 2015, Vol. 35
2015, Vol. 35文章信息
- 马文瑛, 赵传燕, 彭守璋, 高云飞, 袁亚鹏, 李文娟
- MA Wenying, ZHAO Chuanyan, PENG Shouzhang, GAO Yunfei, YUAN Yapeng, LI Wenjuan
- 黑河天涝池五种植被类型土壤呼吸速率动态特征及其影响因子
- Variation in soil respiration rate and factors affecting it in five vegetation types in Tianlaochi catchment in Heihe River
- 生态学报, 2015, 35(17): 5654-5665
- Acta Ecologica Sinica, 2015, 35(17): 5654-5665
- http://dx.doi.org/10.5846/stxb201311102706
-
文章历史
- 收稿日期: 2013-11-10
- 网络出版日期: 2014-11-03






随着人类活动和全球变化的加剧,大气中CO2等温室气体的浓度呈现持续上升的趋势,受其影响预测 2100年全球平均气温将升高 1.1—6.4 ℃[1, 2]。土壤呼吸是大气CO2的重要排放源,也是全球陆地生态系统碳循环的重要组成部分,在区域和全球尺度上对大气CO2浓度调控起着十分关键的作用[3]。据估计,全球陆地生态系统由土壤呼吸每年释放到大气中的碳通量高达 80.4 Pg C/a,占全球碳通量的 10%,是化石燃料燃烧释放CO2的 13倍,仅次于全球陆地总初级生产力的估算值 100—120 Pg C/a,因此,其任何一丝微小变化都会显著改变大气中CO2的浓度和土壤碳的累积速率[4]。高寒区域是环境变化的敏感区,但是对该区域土壤呼吸理解非常有限,尤其是对土壤呼吸的影响因素及生态系统间土壤呼吸变异性的认识缺乏[5, 6, 7]。不同植被类型对碳排放的贡献多大?影响高寒区域土壤呼吸的关键因素是什么?这些问题是研究碳循环必须回答的科学问题。
土壤呼吸是一个复杂的生物化学过程,它不仅受到土壤温度、土壤含水量、降水、凋落物以及土壤C、N含量等非生物因子的影响,而且受到植被类型、叶面积指数 (LAI)、根系生物量等生物因子和人类活动的综合影响[8, 9]。研究表明土壤温度和湿度能解释土壤呼吸的大部分变异[10]。在高寒山区研究报道,土壤温度均与土壤呼吸速率呈显著的指数关系,温度对土壤呼吸的影响在低温时比高温时更显著;当土壤水分含量较低的情况下,随着土壤水分含量的增加,土壤呼吸速率也随着增加,但是当土壤水分含量增加到一定程度时,土壤呼吸速率则表现出降低的趋势[11]。水热组合在海拔梯度上的变化导致土壤呼吸的差异,常宗强在生长季节对祁连山高山草甸土壤CO2通量沿海拔梯度进行了野外定位观测,统计分析得到土壤CO2通量随着海拔梯度的增加而逐渐减小[12]。植被类型对土壤呼吸作用的影响也不容忽视,因为植被状况的不同除直接影响土壤呼吸外,又影响土壤温度和湿度而间接作用土壤呼吸[13],如不同的植被盖度通过改变辐射特征、植物的蒸腾作用等影响土壤温度和水分等环境要素[14],正如Raich等认为植物群落间土壤呼吸的差异原于不同植物群落导致不同土壤微气候环境[13]。陈全胜等[15]指出研究土壤呼吸对温度等变化的响应需按照不同植被状况进行。在同一区域对不同植被类型下土壤的呼吸研究,不但能够精确估算区域土壤碳排放,而且能够准确理解不同植被类型作为碳源或碳汇的作用,此外还有助于了解地下碳分配过程和生态系统的生产力[16]。
黑河上游位于祁连山区的中部,属于高寒山地气候,植被类型呈明显的垂直地带性分布。目前,祁连山区土壤呼吸的研究资料十分缺乏,有关不同植被类型土壤呼吸差异的研究更是鲜见。本研究采用 LI-8100土壤碳通量测量系统,对黑河上游天涝池流域的5种典型植被类型(干草原、亚高山草原、亚高山灌丛、祁连圆柏林和青海云杉林)的土壤呼吸速率及其影响因子进行观测,旨在探讨不同植被类型土壤呼吸的动态变化规律,分析温度和湿度对土壤呼吸速率的影响,建立土壤呼吸速率与温度和湿度的关系模型,以期深入了解土壤呼吸作用的变化过程及变化机理,试图理解山地土壤碳排放机理,为山地森林草地生态系统碳循环研究奠定基础,对合理评价不同植被类型的碳源或碳汇提供科学依据。
1 材料与方法 1.1 研究区概况试验区位于祁连山国家级自然保护区天涝池小流域 (38°20′—38°30 ′N,99°44′—99°59 ′E),海拔 2600—4450 m,面积 12.8 km2。流域气候类型属于高寒半干旱、半湿润山地森林草原气候,无霜期 90—120 d,年均气温 -0.6—2.0 ℃,最高温度为 28 ℃,最低气温为 -36 ℃,年降水量 400—500 mm,降水主要集中在 5—9月份,占全年降水量的 89.2%,潜在蒸发量 1051.7 mm,年日照时数 1892.6 h,年平均相对湿度为 60%。流域内降雨量随海拔升高明显增加,气温显著降低,水热的组合随海拔的不同而异,其植被类型呈现明显的垂直梯度分布。在低海拔的阳坡 (2800 m以下),分布着小面积的干草原,紫花针茅(Stipa purpurea)和冰草(Agropyron cristatum)为优势种;2700—3100 m 较平坦的区域分布着一定面积的亚高山草原,由狐茅(Festuca ovina)、针茅(Stipa capillata)、苔草(Carex tristachya)组成;2700—3300 m 阴坡为青海云杉林,阳坡为祁连圆柏林;3300—3800m 为亚高山灌丛,其中金露梅(Potentilla fruticosa)、鬼箭锦鸡儿(Caragana jubata)和吉拉柳(Salix gilashanica)为优势种;3800 m 以上为稀疏垫状植被、裸岩和冰雪。土壤类型主要为山地灰褐土和亚高山灌丛草甸土,在干草原区分布着小面积的山地灰钙土。研究区 2013年禁牧,观测过程中受人类干扰影响较小。各实验样地的地形特征与植被、土壤状况见表 1。
| 植被类型 Vegetation type | 海拔 Altitude/m | 坡向 Aspect | 坡度 Slope/(°) | 主要植物 Main species composition | 土壤类型 Soil type |
| 干草原 Stipa purpurea steppe | 2715 | 阳坡 | 29—34 | 芨芨草Achnatherum splendens,苔草Carex tristachya,紫花针茅Stipa purpurea,冰草Agropyron cristatum,狼毒Euphorbia fischeriana Steud | 山地灰钙土 |
| 亚高山草原 Subalpine steppe | 3068 | 阳坡 | 6—9 | 苔草Carex tristachya,车前草Plantago asiatica,委陵菜Potentilla,小茴香Foeniculumvulgare | 山地灰褐土 |
| 亚高山灌丛 Subalpine shrub | 3145 | 阳坡 | 18—23 | 金露梅 Potentilla fruticosa,吉拉柳Salix gilashanica,鬼箭锦鸡Caragana jubata,芨芨草 Achnatherum splendens | 亚高山灌丛 草甸土 |
| 青海云杉林 Picea crassifolia forest | 3101 | 阴坡 | 17—23 | 青海云杉picea crassifolia,苔藓Bryophyta,苔草Carex tristachya,珠芽蓼 Polygonaceae | 山地灰褐土 |
| 祁连圆柏林 Sabina przewalskii forest | 3200 | 阳坡 | 24—32 | 祁连圆柏Sabina przewalskii,金露梅Potentilla fruticosa,苔草Carex tristachya,珠芽蓼 Polygonaceae | 山地灰褐土 |
2013 年5 月初,在干草原、亚高山草原、亚高山灌丛林内设置 5 m×5 m 样地,青海云杉林和祁连圆柏林内设置 10 m×10 m 样地,在样地内随机布设 3 个土壤呼吸基座(直径 20 cm,高 12 cm),埋入土壤 10 cm 左右,在整个生长季的观测过程中土壤呼吸基座位置不变。从 2013年 5 月下旬开始,每隔 10 d,采用LI-COR公司生产的Li-8100土壤碳通量自动测量系统(LI-COR,Lincoln,NE,USA)对土壤呼吸进行测定,一直持续到 9 月上旬,共有效观测次数为 9 次,所有样地每次所用时间为 2 d。每个样地测定时间为 8:00—19:00,测量步长为 1 h,若无降雨干扰共测 12 次。在每次测定前 1 d,将土壤基座内的地表植被剪除,并尽量不破坏土壤,以减少因土壤扰动及根系损伤对测量结果的影响。
在测定土壤呼吸的同时,用Li-8100自带的温度和湿度探头测定 10 cm 土壤温度(Ts)、5 cm土壤湿度(RHs)和地表温度(Tas)。同时,每个样地都配有自动气象站(HOBO U30 Station),测量的要素包括:地下 0—20、20—40、40—60 cm 的土壤温度和含水量,光合有效辐射,风速,空气相对湿度,大气温度等环境因子,记录时间步长为 10 min。
1.3 数据处理与分析以同一植被类型为基本单位,将 3 个重复样观测的所有要素进行平均,获得小时的平均值,用于分析不同植被类型下的土壤呼吸及其环境要素的日变化。将小时数据进行平均,获得日平均值,将每个月的日平均值再次平均获得月平均值,用于分析不同植被类型下的土壤呼吸及其环境要素的月变化。并比较5种植被类型土壤呼吸速率日变化和月变化差异。
采用如下指数模型[17]来拟合生长季土壤呼吸速率和温度间的关系:
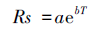
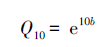
土壤呼吸速率与湿度关系采用线性模型[20]:
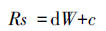
由于影响因子之间存在一定的相关性,如何找出几个综合因子来代替原来众多的因子,使这些综合因子尽可能地代表原来因子的信息量,而且彼此之间互不相关,所以需要进行主成分分析[21]。对影响土壤呼吸速率的环境因子数据,如土壤温度、土壤湿度、空气温度、空气相对湿度、风速等输入SPSS进行主成分分析,具体为Analysis→Data Reduction→Factor,选择Principal Components,通过主成分分析得出影响土壤呼吸速率的综合因子。根据主成分的累积贡献率来决定主成分个数,一般要求累计贡献率达到 85% 以上[21],这样才能保证综合变量能包括原始变量的绝大多数信息。最后将主成分分析得到的综合因子与土壤呼吸速率进行回归分析。
2 结果与分析 2.1 土壤呼吸速率以及环境因子的日变化动态不同植被类型土壤呼吸速率及环境因子的日变化曲线存在差异,日变化幅度大小因植被类型而异(图 1)。5种植被类型中,以亚高山草原土壤呼吸速率日变幅最大 (3.81—8.38 μmol m-2 s-1),以青海云杉林土壤呼吸速率日变幅最小 (1.87—2.35 μmol m-2 s-1)。5种植被类型土壤呼吸速率的日变化规律基本一致,表现为单峰型曲线。除祁连圆柏林土壤呼吸速率在 13:00 时达到最大值之外,其余 4 种植被类型均在 14:00 时达到最大值。5种植被类型生长季平均土壤呼吸速率存在差异(见表 2),各植被类型土壤呼吸速率大小依次为亚高山草原(6.38 μmol m-2 s-1 )>干草原(4.67 μmol m-2 s-1 )>亚高山灌丛 (3.45 μmol m-2 s-1 )>祁连圆柏林 (2.43 μmol m-2 s-1 )>青海云杉林 (2.16 μmol m-2 s-1 )。

|
| 图1 土壤呼吸速率及环境因子的日动态 Fig.1 Diurnal variation of soil respiration rate and environmental Factors in five vegetation types |
地表温度在 14:00 时达到最大,土壤湿度随时间增加先减小后增加的趋势,在 17:00时最小,此时土壤温度升高到了最大值(图 1)。5种植被类型之间,生长季平均土壤湿度 (5 cm)、土壤温度 (10 cm)和地表温度因植被类型而异。其中,干草原由于海拔低平均土壤温度 (13.28 ℃) 和地表温度 (18.53 ℃)最高,而青海云杉林的平均土壤温度 (6.26 ℃)、地表温度 (13.46 ℃) 和土壤湿度 (64.29%) 均表现为最小。
| 植被类型 Vegetation type | 土壤呼吸速率 Soil respiration rate/ (μmol m-2 s-1) | 10cm土壤温度 Soil temperature/℃ | 地表温度 Surface temperature/℃ | 5cm土壤湿度 Soil moisture/% | ||||
| 均值Mean | 标准误SE | 均值Mean | 标准误SE | 均值Mean | 标准误SE | 均值Mean | 标准误SE | |
| 亚高山草原 Subalpine steppe | 6.38 | 0.30 | 11.00 | 0.32 | 17.70 | 0.73 | 71.20 | 1.87 |
| 干草原 Stipa purpurea steppe | 4.67 | 0.21 | 13.28 | 0.39 | 18.53 | 0.85 | 66.02 | 2.25 |
| 亚高山灌丛 Subalpine shrub | 3.45 | 0.21 | 8.30 | 0.34 | 15.99 | 0.99 | 75.51 | 0.98 |
| 祁连圆柏林 Sabina przewalskii forest | 2.43 | 0.58 | 6.74 | 0.36 | 14.20 | 0.57 | 66.90 | 1.36 |
| 青海云杉林 Picea crassifolia forest | 2.16 | 0.87 | 6.26 | 0.31 | 13.46 | 0.82 | 64.29 | 1.98 |
在生长季,土壤呼吸速率呈现明显的月变化(表 3)。表现为随时间推移先增加,在 7 月均达到最大值,之后降低,且不同植被类型土壤呼吸速率变化范围各不相同。5 月份干草原土壤呼吸速率高于其它植被类型,表现为干草原>亚高山草原>亚高山灌丛>青海云杉林>祁连圆柏林,6—9 月份土壤呼吸速率均表现为亚高山草原>干草原>亚高山灌丛>祁连圆柏林>青海云杉林。由图 2可以看出,5 月份的干草原其地表温度和土壤温度均显著高于其它类型,水热良好的组合使干草原植被发芽和生长早于其他植被类型,这就导致其土壤呼吸速率在该月最高。各植被类型生长季土壤呼吸速率变化范围为:亚高山草原 2.01—12.10 μmol m-2 s-1;干草原 1.40—8.45 μmol m-2 s-1;亚高山灌丛 0.85—9.24 μmol m-2 s-1;祁连圆柏林 1.09—4.14 μmol m-2 s-1;青海云杉林 1.25—3.19 μmol m-2 s-1。其中亚高山灌丛土壤呼吸速率的变化幅度最大,青海云杉林最小,最大土壤呼吸速率与最小土壤呼吸速率的倍数在 2.6—10.9 之间。
| 植被类型Vegetation type | 土壤呼吸速率Soil respiration rate/(μmol m-2 s-1) | ||||||
| 5月May | 6月June | 7月July | 8月August | 9月September | |||
| 干草原Stipa purpurea steppe | 4.27 | 4.13 | 5.54 | 4.22 | 3.55 | ||
| 亚高山草原Subalpine steppe | 4.17 | 4.70 | 8.20 | 7.55 | 6.39 | ||
| 亚高山灌丛Subalpine shrub | 2.06 | 2.91 | 4.09 | 3.85 | 2.46 | ||
| 青海云杉林Picea crassifolia forest | 1.67 | 2.05 | 2.58 | 2.36 | 1.89 | ||
| 祁连圆柏林Sabina przewalskii forest | 1.36 | 2.12 | 3.18 | 2.62 | 2.53 | ||

|
| 图2 土壤呼吸速率及环境因子的月变化动态 Fig.2 Monthly variation of soil respiration rate and environmental Factors in five vegetation types |
在所有植被类型中,土壤呼吸速率与 10 cm 深土壤温度、地表温度和大气温度均存在显著的指数相关 (P<0.05),10 cm 土壤温度可以解释各植被类型土壤呼吸速率生长季变化的 23—47%,地表温度可以解释各植被类型土壤呼吸速率生长季变化的 79%—95%,大气温度可以解释各植被类型土壤呼吸速率生长季变化的 50%—77% (表 4)。土壤呼吸速率对地表温度和大气温度的响应高于土壤温度,说明研究区土壤呼吸速率有随空气温度变化而变化的趋势。青海云杉林土壤呼吸速率与土壤温度的相关性高于其他植被类型,说明青海云杉林土壤呼吸速率对土壤温度变化的敏感性较大。5种植被类型的Q10值模拟结果整体表现为亚高山灌丛 (2.77) >干草原 (2.22) >亚高山草原 (1.95) >祁连圆柏林 (1.61) >青海云杉林 (1.31),也就是说升高相同的温度,亚高山灌丛将会放出更多的CO2,这也可以作为不同植被类型生长季变化幅度差异的原因之一。
| 植被类型 Vegetation type | 10 cm土壤温度/℃* Soil temperature | 地表温度/℃ Surface temperature | 大气温度/℃ Air temperature |
| 干草原 Stipa purpurea steppe | Rs=1.60e0.08T,R2=0.31 Q10=2.23 | Rs=1.59e0.06T,R2=0.95 Q10=1.82 | Rs=2.07e0.06T,R2=0.77 Q10=1.82 |
| 亚高山草原 Subalpine steppe | Rs=1.80e0.11T,R2=0.46 Q10=3.00 | Rs=2.39e0.05T,R2=0.82 Q10=1.65 | Rs=2.41e0.07T,R2=0.53 Q10=2.01 |
| 亚高山灌丛 Subalpine shrub | Rs=0.98e0.14T,R2=0.29 Q10=4.06 | Rs=1.04e0.07T,R2=0.79 Q10=2.01 | Rs=0.42e0.08T,R2=0.63 Q10=2.23 |
| 青海云杉林 Pcea crassifolia forest | Rs=1.64e0.04T,R2=0.47 Q10=1.49 | Rs=1.61e0.02T,R2=0.89 Q10=1.22 | Rs=1.70e0.02T R2=0.75 Q10=1.22 |
| 祁连圆柏林 Sabina przewalskii forest | Rs=1.75e0.05T,R2=0.23 Q10=1.65 | Rs=1.67e0.03T,R2=0.87 Q10=1.35 | Rs=1.73e0.06T,R2=0.50 Q10=1.82 |
| P值均<0.05 | |||
由上可以看出土壤呼吸速率与温度变化呈一定的相关关系,而影响土壤呼吸速率的因素不仅仅是温度,所以本研究对土壤湿度、大气相对湿度和风速与土壤呼吸速率进行回归分析,结果见表 5。从表可以看出,5种植被类型土壤呼吸速率与土壤湿度和大气相对湿度均呈显著负相关 (P<0.05),相关系数各有差异。与风速呈显著正相关 (P<0.05),相关性大小表现为干草原>亚高山草原>祁连圆柏林>青海云杉林>亚高山灌丛,由此可以看出风速对土壤呼吸速率的影响可能取决于土壤呼吸基座周围的植被灌丛高度和结构。
| 植被类型 Vegetation type | 土壤湿度 Soil moisture/% | 大气相对湿度 Atmospheric humidity/% | 风速 Wind speed /(m/s) |
| 干草原 Stipa purpurea steppe | Rs=-7.27w+99.86,R2=0.82 | Rs=-10.49w+105.05,R2=0.89 | Rs=0.43ws-0.61,R2=0.73 |
| 亚高山草原 Subalpine steppe | Rs=-1.61w+81.44,R2=0.67 | Rs=-6.37w+92.37,R2=0.93 | Rs=0.58ws+0.64,R2=0.62 |
| 亚高山灌丛 Subalpine shrub | Rs=-1.47w+74.40,R2=0.74 | Rs=-3.89w+62.91,R2=0.72 | Rs=1.21ws+1.40,R2=0.25 |
| 青海云杉林 Picea crassifolia forest | Rs=-28.05w+126.46,R2=0.57 | Rs=-43.47w+144.84,R2=0.55 | Rs=2.01ws-2.56,R2=0.35 |
| 祁连圆柏林 Sabina przewalskii forest | Rs=-23.41w+123.86,R2=0.72 | Rs=-24.58w+111.33,R2=0.64 | Rs=1.30ws-1.89,R2=0.42 |
| P值均<0.05 | |||
土壤呼吸作为一个复杂的生物学过程,受到多种因素的综合作用。对所涉及的与土壤呼吸有关的气温、土温、湿度、土壤含水量和风速等 14 个环境因子进行了主成分分析 (表 6),其结果表明:各植被类型第1公因子F1的贡献率均大于 40%,其中 0—60 cm 的土壤温度和土壤含水量有较大负荷,因此F1主要反映了 0—60 cm 土壤的水热状况。第2公因子F2中湿度、气温、风速、10 cm 土温等因子有较大负荷,所以F2主要反映土壤表层的环境因子。第3公因子F3主要反映太阳辐射的信息。
将土壤呼吸速率与新生成的 3 个主成分F1、F2、F3以及F综进行线性回归分析,拟合的线性方程和决定系数如表 7 所示。从拟合结果可以看出,各植被类型土壤呼吸和F1、F2、F3以及F综的相关性因植被类型而不同,总的来说和F1即 0—60 cm 土壤的水热状况相关性较高,其次为F2土壤表层的环境因子,相关性最小的为F3太阳辐射。由此可以推断水热条件是影响土壤呼吸的重要条件,土壤深层产生的CO2大部分均贮存于土壤孔隙中,向大气排放的速率大小受土壤表层环境因子的影响。
| 植被类型 Vegetation type | 次序 Order | 因素分量 Component | 特征值 Eigenvalue | 贡献率 Variance/% | 累积贡献率 Cumulative/% |
| 亚高山草原 | F1 | W20、W40、W60、T20、T40、T60 | 6.66 | 48.27 | 48.27 |
| Subalpine steppe | F2 | Taa、Tas、Ts、RHa、RHs、WS | 3.05 | 27.74 | 76.01 |
| F3 | PAR、Ras | 2.47 | 10.98 | 86.99 | |
| 干草原 | F1 | W20、W40、W60、T20、T40、T60 | 6.79 | 48.50 | 48.50 |
| Stipa purpurea steppe | F2 | RHa、RHs、Ts、Taa、Tas、WS | 4.10 | 29.27 | 77.77 |
| F3 | PAR、Ras | 1.85 | 13.22 | 90.99 | |
| 亚高山灌丛 | F1 | WS、Taa、W20、W40、W60、T20、T40、T60 | 7.63 | 54.50 | 54.50 |
| Subalpine shrub | F2 | RHa、RHs、Ts、Tas | 4.01 | 24.64 | 79.14 |
| F3 | PAR、Ras | 2.62 | 15.59 | 94.73 | |
| 青海云杉林 | F1 | WS、Taa、W20、W40、W60、T20、T40、T60 | 7.36 | 52.60 | 52.60 |
| Picea crassifolia forest | F2 | RHa、RHs、Tas、Ts | 4.20 | 29.99 | 82.59 |
| F3 | PAR、Ras | 2.15 | 15.37 | 97.96 | |
| 祁连圆柏林 | F1 | Ts、W20、W40、W60、T20、T40、T60 | 6.66 | 47.58 | 47.58 |
| Sabina przewalskii forest | F2 | RHa、RHs、Taa、Tas、WS | 3.89 | 27.81 | 75.39 |
| F3 | PAR、Ras | 2.12 | 15.16 | 90.55 | |
| Ts:10 cm土壤温度,Tas:地表温度,RHs:5 cm土壤湿度,Taa:大气温度,RHa:大气相对湿度,T20、T40和T60:0—20、20—40、40—60 cm土壤温度,W20、W40和W60:0—20、20—40、40—60 cm土壤含水量,WS:风速,PAR:光合有效辐射,Ras:太阳辐射 | |||||
| 植被类型 Vegetation type | F1 | F2 | F3 | F综 |
| 亚高山草原Subalpine steppe | Rs=6.33+0.43F1 R2=0.62 P=0.00 | Rs=6.33+1.28F2 R2=0.46 P=0.00 | Rs=6.33-0.36F3 R2=0.18 P=0.02 | Rs=6.33+0.37F1+1.26F2+0.14F3 R2=0.88 P=0.00 |
| 干草原 Stipa purpurea steppe | Rs=4.64+0.48F1 R2=0.62 P=0.00 | Rs=4.64-0.14F2 R2=0.24 P=0.02 | Rs=4.64+0.11F3 R2=0.20 P=0.04 | Rs=4.64+0.22F1+0.48F2-0.14F3 R2=0.69 P=0.00 |
| 亚高山灌丛 Subalpine shrub | Rs=3.46+0.03F1 R2=0.50 P=0.00 | Rs=3.46+0.06F2 R2=0.36 P=0.00 | Rs=3.46+0.76F3 R2=0.34 P=0.00 | Rs=3.46+1.03F1+0.02F2-2.14F3 R2=0.75 P=0.00 |
| 青海云杉林 Picea crassifolia forest | Rs=2.32+0.32F1 R2=0.51 P=0.00 | Rs=2.32-0.30F2 R2=0.47 P=0.00 | Rs=2.32-0.09F3 R2=0.21 P=0.01 | Rs=2.32+0.32F1-0.30F2-0.179F3 R2=0.70 P=0.00 |
| 祁连圆柏林 Sabina przewalskii forest | Rs=2.50+0.53F1 R2=0.0.75 P=0.00 | Rs=2.50+0.33F2 R2=0.46 P=0.00 | Rs=2.50-0.08F3 R2=0.27 P=0.04 | Rs=2.50+0.54F1+0.33F2-0.05F3 R2=0.88 P=0.00 |
本研究表明在整个生长季 5 种植被类型的土壤呼吸速率日变化均表现为单峰型曲线,亚高山草原、干草原、亚高山灌丛和青海云杉林在 14:00 时达到最大值,祁连圆柏林在 13:00 时达到最大值,可能与地表温度的差异有关,祁连圆柏林分布在研究区的阳坡,地表温度达到最大值要提前于其它类型。5种植被类型的土壤呼吸速率表现出明显的月变化,最大值在 7 月份,最小值在生长季初期 (表 3)。整个生长季5种植被类型平均土壤呼吸速率存在差异,表现为亚高山草原>干草原>亚高山灌丛>祁连圆柏林>青海云杉林 (表 2)。2013 年生长季期间青海云杉林土壤呼吸速率在 1.25—3.19 μmol m-2 s-1 之间,要小于Zheng等[22]的观测结果,亚高山草原土壤呼吸速率在 0.85—9.24 μmol m-2 s-1 之间,远远高于Chang等[23] 0.31—6.98 μmol m-2 s-1 的观测结果,虽然植被类型和土壤类型相同,研究结果的差异可能是研究区位置的不同,也可能是年与年之间的土壤呼吸速率波动的原因,也许与土地利用(放牧与否)有关。刘贤德等[24]在祁连山的浅山区研究表明,放牧草地与青海云杉林地和灌木相比,土壤呼吸速率最小,该结果与本研究结果相反,可能由于放牧降低草地的土壤呼吸速率。本研究区在 2013 年为禁牧期,没有放牧的干扰,草地土壤呼吸速率 (2.01—12.10 μmol m-2 s-1) 显著大于刘贤德等[24]的研究结果 (0.37—8.66 μmol m-2 s-1)。储金翔等[25]对川西亚高山林区不同土地利用方式的土壤呼吸研究发现,云杉人工林和冷杉原始林样地土壤呼吸速率均在 8 月份达到顶峰。王光军等[26]对樟树人工林土壤呼吸动态变化研究表明,7 月底土壤呼吸速率达到一年中的最大值,本研究中的最大值出现在 7 月份,高值的出现可能与温度有密切关系。Lou等[27]研究表明在夏季,较高土壤温度下根系生长活跃促进了根系呼吸,同时微生物活性较高促进了有机质转化从而增加微生物呼吸,因而产生了较高的土壤呼吸速率。有报道指出土壤呼吸速率的季节变化是由土壤温度、土壤湿度和物候期的改变造成的[8, 28]。Scott-Denton等[29]研究指出温度是土壤呼吸速率季节变化的主要控制因子,此外,与植物生理活动相关的物候期也影响土壤呼吸速率,Thomas等[30]发现土壤呼吸速率的增长与根系和生物量的增长紧密相关,Goulden等[31]认为夏季因冻土消融而增加的有效土壤体积也会增加土壤呼吸速率。关于根系、生物量、冻土消融对土壤呼吸速率的影响是下一步工作的重要内容。
3.2 土壤呼吸速率与温度的相关性尽管影响土壤呼吸速率的因素众多,但温度(包括土壤温度和空气温度)是最重要的因子,由此常被用来解释土壤呼吸速率的大部分变化[32, 33]。土壤温度升高对植物根系的呼吸、土壤微生物活动、凋落物和土壤中有机质分解都有促进和加速作用[34]。多数研究表明指数模型能够较好的描述土壤呼吸速率与温度之间的关系,本研究对不同植被类型土壤呼吸速率与 10 cm 土壤温度,地表温度和大气温度进行了指数模型拟合,发现指数模型在各植被类型下表现相对较好,相关系数地表温度>大气温度>土壤温度 (表 4),与李红生[4]、齐玉春[35]、张金霞[36]、Osozawa[37]等人的研究结果相似。5种植被类型的Q10值整体表现为亚高山灌丛(2.77)>干草原(2.22)>亚高山草原(1.95)>祁连圆柏林(1.61)>青海云杉林(1.31),这可能与群落组成和结构有关。青海云杉林和祁连圆柏林冠幅大,对地表的遮阴度高,地表温度和土壤温度受气温的影响相对小于其他植被类型,因此,土壤呼吸速率对温度的敏感度也相对较小。本文得出不同植被类型土壤呼吸速率日变化与地表温度同步,呼吸速率最大值的出现时间与地表温度一致,土壤温度滞后土壤呼吸 3—4 h (图 1),与有些研究报道一致[38, 39, 40]。而Gaumont-Guay[41]报道土壤呼吸速率滞后土壤温度 4—5 h,Riveros-Iregui等[42]得出土壤呼吸速率最大值出现在土壤温度最大值之前。但也有报道土壤呼吸速率与土壤温度有较好的同步性[43, 44]。直到现在,滞后类型的机理仍没有得到合理和公认的解释。
3.3 土壤呼吸速率与湿度的相关性土壤水分是影响土壤呼吸速率的重要因子,一方面它直接能抑制微生物活性和根系呼吸能力而影响土壤呼吸速率,另一方面,土壤温度与土壤水分的交互作用间接对土壤呼吸速率产生影响[45]。刘绍辉等[46]研究发现土壤水分的含量较充足,没有成为限制因子时土壤呼吸与土壤温度呈正相关关系;当水分含量作为限制因子时土壤温度和水分含量就成为共同起作用的因素。另有研究表明,只有土壤含水量达到土壤微生物永久性萎蔫点或超过了田间持水力的情况下,土壤呼吸速率才会明显地受到土壤水分的影响,如果水分的变化没有超出极端范围,并不足以影响微生物和植物根系的活动,则很难明显地测出水分对土壤呼吸的影响,此时土壤水分的影响很可能被其他因子的影响所掩盖[15, 47]。本研究土壤呼吸速率日动态变化表明,当土壤湿度下降时土壤呼吸速率增加,但土壤呼吸速率出现峰值时土壤湿度没有降到最低,在整个生长季土壤呼吸速率与土壤湿度存在显著的负相关关系。Martin and Bolstad[48]对 5 个不同森林类型土壤呼吸的研究发现土壤呼吸随立地平均土壤水分的增加而降低。Oberbauer等[49]在北极冻原的研究表明随着土壤地下水位的下降,土壤呼吸逐渐增加。但Davidson等[50]在Amazon Basin的放牧地和林地的研究发现土壤呼吸随着土壤含水量的降低而降低。还有些研究表明在有些地区,土壤呼吸速率在夏季主要受土壤含水量控制,但冬季时尽管土壤含水量很高,土壤呼吸却很低[51, 52]。
3.4 土壤呼吸速率与其他因子的相关性为了更准确的研究生态系统的土壤呼吸,仅仅研究温度和土壤水分是不够的,需要对土壤呼吸的影响因子进行分类[53, 54]。Alf Ekblad等[55]研究认为虽然土壤呼吸与土壤温度和土壤水分有关,但很少能解释土壤呼吸的季节变异。本研究主成分分析表明影响不同植被类型土壤呼吸速率的主要因素各不相同,但总的来说在整个生长季 0—60 cm 土壤的水热状况是主要的影响因素,其次是土壤表层的环境因子(温、湿度及风速的影响),最后为太阳辐射。风速对土壤呼吸速率的影响取决于土壤呼吸基座周围的植被灌丛高度和结构。常宗强等[56]对祁连山亚高山灌丛林的研究发现土壤呼吸的空间变异主要受温度、水分和植物根系的综合影响。Siyan等[57]研究发现异质森林中植被、根系、微生物群落在理解土壤呼吸变异模式方面的重要性,植被叶面积指数、生物量、植被凋落物、微生物群落等生物因子,土壤质地、有机质含量、土壤C/N等非生物因子以及人类活动均不同程度的影响着土壤呼吸,因此在具体分析土壤呼吸变化时需综合考虑这些因子的作用。为了进一步探明不同植被类型土壤呼吸的日和季节变异模式及其机制,需要开展多因素的长期观测研究,对了解其它相似植被类型的土壤呼吸可提供一定的参考依据,也可为研究区土壤碳循环和不同时间尺度植被作为碳源/汇作用的准确预测提供基础数据。
4 结论(1) 5种植被类型土壤呼吸速率日动态均呈单峰型曲线,除祁连圆柏林土壤呼吸速率在 13:00 时达到最大值外,其余4种均在 14:00 时达到最大值。地表温度也呈单峰型曲线,与土壤呼吸速率变化同步,土壤湿度呈现先减小后增加的趋势,在 17:00 时达到最小值,此时土壤温度升高到最大值。
(2) 5种植被类型的土壤呼吸速率的月变化动态呈先增加后降低的趋势,在 7 月份达到最大值。土壤呼吸速率大小表现为亚高山草原>干草原>亚高山灌丛>祁连圆柏林>青海云杉林,土壤呼吸速率的变化范围因植被类型的不同而有所差异。
(3) 5种植被类型土壤呼吸速率都与土壤温度、地表温度和大气温度具有显著的指数关系,且与地表温度和大气温度的相关性强于土壤温度。不同植被类型的土壤呼吸速率对温度的敏感性有所不同,在整个生长季土壤呼吸速率对温度的敏感性Q10表现为亚高山灌丛(2.77)>干草原(2.22)>亚高山草原(1.95)>祁连圆柏林(1.61)>青海云杉林(1.31)。
(4) 5种植被类型的土壤呼吸速率与湿度之间都存在显著的线性负相关,R2为 0.55—0.93,与风速呈显著的线性正相关关系。
(5) 通过主成分分析表明影响5种植被类型土壤呼吸速率的主要因素各不相同,整体表现为在整个生长季 0—60 cm 土壤的水热状况是第一影响因素,其次是土壤表层的环境因子,最后为太阳辐射。
| [1] | Parry M L, Canziani O F, Palutikof J P, van der Linden P J, Hanson C E. Climate Change 2007: Impacts, Adaptation and Vulnerability.Cambridge: Cambridge University Press, 2007: 7-22. |
| [2] | 韩广轩, 周广胜. 土壤呼吸作用时空动态变化及其影响机制研究与展望. 植物生态学报, 2009, 33(1): 197-205. |
| [3] | 张丽华, 陈亚宁, 李卫红, 赵锐锋, 葛洪涛. 干旱区荒漠生态系统的土壤呼吸. 生态学报, 2008, 28(5): 1911-1922. |
| [4] | 李红生, 刘广全, 王鸿喆, 李文华, 陈存根. 黄土高原四种人工植物群落土壤呼吸季节变化及其影响因子. 生态学报, 2008, 28(9):4099-4106. |
| [5] | Zhao L, Li Y N, Xu S X, Zhou H K, Gu S, Yu G R, Zhao X Q. Diurnal, seasonal and annual variation in net ecosystem CO2 exchange of an alpine shrubland on Qinghai-Tibetan plateau. Global Change Biology, 2006, 12(10): 1940-1953. |
| [6] | Shi P L, Sun X M, Xu L L, Zhang X Z, He Y T, Zhang D Q, Yu G R. Net ecosystem CO2 exchange and controlling factors in a steppe—Kobresia meadow on the Tibetan Plateau. Science in China Series D: Earth Sciences, 2006, 49(2): 207-218. |
| [7] | 吴力博, 古松, 赵亮, 徐世晓, 周华坤, 冯超, 徐维新, 李英年, 赵新全, 唐艳鸿. 三江源地区人工草地的生态系统CO2净交换、总初级生产力及其影响因子. 植物生态学报, 2010, 34(7): 770-780. |
| [8] | 张东秋, 石培礼, 张宪洲. 土壤呼吸主要影响因素的研究进展. 地球科学进展, 2005, 20(7): 778-785. |
| [9] | 鲍芳, 周广胜. 中国草原土壤呼吸作用研究进展. 植物生态学报, 2010, 34(6): 713-726. |
| [10] | 陈书涛, 刘巧辉, 胡正华, 刘艳, 任景全, 谢薇. 不同土地利用方式下土壤呼吸空间变异的影响因素. 环境科学, 2013, 34(3):1017-1025. |
| [11] | 彭家中, 常宗强, 冯起. 温度和土壤水分对祁连山青海云杉林土壤呼吸的影响. 干旱区资源与环境, 2008, 22(3): 165-169. |
| [12] | 常宗强, 冯起, 司建华, 苏永红, 席海洋, 郭瑞. 祁连山高山草甸土壤CO2 通量的时空变化及其影响分析. 环境科学, 2007, 28(10):2389-2395. |
| [13] | Raich J W, Tufekciogul A. Vegetation and soil respiration: correlations and controls. Biogeochemistry, 2000, 48(1): 71-90. |
| [14] | Palmroth S, Maier C A, McCarthy H R, Oishi A C, Kim H S, Johnsen K H, Katul G G, Oren R. Contrasting responses to drought of forest floor CO2 efflux in a Loblolly pine plantation and a nearby Oak-Hickory forest. Global Change Biology, 2005, 11(3): 421-434. |
| [15] | 陈全胜, 李凌浩, 韩兴国, 闫志丹, 王艳芬, 张焱, 袁志友, 唐芳. 温带草原11 个植物群落夏秋土壤呼吸对气温变化的响应. 植物生态学报, 2003, 27(4): 441-447. |
| [16] | Tang J W, Baldocchi D D. Spatial-temporal variation in soil respiration in an oak-grass savanna ecosystem in California and its partitioning into autotrophic and heterotrophic components. Biogeochemistry, 2005, 73(1): 183-207. |
| [17] | Luo Y Q, Wan S Q, Hui D F, Wallace L L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature, 2001, 413(6856):622-625. |
| [18] | Rey A, Pegoraro E, Tedeschi V, de Parri I, Jarvis P G, Valentini R. Annual variation in soil respiration and its components in a coppice oak forest in central Italy. Global Change Biology, 2002, 8(9): 851-866. |
| [19] | Xu M, Qi Y. Spatial and seasonal variations of Q10 determined by soil respiration measurements at a Sierra Nevadan forest. Global Biogeochemical Cycles, 2001, 15(3): 687-696. |
| [20] | Wildung R E, Garland T R, Buschbom R L. The interdependent effects of soil temperature and water content on soil respiration rate and plant root decomposition in arid grassland soils. Soil Biology and Biochemistry, 1975, 7(6): 373-378. |
| [21] | 李小胜, 陈珍珍. 如何正确应用SPSS 软件做主成分分析. 统计研究, 2010, 27(8): 105-108. |
| [22] | Zheng X L, Zhao C Y, Peng S Z, Jian S Q, Liang B, Wang X P, Yang S F, Wang C, Peng H H, Wang Y. Soil CO2 efflux along an elevation gradient in Qinghai spruce forests in the upper reaches of the Heihe River, northwest China. Environmental Earth Sciences, 2014, 71(5):2065-2076. |
| [23] | Chang Z Q, Feng Q, Si J H, Su Y H, Xi H Y, Li J L. Analysis of the spatial and temporal changes in soil CO2 flux in alpine meadow of Qilian Mountain. Environmental Geology, 2009, 58(3): 483-490. |
| [24] | 刘贤德, 车宗玺, 金铭, 敬文茂, 罗龙发, 马钰. 祁连山不同海拔梯度和放牧强度土壤呼吸变化特征. 干旱区研究, 2009, 26(1): 8-13. |
| [25] | 褚金翔, 张小全. 川西亚高山林区三种土地利用方式下土壤呼吸动态及组分区分. 生态学报, 2006, 26(6): 1693-1700. |
| [26] | 王光军, 李树战, 闫文德, 郑威, 禹茜. 樟树人工林土壤呼吸的动态变化. 中南林业科技大学学报: 自然科学版, 2008, 28(4): 118-122. |
| [27] | Lou Y S, Li Z P, Zhang T L. Carbon dioxide flux in a subtropical agricultural soil of China. Water, Air, and Soil Pollution, 2003, 149(1/4):281-293. |
| [28] | Yuste J C, Janssens I A, Carrara A, Ceulemans R. Annual Q10 of soil respiration reflects plant phenological patterns as well as temperature sensitivity. Global Change Biology, 2004, 10(2): 161-169. |
| [29] | Scott-Denton L E, Sparks K L, Monson R K. Spatial and temporal controls of soil respiration rate in a high-elevation, subalpine forest. Soil Biology and Biochemistry, 2003, 35(4): 525-534. |
| [30] | Thomas S M, Cook F J, Whitehead D, Adams J A. Seasonal soil-surface carbon fluxes from the root systems of young Pinus radiata trees growing at ambient and elevated CO2 concentration. Global Change Biology, 2000, 6(4): 393-406. |
| [31] | Goulden M L, Wofsy S C, Harden J W, Trumbore S E, Crill P M, Gower S T, Fries T, Daube B C, Fan S M, Sutton D J, Bazzaz A, Munger J W. Sensitivity of boreal forest carbon balance to soil thaw. Science, 1998, 279(5348): 214-217. |
| [32] | Lloyd J, Taylor J A. On the temperature dependence of soil respiration. Functional Ecology, 1994, 8(3): 315-323. |
| [33] | Rodeghiero M, Cescatti A. Main determinants of forest soil respiration along an elevation/ temperature gradient in the Italian Alps. Global Change Biology, 2005, 11(7): 1024-1041. |
| [34] | 刘春霞, 王玉杰, 王云琦, 冯华敏, 胡波. 重庆缙云山3 种林型土壤呼吸及其影响因子. 土壤通报, 2013, 44(3): 587-593. |
| [35] | Qi Y C, Dong Y S, Liu J Y, Li M F, Yang X H, Liu L X. Daily variation characteristics of CO2emission fluxes and contributions of environmental factors in semiarid grassland of Inner Mongolia, China. Science in China: Series D, 2005, 48(7): 1052-1064. |
| [36] | Zhang J X, Cao G M, Zhou D W, Zhao X Q, Zhou X M. Carbon dioxide emission of Mat cryo-sod soil in the Haibei alpine meadow ecosystem.Acta Ecologica Sinica, 2001, 21(4): 544-549. |
| [37] | Osozawa S, Hasegawa S. Diel and seasonal changes in carbon dioxide concentration and flux in an arid soil. Soil Science, 1995, 160(2): 117-124. |
| [38] | Frank A B, Liebig M A, Hanson J D. Soil carbon dioxide fluxes in northern semiarid grasslands. Soil Biology and Biochemistry, 2002, 34(9):1235-1241. |
| [39] | 周洪华, 李卫红, 杨余辉, 曹志超, 李稚. 干旱区不同土地利用方式下土壤呼吸日变化差异及影响因素. 地理科学, 2011, 31(2):190-196. |
| [40] | Jin Z, Qi Y C, Dong Y S. Diurnal and seasonal dynamics of soil respiration in desert shrubland of Artemisia Ordosica on Ordos Plateau of Inner Mongolia, China. Journal of Forestry Research, 2007, 18(3): 231-235. |
| [41] | Jassal R, Black A, Novak M, Morgenstern K, Nesic Z, Gaumont-Guay D. Relationship between soil CO2 concentrations and forest-floor CO2 effluxes. Agricultural and Forest Meteorology, 2005, 130(3/4): 176-192. |
| [42] | Riveros-Iregui D A, Emanuel E R, Muth D J, McGlynn B L, Epstein H E, Welsch D L, Pacific V J, Wraith J M. Diurnal hysteresis between soil CO2 and soil temperature is controlled by soil water content. Geophysical Research Letters, 2007, 34(17): LI7404. |
| [43] | Drewitt G B, Black T A, Nesic Z, Humphreys E R, Jork E M, Swanson R, Ethier G J, Griffis T, Morgenstern K. Measuring forest floor CO2 fluxes in a Douglas-fir forest. Agricultural and Forest Meteorology, 2002, 110(4): 299-317. |
| [44] | Tang J W, Baldocchi D D, Xu L K. Tree photosynthesis modulates soil respiration on a diurnal time scale. Global Change Biology, 2005, 11(8):1298-1304. |
| [45] | 李凌浩, 王其兵, 白永飞, 周广胜. 锡林河流域羊草草原群落土壤呼吸及其影响因子的研究. 植物生态学报, 2000, 24(6): 680-686. |
| [46] | 刘绍辉, 方精云. 土壤呼吸的影响因素及全球尺度下温度的影响. 生态学报, 1997, 17(5): 469-476. |
| [47] | Kucera C L, Kirkham D R. Soil respiration studies in tallgrass prairie in Missourt. Ecology, 1971, 52(5): 912-915. |
| [48] | Martin J G, Bolstad P V. Annual soil respiration in broadleaf forests of northern Wisconsin: influence of moisture and site biological, chemical, and physical characteristics. Biogeochemistry, 2005, 73(1): 149-182. |
| [49] | Oberbauer S F, Gillespie C T, Cheng W, Gebauer R, Serra A S, Tenhunen J D. Environmental effects on CO2efflux from riparian tundra in the northern foothills of the Brooks Range, Alaska, USA. Oecologia, 1992, 92(4): 568-577. |
| [50] | Davidson E A, Verchot L V, Cattànio J H, Ackerman I L, Carvalho J E M. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biochemistry, 2000, 48(1): 53-69. |
| [51] | Fernandez D P, Neff J C, Belnap J, Reynolds R L. Soil respiration in the cold desert environment of the Colorado Plateau(USA): abiotic regulators and thresholds. Biogeochemistry, 2006, 78(3): 247-265. |
| [52] | Keith H, Jacobsen K L, Raison R J. Effects of soil phosphorus availability, temperature and moisture on soil respiration in Eucalyptus pauciflora forest. Plant and Soil, 1997, 190(1): 127-141. |
| [53] | Risk D, Lisa K, Hugo B. Are soil respiration processes geographically invariant? / / Canadian Geophysical Union Meeting. Canada: Banff, 2002. |
| [54] | Campbell J L, Law B E. Forest soil respiration across three climatically distinct chronosequences in Oregon. Biogeochemistry, 2005, 73(1):109-125. |
| [55] | Ekblad A, Bostrom B, Holm A, Comstedt D. Forest soil respiration rate and d13 C is regulated by recent above ground weather conditions.Oecologia, 2005, 143(1): 136-142. |
| [56] | 常宗强, 冯起, 吴雨霞, 苏永红. 祁连山亚高山灌丛林土壤呼吸速率的时空变化及其影响分析. 冰川冻土, 2005, 27(5): 666-672. |
| [57] | Ma S Y, Chen J Q, Butnor J R, North M, Euskirchen E S, Oakley B. Biophysical controls on soil respiration in the dominant patch types of an old-growth, mixed-conifer forest. Forest Science, 2005, 51(3): 221-232. |