol/L硫酸钾溶液浸提[20](做氯仿熏蒸和未熏蒸2种处理),总有机碳分析仪(TOC-VcPH+TNM-1,Shimazu Inc,Japan)测定总碳和无机碳,根据所测得的指标计算MBC,计算方公式为:
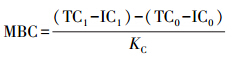
式中,MBC为微生物生物量碳,TC1为熏蒸样品总碳,IC1为熏蒸样品无机碳,TC0为未熏蒸样品总碳,IC0为未熏蒸样品无机碳,KC为微生物生物量碳转换系数(0.45 [20])。
EDOC采用0.5mol/L硫酸钾[21]浸提,浸提液过0.45 μm微孔滤膜后用总有机碳分析仪测定。ROC采用0.333mol/L高锰酸钾氧化法测定[14]。
1.2.2 数据计算与处理对土壤有机碳密度进行分层计算的方法:
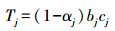
式中,Tj为第j层土壤有机碳密度(kg/m3);αj为第j层砾石(粒径>2mm)体积百分数,本研究中各土层砾石含量小于1%,以0%进行计算;bj为第j层平均土壤容重(g/cm3);cj为第j层总有机碳平均含量(g/kg)。
对0—100cm土层土壤有机碳储量计算的方法:
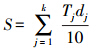
式中,S为土壤有机碳储量(t/hm2);dj为第j层土壤厚度(cm);10为单位换算系数;k为土壤层数。
为了比较各土层各指标之间的差异,利用 SPSS16. 0(SPSS Inc.,USA)对土层各指标进行方差分析(One-way ANOVA),并进行LSD多重比较(α=0.05),统计显著水平为α=0.05。用Sigmaplot 10.0(Systat software Inc.,USA)绘图。
2 结果与分析 2.1 不同土层深度土壤pH值和物理性质本研究结果表明,各土层土壤均呈酸性,潜在酸度3.2—4.0,田间持水量为63%—190%,总孔隙度和容重分别为63.0%—83.3%和0.41—0.99 g/cm3,土壤pH值和容重随土层深度的增加而升高,而土壤田间持水量和总孔隙度随土层的加深而降低(表 2)。各个土层pH值、总孔隙度和容重的变异数均较小,只有70—100cm土层田间持水量的变异系数略大(CV=35%),表明调查群落内土壤pH值和物理性质在水平方向上较为均一。4个土层间的pH值、总孔隙度和容重均存在显著差异,而土壤田间持水量除40—70cm和70—100cm两个土层间无显著差异外,其余各土层间均有显著差异,表明该林地内土壤pH值和物理性质在垂直方向上具有较大变异。其中0—10cm层的田间持水量均值是70—100cm层的3倍,而70—100cm层土壤容重为0—10cm层的2.4倍。
| 土层深度 Soil depth/cm | pH | 田间持水量/% Fieldmoisture capacity | 总孔隙度/% Total porosity | 容重/(g/cm3) Soil bulk density | ||||
| Mean±SD | CV/% | Mean±SD | CV/% | Mean±SD | CV/% | Mean±SD | CV/% | |
| Mean±SD: 平均值±标准差 Standard deviation; CV: 变异系数 Coefficient of variation; 同列相同写字母表示不同土层间差异不显著(单因素方差分析,LSD多重比较法,α= 0.05) | ||||||||
| 0—10 | 3.2±0.1d | 4.5 | 190±46a | 24 | 83.3±5.3a | 6.4 | 0.41±0.10d | 25.27 |
| 10—40 | 3.8±0.2c | 4.0 | 108±26b | 24 | 72.2±5.6b | 7.7 | 0.66±0.14c | 21.66 |
| 40—70 | 4.0±0.1b | 2.2 | 73±16c | 22 | 65.6±4.3c | 6.5 | 0.89±0.13b | 14.62 |
| 70—100 | 4.0±0.1a | 1.8 | 63±23c | 35 | 63.0±5.7d | 9.0 | 0.99±0.17a | 17.28 |
0—10cm土层的SOC、MBC、EDOC和ROC含量分别为:(121.9±40.4)、(1.93±0.6)、(0.7±0.2)和(21.0±11.8) g/kg,土壤有机碳和3个碳组分含量均随土层深度的增加而降低(图 1),且0—10cm土层各碳组分含量均显著高于下面3层。其中0—10cm土层SOC含量为10—40cm、40—70cm和70—100cm 3个土层的3.6、8.6和14.6倍,0—10cm土层MBC含量也比下面3土层分别高出2.6、5.7和7.8倍。而40—70与70—100cm两土层中4个碳组分的含量均不存在显著性差异。凋落物层有机碳含量为(443±25) g/kg,且各样点测定结果差异不大,变异系数仅为6.2%。

|
| 图1 瓦屋山常绿阔叶次生林不同土层深度有机碳、微生物生物量碳、可浸提性有机碳和易氧化碳含量 Fig.1 Content of SOC、MBC、EDOC and ROC in deferent soil depth of evergreen broad-leaved secondary forest in Wawushan Mountain SOC: 土壤有机碳soil organic carbon; MBC: 微生物生物量碳microbial biomass carbon; EDOC: 可浸提溶解性有机碳extractable dissolved organic carbon; ROC: 易氧化碳readily oxidizable carbon; 图中矩形盒表示25%—75%的数据,矩形盒内横线代表该组数据的中位数,上下颈须代表该组数据中的10%—90%的数据 |
不同土层各碳组分碳密度值由各土层各碳组分含量和相应的土壤容重计算(表 3)。结果表明该森林群落土壤SOC、MBC、EDOC和ROC密度分别为7.5—48.9、0.20-0.81、0.25-0.37和0.8—8.4 kg/m3。SOC、MBC和ROC密度均随土层深度的增加而降低,且0—10cm层的密度均显著高于其下面3层,但40—70与70—100cm两土层中这3个碳组分含量差异均不显著。EDOC密度呈现先增加后降低的趋势,在10—40cm土层含量最高(0.37 kg/m3),40—70cm土层的EDOC密度高于0—10cm土层,但差异未达显著水平。SOC、MBC、ROC密度随土层深度增加而下降趋势不如其含量明显,且EDOC密度在垂直剖面上的变化规律与其含量存在差异也表明各组分碳密度明显受土层物理特性的影响。
| 土层深度 Soil depth/cm | SOC密度/(kg/m3) SOC density | MBC密度/(kg/m3) MBC density | EDOC 密度/(kg/m3) EDOC density | ROC 密度/(kg/m3) ROC density | ||||
| Mean±SD | CV/% | Mean±SD | CV/% | Mean±SD | CV/% | Mean±SD | CV/% | |
| SOC: 土壤有机碳soil organic carbon; MBC: 微生物生物量碳microbial biomass carbon; EDOC: 可浸提溶解性有机碳extractable dissolved organic carbon; ROC: 易氧化碳readily oxidizable carbon; Mean±SD: 平均值±标准差 Standard deviation; CV: 变异系数Coefficient of variation; 同列相同大写字母表示不同土层间差异不显著(单因素方差分析,LSD多重比较法,α=0.05) | ||||||||
| 0—10 | 48.9±18.5a | 37.9 | 0.8±0.3a | 40.4 | 0.27±0.08bc | 29.4 | 8.4±4.0a | 48.2 |
| 10—40 | 17.3±5.9b | 34 | 0.4±0.2b | 55.4 | 0.37±0.09a | 23.6 | 2.3±1.1b | 48.3 |
| 40—70 | 11.0±3.0c | 26.9 | 0.3±0.1bc | 50.7 | 0.29±0.09b | 29 | 1.1±0.5c | 41.5 |
| 70—100 | 7.5±2.3c | 31 | 0.2±0.1c | 60.7 | 0.25±0.08c | 33.2 | 0.8±0.4c | 52 |
土壤0—100cm层SOC储量为156 t/hm2,ROC储量(21.00 t/hm2)显著高于MBC(3.20 t/hm2)和EDOC(3.03 t/hm2),同时ROC储量占SOC储量的比例也显著高于MBC和EDOC储量占SOC含量的比例,而EDOC与MBC储量之间不存在显著差异(图 2)。凋落物层凋落物储量和凋落物层有机碳储量分别为10.4 t/hm2和4.5 t/hm2。

|
| 图2 瓦屋山常绿阔叶次生林土壤(0—100cm)各碳组分储量 Fig.2 Stock of each carbon fraction (0—100cm) in an evergreen broad-leaved secondary forest,Wawushan Mountain SOC: 土壤有机碳soil organic carbon; MBC: 微生物生物量碳microbial biomass carbon; EDOC: 可浸提溶解性有机碳extractable dissolved organic carbon; ROC: 易氧化碳readily oxidizable carbon; 不同字母表示处理间差异显著(单因素方差分析,LSD多重比较法,α=0.05) |
本研究发现,瓦屋山中山区扁刺栲-中华木荷常绿阔叶次生林土壤为酸性,且随土层深度的增加pH值升高,0—10cm土层潜在酸度(氯化钾浸提液测定)为3.2。森林土壤有机质主要由胡敏酸和富里酸等有机酸类腐殖质组成,因此土壤有机质含量较高时土壤酸度增加。田间持水量是土壤中所能保持的毛管悬着水的最大量,被认为是土壤所能稳定保持的最高含水量[22, 23]。本研究发现试验地田间持水量随土层深度的增加而降低,这与李灵等[24]对位于中亚热带湿润季风气候的武夷山0—60cm土层的研究结果相似。土壤总孔隙度和容重相互联系,反映土壤的透水性、通气性和根系生长阻力状况等[25]。本研究中土壤容重随土层深度的增加而增大,总孔隙度随土层深度的增加而减小,与李灵等[24]和王燕等[26]对亚热带常绿阔叶林地的研究结果相似。
3.2 扁刺栲-中华木荷常绿阔叶次生林土壤碳组分特征土壤有机碳水平在特定森林生态系统一定时期内处于平衡状态,主要取决于森林凋落物和植物根系状况[27, 28, 29]。目前,该次生林凋落物层有机碳储量较高(4.5 t/hm2),表层土壤腐殖质较丰富。因此,本研究中表层土壤有机碳含量较高的一个重要原因可能在于植被的快速生长带来的大量根际沉降物、死亡细根和地表凋落物输入。其另一重要原因可能与海拔有关,本试验地海拔为1600m,比处于中亚热带的浙江天童山(表 4)常绿阔叶林高出1400多m[27],年平均温度低于其他试验地,再加之该地位于华西雨屏区的中心,雨量充沛,且表层土壤较疏松,凋落物层有机碳能大量淋溶到土壤表层,但由于温度相对较低,有机碳的分解相对较慢,表层土壤有机碳的积累就较高。黄从德等[30]对四川省森林土壤(表 5)进行的大范围研究表明,处于高海拔的冷、云杉表层土壤有机碳含量是所有森林类型中最高的。
| 气候带 Climate zone | 地点 Location | 海拔 Elevation/m | 阔叶林优势树种 Dominant tree species | 林龄 Stand age/a | 有机碳含量 SOC/(g/kg) | 文献 References |
| 壳斗科 Fagaceae;樟科 Lauraceae;木荷 Schima superba;锥栗 Castanopis chinensis;厚壳桂 Cryptocarya chinesis; SOC: soil organic carbon;—: 无数据 | ||||||
| 北亚热带 Northern subtropical zone | 浙江富阳 | — | 壳斗科+樟科 | 40 | 38.0±11.8 | [31] |
| 中亚热带 Mid-subtropical zone | 浙江宁波 | 130 | 栲树+木荷 | — | 36.19 | [27] |
| 中亚热带 Mid-subtropical zone | 浙江宁波 | 180 | 栲树 | — | 57.25 | [27] |
| 中亚热带 Mid-subtropical zone | 四川瓦屋山 | 1600 | 扁刺栲、 中华木荷 | 56 | 121.90±40.4 | 本研究 |
| 南亚热带 Southern subtropical zone | 广东鼎湖山 | 50—200 | 锥栗、木荷、 厚壳桂 | 400 | 37.2 ± 7.3 | [32] |
| 林分类型 Type of stand | 凋落物层有机碳储量 Carbon stock in litter layer/(t/hm2) | 0—10cm土层有机碳含量 Concentration of SOC in 0—10cm soil/(g/kg) | 0—100cm土层有机碳储量 Carbon stock in 0—100cm soil/(t/hm2) | 参考文献 References |
| 冷、云杉 Abies fabri and Picea asperata | 6.70±2.80 | 72.4±25.5 | 264.4±49.2 | [30, 33] |
| 杉木 Cunninghamia lanceolata | 0.71±0.22 | 24.0±6.3 | 126.8±18.5 | [30, 33] |
| 马尾松 Pinusmassoniana | 2.00±0.61 | 18.7±4.4 | 102.7±21.1 | [30, 33] |
| 柏木 Cupressus funebris | 0.87±0.31 | 19.2±7.0 | 107.6±22.8 | [30, 33] |
| 柳杉 Cryptomeria fortunei | 0.56±0.03 | 55.6±7.4 | 220.5±34.5 | [30, 33] |
| 楠、樟 Phoebe and Cinnamomum | 1.79±0.54 | 51.5±18.0 | 186.8±60.2 | [30, 33] |
| 栎类、硬阔 Quercus and Hardwood | 1.26±0.38 | 28.6±1.0 | 156.4±28.5 | [30, 33] |
| 软阔 Softwood | 2.20±0.17 | 62.3±25.1 | 240.0±46.2 | [30, 33] |
| 栎类、硬阔 Quercus and Hardwood | 10.03±3.1 | 121.9±40.4 | 156.1±32.8 | 本研究 |
土壤微生物生物量碳在土壤碳库中所占比例很小,一般只占有机碳的1%—4%[34],但它与全量养分存在一定的相关性,其含量越高,其土壤肥力往往越高,因此,被视为土壤肥力变化的重要指标之一[35, 36]。本研究结果0—10cm土层MBC含量为1931.82mg/kg,高于刘爽等[37]报道的温带硬阔叶林(1302mg/kg),杨桦林(1074mg/kg)和蒙古栎林(721mg/kg);也高于李灵等[38]报道的福建三明格氏栲人工林[(1035±103)mg/kg]和杉木人工林[(820±76)mg/kg]。本研究结果高于温带硬阔林和栎林的一个重要原因可能是本研究地所处中亚热带,水热条件均优于黑龙江帽儿山;而高于同一气候带且林分年龄差不多(约50a)的栲和杉木人工林的重要原因可能是人工林地中树种过于单一,使得微生物群落不如本研究地的丰富多样。本研究中,土壤MBC含量随土层的加深而降低(图 1),这与尉海东等[39]对杨树人工林的研究,李灵等[38]对格氏栲和杉木人工林的研究和刘爽等[37]对温带森林的研究结果一致。土壤微生物多分布在表层土中[39],原因有两点,一方面,地表大量的枯枝落叶提供充分的营养源,通气和水热状况也较好,利于微生物的生长和繁殖;另一方面,表层土壤中细根相对密集,土壤有机质也较多、土壤通气状况较好,但随着土层的加深,生境条件变差,从而影响微生物的分布。
土壤可浸提溶解性碳是有机碳中具有一定溶解性、易分解、移动快、可以被土壤微生物直接利用的有机碳源[40, 41, 42],对森林土壤生态系统中元素的生物地球化学循环及铝、重金属、水和养分循环及有效性等影响较大[43]。李淑芬等[44]的研究结果表明,南方酸性森林土壤中溶解性有机碳与一些全量和速效养分、有机络合态铁和有机络合态铝之间呈极显著或显著的正相关关系,因此可以作为土壤肥力性状及调整土壤中铝和铁的溶出和迁移生物学指标。本研究中0—10cm土层EDOC含量697.42mg/kg,0—100cm土层储量为302.59 g/cm2,均远远大于福建省万木林省级自然保护区罗浮栲天然林土壤表层可溶性有机碳含量(74.84mg/kg)与0—100cm土层储量(28.40 g/cm2)[45]。这表明本研究地由土壤总有机碳转化的可溶性有机碳较为丰富,为整个扁刺栲-中华木荷次生林养分元素循环和微生物分布提供保证,该次生林的整个养分流动较为活跃。
土壤易氧化碳含量及其与土壤总有机碳的比值反映了土壤碳的稳定性,土壤总有机碳中ROC所占比例越高,说明土壤碳的活性越大,稳定性越差[35],同时作为有机碳中的活性组分,ROC直接参与土壤生物化学转化过程,为微生物活动提供能源并驱动土壤养分循环,对土壤碳及其质量变化具有更高的灵敏性[46],因此,ROC对土壤碳库平衡和土壤化学、生物学稳定性维持具有重要的意义,土壤ROC多少往往表征着土壤能释放的有效肥力的多少[47]。本研究中0—10cm土层ROC含量为20.98 g/kg,占SOC含量的17.21%,0—100cm层ROC储量占SOC储量的比例也达到13.50%,高于江苏省瓦屋山林场杉木、毛竹、麻栎和湿地松人林0—10cm层ROC含量[48];也高于浙江山坞壳斗科樟科为主的常绿阔叶天然次生林[(15.58±3.15) g/kg)和杉木人工林\[(13.30±1.20) g/kg][49]。黄宗胜等[47]在贵州茂兰所做的关于喀斯特森林植被自然恢复过程中土壤有机碳库特征演化结果表明ROC含量在早期阶段(草本和草灌阶段)最低,中期阶段(灌丛灌木和灌乔过渡阶段)次之,后期阶段(乔木和顶极阶段)最高。本研究地处于该区域植被恢复的后期阶段,ROC含量也高于大多人工林和天然次生林地,表明本研究地土壤能为微生物生存提供充足能源,凋落物层和土壤中有效肥力释放的潜力较大。
试验地0—100cm土壤SOC、MBC、EDOC和ROC储量分别为154.9、3.1、3.0、21.0 t/hm2,其中土壤有机碳储量在四川省各类森林中处于中等水平,高于四川省杉木(Cunninghamia lanceolata)、云南松(Pinus yunnanensis)、马尾松(Pinusmassoniana)等部分天然林和桉(Eucalyptus)、柏木(Cupressus funebris)、杨(Populus)等大部分人工林[29, 30],且高于我国亚热带地区大部分人工林(71.5—147.0 t/hm2)[50, 51, 52]与四川西北部亚高山云杉天然林土壤有机碳储量(156.1 t/hm2)[53]非常接近(表 5,表 6)。瓦屋山常绿阔叶次生林有机碳密度和储量较高的一个重要原因可能是表层中有机碳含量非常高。黄从德等[30]研究的四川省各类森林中,0—10cm层SOC含量介于19.2—72.4 g/kg,均明显小于本试验地该土层SOC含量(121.9 g/kg),而本研究中10—100cm各土层SOC含量与四川省其他各类森林差异不大。
综上所述,位于华西雨屏区中心地带的瓦屋山扁刺栲-中华木荷常绿阔叶次生林,土壤有机碳储量高于同一气候带人工林和天然林,在四川省各类森林中处于中等水平,而凋落物层有机碳储量和土壤表层有机碳含量均明显高于四川省各类人工林和天然林,微生物生物量碳、可浸提溶解性有机碳和易氧化碳高于许多已报道的天然次生林和人工林,说明该生态系统活性碳含量较大,微生物活动和养分流动较为活跃,凋落物转化为土壤碳的潜力较大,碳储量仍处在不断积累的过程。因此可以预见这类生态系统将在区域碳循环过程中扮演更为重要的角色。
| [1] | Chokkalingam U, Jong W D. Secondary forest: a working definition and typology. International Forestry Review, 2001, 3(1): 19-26. |
| [2] | Guariguata M R, Ostertag R. Neotropical secondary forest succession: changes in structural and functional characteristics. Forest Ecology Management, 2001, 148(1/3): 185-206. |
| [3] | FAO. 2011. Global forest resource assessment 2010. United Nations Food and Agriculture Organization (UN FAO), Rome, Italy. |
| [4] | Shevliakova E, Pacala SW, Malyshev S, Jurtt C C, Milly P C D, Caspersen J P, Sentman L T, Fisk J P, Wirth C, Crevoisier C. Carbon cycling under 300 years of land use change: Importance of the secondary vegetation sink. Global Biogeochemical Cycles, 2009, 23(2), doi: 10.1029/2007 GB003176. |
| [5] | Yang X, Richardson T K, Jain A K. Contributions of secondary forest and nitrogen dynamics to terrestrial carbon uptake. Biogeosciences, 2010, 7(10): 3041-3050. |
| [6] | Xiang W H, Liu S H, Lei X D, Frank S C, Tian D L, Wang G J, Deng X W. Secondary forest floristic composition, structure, and spatial pattern in subtropical China. Journal of Forest Research, 2013, 18(1): 111-120. |
| [7] | LAL R. Soil Carbon Sequestration Impacts on Global Climate Change and Food Security. Science, 2004, 304(5677): 1623-1627. |
| [8] | Tarnocai C, Canadell J G, Schuur E A G, Kuhry P, Mazhitova G, Zimov S. Soil organic carbon pools in the northern circumpolar permafrost region. Global Biogeochemical Cycles, 2009, 23(2), doi: 10.1029/2008GB003327. |
| [9] | Nair P K R, Nair V D, Kumar B M, Haile S G. Soil carbon sequestration in tropical agroforestry systems: a feasibility appraisal. Environmental Science and Policy, 2009, 12(8): 1099-1011. |
| [10] | Mishra U, Ussiri D A D, Lal R. Tillage effects on soil organic carbon storage and dynamics in Corn Belt of Ohio USA. Soil and Tillage Research, 2010, 107(2): 88-96. |
| [11] | Xi X H, Yang Z F, Cui Y J, Sun S M, Yu C G, Li M. A study of soil organic carbon distribution and storage in the Northeast Plain of China. Geoscience Frontiers, 2011, 2(2): 115-123. |
| [12] | Li M M, Zhang X C, Pang G W, Han F P. The estimation of soil organic carbon distribution and storage in a small catchment area of the Loess Plateau. Catena, 2013, 101: 11-16. |
| [13] | 王绍强, 周成虎, 李克让, 朱松丽, 黄方红. 中国土壤有机碳库及空间分布特征分析. 地理学报, 2000, 55(5): 533-544. |
| [14] | Blair G J, Rod D B L, Leanne L. Soil carbon fractions based on their degree of oxidation, and the development of a carbon management index for agricultural systems. Australian Journal of Agricultural Research, 1995, 46(7): 1459-1466. |
| [15] | Biederbeck V O, Janzen H H, Campbell C A, Zentner R P. Labile soil organic matter as influenced by cropping practices in an arid environment. Soil Biology and Biochemistry, 1994, 26(12): 1647-1656. |
| [16] | Whitbread A M, Lefroy R D B, Blair G J. A survey of the impact of cropping on soil physical and chemical properties in north-western New South Wales. Australian Journal of Soil Research, 1998, 36(4): 669-682. |
| [17] | 庄平, 高贤明. 华西雨屏带及其对我国生物多样性保育的意义. 生物多样性, 2002, 10(3): 339-344. |
| [18] | 包维楷, 刘照光, 刘朝禄, 袁亚夫, 刘仁东. 中亚热带湿性常绿阔叶次生林自然恢复15年来群落乔木层的动态变化. 植物生态学报, 2000, 24(6): 702-709. |
| [19] | 包维楷, 刘照光, 刘朝禄, 袁亚夫, 刘仁东. 亚热带次生常绿阔叶林主要乔木种群自然恢复15年来的变化. 林业科学, 2001, 37(1): 7-14. |
| [20] | Wu J, Joergensen R G, Pommerening B, Chaussod R, Brookes P C. Measurement of soil microbial biomass C by fumigation-extraction-an automated procedure. Soil Biology and Biochemistry, 1990, 22(8): 1167-1169. |
| [21] | Murphy D V, Macdonald A J, Stockdale E A, Goulding K W T, Fortune S, Gaunt J L, Poulton P R, Wakefield J A, Webster C P, Wilmer W S. Soluble organic nitrogen in agricultural soils. Biology and Fertility of Soils, 2000, 30(5/6): 374-387. |
| [22] | 杨绍锷, 吴炳方, 闫娜娜. 基于AMSR-E数据估测华北平原及东北地区土壤田间持水量. 土壤通报, 2012, 43(2): 301-305. |
| [23] | 王丹丹, 郑纪勇, 颜永毫, 张兴昌. 生物炭对宁南山区土壤持水性能影响的定位研究. 水土保持学报, 2013, 27(2): 101-109. |
| [24] | 李灵, 张玉, 孔丽娜, 江慧华, 陈家晖, 周艳. 武夷山风景区不同林地类型土壤水分物理性质及土壤水库特性. 水土保持通报, 2011, 31(3): 60-65. |
| [25] | 庞学勇, 刘世全, 刘庆, 吴彦, 林波, 何海, 张宗锦. 川西亚高山针叶林植物群落演替对土壤性质的影响. 水土保持学报, 2003, 17(4): 42-45, 50-50. |
| [26] | 王燕, 王兵, 赵广东, 郭浩, 马向前, 邓宗富. 江西大岗山3种林型土壤水分物理性质研究. 水土保持学报, 2008, 22(1): 151-153. |
| [27] | 张庆费, 由文辉, 宋永昌. 浙江天童植物群落演替对土壤化学性质的影响. 应用生态学报, 1999, 10(1): 19-22. |
| [28] | Cui X Y, Wang Y F, Niu H S, Wu J, Wang S P, Schnug E, Rogasik J, Fleckenstein J, Tang Y H. Effect of long-term grazing on soil organic carbon content in semiarid steppes in Inner Mongolia. Ecological Research, 2005, 20(5): 519-527. |
| [29] | 马秀枝, 王艳芬, 汪诗平, 王金枝, 李长生. 放牧对内蒙古锡林河流域草原土壤碳组分的影响. 植物生态学报, 2005, 29(4): 569-576. |
| [30] | 黄从德, 张健, 杨万勤, 张国庆, 王永军. 四川森林土壤有机碳储量的空间分布特征. 生态学报, 2009, 29(3): 1217-1225. |
| [31] | 马少杰, 李正才, 周本智. 北亚热带天然次生林群落演替对土壤有机碳的影响. 林学科学研究, 2010, 23(6): 845-849. |
| [32] | 欧阳学军, 周国逸, 魏识广, 黄忠良, 李炯, 张德强. 南亚热带森林植被恢复演替序列的土壤有机碳氮矿化. 应用生态学报, 2007, 18(8): 1688-1694. |
| [33] | 黄从德, 张健, 杨万勤, 张国庆. 四川人工林生态系统碳储量特征. 应用生态学报, 2008, 19(8): 1644-1650. |
| [34] | Jenkinson D S, Ladd J N. Microbial biomass in soil, measurement and turnover. Soil Biochemistry, 1981, 5: 415-471. |
| [35] | 朱志建, 姜培坤, 徐秋芳. 不同森林植被下土壤微生物量碳和易氧化态碳的比较. 林业科学研究, 2006, 19(4): 523-526. |
| [36] | Powlson D S, Jenkinson D S. A comparison of the organic matter, biomass, adenosine triphosphate and mineralizable nitrogen contents of ploughed and direct-drilled soils. Journal of Agricultural Science, 1981, 97(3): 713-721. |
| [37] | 刘爽, 王传宽. 五种温带森林土壤微生物微生物量碳氮的时空格局. 生态学报, 2010, 30(12): 3135-3143. |
| [38] | 李灵, 张玉, 王利宝, 王丽梅. 不同林地土壤微生物生物量垂直分布及相关性分析. 中南林业科技大学学报, 2007, 27(2): 52-56, 60-60. |
| [39] | 尉海东, 董彬. 鲁东南杨树人工林土壤微生物生物量碳的时空动态. 生态环境学报, 2013, 22(2): 233-238. |
| [40] | 刘淑霞, 刘景双, 赵明东, 王金达, 于君宝, 丛晓明. 土壤活性有机碳与养分有效性及作物产量的关系. 吉林农业大学学报, 2003, 25(5): 539-543. |
| [41] | 李忠佩, 张桃林, 陈碧云. 可溶性有机碳的含量动态及其与土壤有机碳矿化的关系研究. 土壤学报, 2004, 41(4): 544-552. |
| [42] | Liang B C, MacKenzie A F, Schnitzer M, Monreal C M, Voroney P R, Beyaert R P. Management-induced change in labile soil organic matter under continuous corn in eastern Canadian soils. Biology and Fertility of Soils, 1997, 26(2): 88-94. |
| [43] | Zsolnay A. Dissolved organic matter: artifacts, definitions, and functions. Geoderma, 2003, 113(3/4): 187-209. |
| [44] | 李淑芬, 俞元春, 何晟. 南方森林土壤溶解有机碳与土壤因子的关系. 浙江林学院学报, 2003, 20(2): 119-123. |
| [45] | 汪伟, 杨玉盛, 陈光水, 郭剑芬, 钱伟. 罗浮栲天然林土壤可溶性有机碳的剖面分布及季节变化. 生态学杂志, 2008, 27(6): 924-928. |
| [46] | 杨丽霞, 潘剑君. 土壤活性有机碳库测定方法研究进展. 土壤通报, 2004, 35(4): 502-506. |
| [47] | 黄宗胜, 符裕红, 喻理飞. 喀斯特森林植被自然恢复过程中土壤有机碳库特征演化. 土壤学报, 2013, 50(2): 306-314. |
| [48] | 李平, 王国兵, 郑阿宝, 沈玉娟, 赵琦齐, 王琳飞, 蒋如生, 李莉, 阮宏华. 苏南丘陵区4种典型人工林土壤活性有机碳分布特征. 南京林业大学学报: 自然科学版, 2012, 36(4): 79-83. |
| [49] | 刘荣杰, 吴亚丛, 张英, 李正才, 马少杰, 王斌, 格日乐图. 中国北亚热带天然次生林与杉木人工林土壤活性有机碳库的比较. 植物生态学报, 2012, 36(5): 431-437. |
| [50] | 雷丕锋, 项文化, 田大伦, 方晰. 樟树人工林生态系统碳素贮量与分布研究. 生态学杂志, 2004, 23(4): 25-30. |
| [51] | 李跃林, 胡成志, 张云, 文锦柱. 几种人工林土壤碳储量研究. 福建林业科技, 2004, 31(4): 4-7. |
| [52] | 杨玉盛, 谢锦升, 盛浩, 陈光水, 李旭. 中亚热带山区土地利用变化对土壤有机碳储量和质量的影响. 地理学报, 2007, 62(11): 1123-1131. |
| [53] | 马东明, 江洪, 罗承德, 刘跃建. 四川西北部亚高山云杉天然林生态系统碳密度、净生产量和碳贮量的初步研究. 植物生态学报, 2007, 31(2): 305-312. |