 2015, Vol. 35
2015, Vol. 35文章信息
- 阮得孟, 孙勇, 程嘉伟, 刘大伟, 鲁长虎
- NGUYEN Dacmanh, SUN Yong, CHENG Jiawei, LIU Dawei, LU Changhu
- 盐城自然保护区新洋港河口不同生境冬季鸟类群落组成及其梯度变化
- Winter bird community structure and gradient change in different habitats at Xinyanggang Estuary, Yancheng Nature Reserve
- 生态学报, 2015, 35(16): 5437-5448
- Acta Ecologica Sinica, 2015, 35(16): 5437-5448
- http://dx.doi.org/10.5846/stxb201405211044
-
文章历史
- 收稿日期: 2014-05-21
- 修订日期: 2015-02-16






2. 越南林业大学, 越南河内 156204;
3. 盐城湿地珍禽自然保护区, 盐城 224057
2. Faculty of Forest Resources and Environment Management, Vietnam Forestry University, Hanoi, Vietnam, 156204;
3. Yancheng Wetland Natural Reserve, Yancheng 224057, China
湿地是地球上生产力最高的生态系统之一,具有丰富的生物多样性,其中鸟类是湿地中主要的高等动物类群之一[1,2,3]。湿地为众多水鸟提供了栖息地,具有丰富的鸟类多样性[4,5],同时也是很多鸟类躲避人类活动干扰的天然场所[6]。人类的活动,如围垦、水产养殖等,导致了自然湿地破碎化、旱化、甚至丧失,湿地鸟类群落发生改变,一些水鸟的生存受到威胁[7]。湿地鸟类群落的物种组成受到人为干扰以及水体条件等不同的影响[8],鸟类群落季节性变化也受到明显的人类活动干扰[5]。湿地景观尺度上的变化影响到鸟类群落的丰富度,鸟类对生境的选择随着湿地景观的季节性变化而有所改变[9,10]。鸟类对湿地生境的这种改变具有不同的适应能力,一些鸟类能够适应围垦湿地生境[11,12],一些鸟类则能够适应水产养殖塘生境[13,14]。因此,湿地生境的改变会导致不同生境梯度中鸟类种类组成、代表或指示性物种等发生较大的变化。
沿海滩涂湿地处于海洋与陆地的交错地带,富含大量的鱼虾和贝类,是鸟类重要的栖息和繁殖地。盐城湿地珍禽国家级自然保护区是我国滩涂湿地的主要分布区之一,也是我国优先保护的17个生物多样性关键地区之一。近年来,随着沿海经济发展和对土地需求的增加,盐城湿地自然保护区碱蓬滩、芦苇滩、草滩等自然景观大面积下降而人工景观面积不断增加,逐渐构成了拥有不同水域范围、不同水位高度的自然湿地与人工湿地,在缓冲区、实验区由海向陆地方向大体形成了自然滩涂、人工养殖塘、农田及夹杂在其间的芦苇湿地生境梯度[15]。生境的梯度变化必然会对鸟类群落产生不同影响。因此,本研究在盐城湿地珍禽国家级自然保护区内,选择芦苇沼泽、农田、鱼塘和滩涂4种不同干扰程度的生境,运用样线法对越冬鸟类的种类、数量进行了调查,试图回答以下问题:1)不同生境鸟类指示种的组成差异;2)不同生境中鸟类群落组成及其梯度变化特点。这对于在沿海地区人为干扰与生境破碎化的影响下如何进行鸟类多样性保护具有借鉴意义。
1 材料与方法 1.1 研究地概况研究区域位于盐城自然保护区。该保护区是中国最大的海岸带保护区,地处江苏中部沿海,海岸线长582 km, 南北长约200 km,东西最大宽度约20 km,总面积247260 hm2, 其中核心区为22600 hm2。研究地点位于保护区的北缓冲区, 新洋港镇东北4 km,新洋港河入黄海河口处(33°38′— 33°39′N, 120°34′ — 120°35′E)(图1)。

|
| 图 1 研究地位置与样地示意图 Fig.1 Map of the study area and the 4 block locations 1: 芦苇沼泽,2: 农田,3: 鱼塘,4: 滩涂 |
研究区域内由陆向海生境主要由芦苇沼泽、农田、鱼塘、滩涂组成(图1)。
芦苇沼泽主要为人工芦苇湿地,部分人工芦苇(Phragmites australis)种植区冬季地表干旱,每块芦苇田被蓄水沟渠分割开来;部分芦苇湿地冬季保持一定水位,水深最深约1 m,除了芦苇植被还有空心莲子草(Alternanthera philoxcroides)等水生植被,人为利用于水产养殖;沿新洋港河河道边也生有少量原生芦苇;旱地芦苇收割与河道边原生芦苇收割是冬季此类型芦苇生境的主要人为干扰,水生芦苇湿地不收割芦苇,主要人为干扰是定期的鱼类捕捞,相对芦苇收割干扰较小。
农田生境靠近公路,道路两侧种植的防护林为意杨(Populus euramevicana)、柳树(Salix babylonica),田边排水渠内长有带状芦苇、空心莲子草等水生植被。冬季农田种有小麦(Triticum aestivum)、棉花(Gossypium sp)以及部分收割后稻田未利用。鸟类常栖息于田间林地、草地、灌木丛、田边空地和沟渠等地。主要人为活动有冬季小麦的种植和梅花的采摘。
鱼塘生境位于农田与滩涂的中间,生境类型以广阔明水面为主,塘岸为渔民生活区、电线杆,沟道边生长有灌木和芦苇、互花米草(Spartina alterniflora)等水生植被。所有鱼塘分期排水收鱼,鱼塘水位下降,露出大面积淤泥,翻塘、修整后塘底成为荒地。主要人为活动为收鱼活动。
滩涂是天然湿地,低潮位光滩无植被生长,在中、高潮位潮间带上长有互花米草、碱菀 (Tripolium pannonicum)等植物,退潮时潮间带显现出多条曲折的潮沟,裸露出约1 km宽的光滩。退潮时人类在潮间带的底栖动物采集为此生境最主要的人为干扰。
1.2 鸟类调查方法前期在盐城自然保护区缓冲区内沿着陆地向海方向,分别选择芦苇沼泽、农田、鱼塘、滩涂四种鸟类栖息生境,设置4个样地(图1),每个生境样地设置3条长度为3 km的样线。选择冬季鸟类群落较为稳定时期(2013年12月—2014年1月),采用样线法调查鸟类,记录两侧各宽100 m内鸟类种类、数量、行为习性等,步行速度为每小时1— 2 km。使用Leica77单筒望远镜(20 x 60倍)和OLYMPUS双筒望远镜(42×10倍)观察。由于鸟类对滩涂的利用主要集中在是退潮时光滩裸露的时间段内,所以为配合退潮时间以及对整个研究做更好的补充与讨论,选择晴好天气,分别在7:00—9:30、9:30—11:30、15:00—17:00 3个时段对这四种生境进行调查。
进行鸟类调查时,从样带内向两侧飞出的鸟类记入统计,从两侧飞入的鸟类则不记录,同样的,从样带前方向后方飞出的鸟类记入统计,不记录从后方飞入的鸟类。当鸟类从前一样带进入后一样带时,在后一样带的统计会扣除飞入鸟类的相应数量。当鸟类集群较小时,采用直接计数法,记录各种鸟类的种类和数量;在鸟类集群较大,同时鸟类又处于迅速活动因而无法准确统计到鸟类的数量时,辅助拍照估计集群数量。统计时采用最大值保留法[16],即从数次调查的同种鸟类统计数值中保留最大值代表该鸟类的数量。
1.3 数据分析 1.3.1 鸟类的物种组成特征每个生境,按照鸟类不同种群数量占鸟类统计总数的百分比(P)来确定优势种和数量级,将P>10% 定为优势种;1%<P<10% 定为常见种;0.1%<P<1% 的定为稀有种;P<0.1% 定为罕见种[16]。
1.3.2 鸟类的多样性计算每个生境7:00—9:30、9:30—11:30和15:00—17:00时段鸟类的丰富度指数(S)、均匀度指数(E)、辛普森多样性指数(D′)和香浓-维纳多样性指数(H′)[17,18,19]。
香浓-维纳Shannon-Wiener多样性指数 (H′):
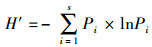
Simpson多样性指数(D′):
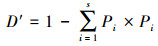
Pielou均匀度指数(E):
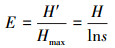
式中,S为物种数,Pi 为第i种物个体数与总个体数的比值; H′为实测群落的多样性指数值;Hmax为理论上最大多样性指数值。
1.3.3 分析方法多响应置换过程分析(MRPP)、指示种分析(ISA)需要 2 个或更多先验组的样本单位[20],将4个生境3时段调查-鸟种类个体数量数据矩阵,输入软件PC-ORD 6.0进行分析。MRPP采用Sorensen(Bray-Curtis)距离系数和n/sum(n)加权组法,来检查鸟类组成在各生境间是否存在差异。同时ISA进行确定每个生境的代表种类。
将观测条件-鸟类个体数量转化为资源利用百分比数据矩阵,输入软件PC-ORD 6.0进行双向聚类分析(Two-way cluster analysis),聚类方法采用Sorensen(Bray-Curtis)距离系数和组平均法,分析鸟类资源集团的结构。同时除趋势对应分析(Detrended correspondence analysis,DCA)与双向聚类分析法进行相互验证;从而归类相似生态位的鸟类,反映其栖息的生境特点。
2 结果与分析 2.1 鸟类种类组成与多样性本次调查中,共记录到鸟类12目67种(表1),其中非雀形目鸟类有45种,占所有鸟类种数的65.7%。
| 种类Species | 名称缩写Abbreviation | 优势度等级Dominance level | |||
| 芦苇沼泽Reed marshn=36 | 农田Farmn=36 | 鱼塘Fishpondn=36 | 滩涂Tidal flatsn=36 | ||
| *种群数量:+ + + + 为优势种, + + + 为常见种,+ + 为稀有种,+ 为罕见种,n为样本量 | |||||
| 小 Tachybaptus ruficollis | TARU | ++ | ++ | ++ | |
| 普通鸬鹚 Phalacrocorax carbo | PHCA | + | ++++ | ||
| 苍鹭 Ardea cinerea | ARCI | ++ | ++ | +++ | ++ |
| 白鹭 Egretta garzetta | EGGA | +++ | +++ | +++ | ++ |
| 大白鹭 Egretta alba | EGAL | ++ | ++ | ||
| 中白鹭 Egretta intermedia | EGIN | + | |||
| 绿鹭 Butorides striatus | BUST | ++++ | ++ | + | + |
| 大麻鳽 Botaurus stellaris | BOST | ++ | ++ | + | + |
| 夜鹭 Nycticorax nycticorax | NYNY | ++++ | |||
| 白琵鹭 Platalea leucorodia | PLLE | ++ | |||
| 豆雁 Anser fabalis | ANFA | ++ | |||
| 绿头鸭 Anas platyrhynchos | ANPL | ++ | +++ | ++ | |
| 斑嘴鸭 Anas poecilorhyncha | ANPO | +++ | +++ | +++ | +++ |
| 罗纹鸭 Anas falcata | ANFA | ++ | |||
| 红头潜鸭 Aythya ferina | AYFE | ++ | |||
| 白秋沙鸭 Mergus albellus | MEAL | ++ | |||
| 红隼 Falco tinnunculus | FATI | + | |||
| 苍鹰 Accipiter gentilis | ACGE | + | |||
| 日本鹌鹑 Coturnix japonica | COJA | ++ | ++ | + | + |
| 环颈雉 Phasianus colchicus | PHCO | +++ | + | ++ | |
| 普通秧鸡 Rallus aquaticus | RAAQ | ++ | |||
| 黑水鸡 Gallinula chloropus | GACH | ++ | ++ | + | + |
| 骨顶鸡 Fulica atra | FUAT | +++ | + | + | ++ |
| 白鹤 Grus leucogeranus | GRLE | + | |||
| 丹顶鹤 Grus japonensis | GRJA | ++ | +++ | ||
| 灰鹤 Grus grus | GRGR | ++++ | +++ | ||
| 灰斑鸻 Pluvialis squatarola | PLSQ | ++ | |||
| 环颈鸻 Charadrius alexandrinus | CHAL | ++ | ++ | ||
| 中杓鹬 Numenius phaeopus | NUPH | + | |||
| 白腰杓鹬 Numenius arquata | NUAR | +++ | |||
| 大杓鹬 Numenius madagascariensis | NUMA | +++ | |||
| 黑尾塍鹬 Limosa limosa | LILI | ++ | |||
| 鹤鹬 Tringa erythropus | TRER | ++ | |||
| 白腰草鹬 Tringa ochropus | TROC | ++ | |||
| 矶鹬 Actitis hypoleucos | ACHY | ++ | ++ | ||
| 扇尾沙锥 Gallinago gallinago | GAGA | + | |||
| 黑腹滨鹬 Calidris alpina | CAAL | +++ | ++++ | ||
| 黑尾鸥 Larus crassirostris | LACR | ++ | |||
| 织女银鸥 Larus vegae | LAVE | +++ | +++ | ||
| 红嘴鸥 Larus ridibundus | LARI | ++++ | |||
| 珠颈斑鸠 Streptopelia chinensis | STCH | ++ | ++ | ||
| 山斑鸠 Streptopelia orientalis | STOR | + | |||
| 普通翠鸟 Alcedo atthis | ALAT | ++ | + | ++ | |
| 戴胜 Upupa epops | UPEP | ++ | + | + | |
| 白鶺鸰 Motacilla alba | MOAL | + | + | ||
| 黄腹鹨 Anthus rubescens | ANRU | + | |||
| 白头鹎 Pycnonotus sinensis | PYSI | ++ | |||
| 棕背伯劳 Lanius schach | LASC | ++ | ++ | ++ | ++ |
| 楔尾伯劳 Lanius sphenocercus | LASP | + | |||
| 灰椋鸟 Sturnus cineraceus | STCI | ++ | |||
| 紫翅椋鸟 Sturnus vulgaris | STVU | +++ | |||
| 喜鹊 Pica pica | PIPI | +++ | +++ | +++ | ++ |
| 灰喜鹊 Cyanopica cyana | CYCY | ++ | |||
| 北红尾鸲 Phoenicurus auroreus | PHAU | ++ | ++ | + | + |
| 乌鸫 Turdus merula | TUME | + | |||
| 震旦鸦雀 Paradoxornis heudei | PAHE | +++ | ++ | +++ | |
| 棕头鸦雀 Paradoxornis webbianus | PAWE | +++ | +++ | + | |
| 中华攀雀 Remiz consobrinus | RECO | ++ | |||
| 麻雀 Passer montanus | PAMO | ++++ | ++++ | +++ | +++ |
| 金翅雀 Carduelis sinica | CASI | +++ | |||
| 小鹀 Emberiza pusilla | EMPU | ++ | +++ | + | ++ |
| 灰头鹀 Emberiza spodocephala | EMSP | ++ | +++ | ||
| 三道眉草鹀 Emberiza cioides | EMCI | +++ | ++ | ||
| 苇鹀 Emberiza pallasi | EMPA | +++ | +++ | +++ | +++ |
| 芦鹀 Emberiza schoeniclus | EMSC | +++ | ++ | ++ | |
| 田鹀 Emberiza rustica | EMRU | ++ | +++ | ++ | |
| 雪鹀 Plectrophenax nivalis | PLNI | + | |||
鱼塘生境的鸟类种类最多,为37种;芦苇沼泽、滩涂生境的鸟类多样性较高,滩涂生境的D′、 H′多样性指数分别在各生境不同时段中为最高和第二,芦苇沼泽生境的H′、D′多样性指数分别在各生境不同时段中为最高和第三。与其余生境比较,芦苇沼泽、滩涂的均匀度也更高。发现9:30—11:30是芦苇沼泽各种鸟类的休息时段,各项指数均低于其余两个时段,并且相差很大。农田和鱼塘生境不同时段鸟类活动变动很大,这可能是由于受到棉花收割以及排水收鱼的影响(表2)。
| 观测条件Observation | 平均个体数及其标准差Mean value and Standard deviation | 总个体数Total individuals | S | E | H′ | D′ |
| S:物种数Richness;E:均匀度Evenness;H′:Shannon-wiener多样性指数 biodiversity index;D′:Simpson多样性指数; 观测条件即不同时段在各生境的观测,其中Obs1:7:00—9:30-农田;Obs2:9:30—11:30-农田;Obs3:15:00—17:00-农田;Obs4:7:00—9:30-鱼塘;Obs5:9:30—11:30-鱼塘;Obs6:15:00—17:00-鱼塘;Obs7:7:00—9:30-滩涂;Obs8:9:30—11:30-滩涂;Obs9:15:00—17:00-滩涂; Obs10:7:00—9:30-芦苇沼泽;Obs11:9:30—11:30-芦苇沼泽;Obs12:15:00—17:00-芦苇沼泽 | ||||||
| Obs1(n=12) | 9.418±32.056 | 631 | 20 | 0.659 | 1.975 | 0.8147 |
| Obs2(n=12) | 1.687±9.955 | 113 | 10 | 0.499 | 1.148 | 0.4729 |
| Obs3(n=12) | 26.970±124.631 | 1807 | 27 | 0.474 | 1.564 | 0.6711 |
| 平均值Average | γ= 35 | 0.544 | 1.562 | 0.653 | ||
| Obs4(n=12) | 23.149±158.986 | 1551 | 23 | 0.246 | 0.772 | 0.2916 |
| Obs5(n=12) | 15.955±90.089 | 1069 | 25 | 0.413 | 1.328 | 0.5163 |
| Obs6(n=12) | 10.209±32.007 | 684 | 28 | 0.675 | 2.251 | 0.8406 |
| 平均值Average | γ= 37 | 0.445 | 1.45 | 0.5495 | ||
| Obs7(n=12) | 5.313±20.141 | 356 | 12 | 0.748 | 1.858 | 0.7738 |
| Obs8(n=12) | 16.194±58.358 | 1085 | 24 | 0.621 | 1.974 | 0.7941 |
| Obs9(n=12) | 21.403±103.687 | 1434 | 22 | 0.486 | 1.503 | 0.64 |
| 平均值Average | γ= 32 | 0.618 | 1.778 | 0.736 | ||
| Obs10(n=12) | 13.358±42.897 | 895 | 29 | 0.688 | 2.315 | 0.8335 |
| Obs11(n=12) | 11.642±80.053 | 780 | 15 | 0.281 | 0.761 | 0.2899 |
| Obs12(n=12) | 3.433±11.487 | 230 | 22 | 0.75 | 2.317 | 0.8205 |
| 平均值Average | γ= 35 | 0.573 | 1.798 | 0.648 | ||
MRPP分析表明,芦苇沼泽、农田生境的鸟类组成差异不显著(P=0.685>0.05);其余组合各生境间的鸟类组成具有显著差异(表3)。其原因是芦苇沼泽周围都是农田,造成鸟类在相邻生境下出现的机会增加,另外农田上仍有芦苇斑块,提供了类似沼泽的鸟类栖息环境。在总体上(12种观测条件),鸟类组成具有显著差异(P=0.0003<0.05);从T和A看来,不同观测值具有一定的分组界限和组内一致性,说明鸟类群落在不同生境以及不同时间段的划分具有可行性,同时也表明了这两个因素对鸟类群落的影响很大。
| 各生境间的比较 Habitat comparisons | 观测值 Observed | 预测值 Expected | 方差 Variance | 偏度 Skewness | T | A | P |
| T:检验统计量Test statistic;A:一致性统计量agreement statistic | |||||||
| 芦苇沼泽-农田Reed swamp-Farmland | 0.524 | 0.5 | 0 | 0.3 | 0.442 | -0.048 | 0.685 |
| 芦苇沼泽-鱼塘Reed swamp-Fishpond | 0.341 | 0.5 | 0 | -1.345 | -2.544 | 0.317 | 0.024 |
| 芦苇沼泽-滩涂Reed swamp-Tidal flat | 0.349 | 0.5 | 0 | -1.639 | -2.671 | 0.302 | 0.023 |
| 农田-鱼塘 Farmland-Fishpond | 0.317 | 0.5 | 0 | -1.481 | -2.604 | 0.365 | 0.023 |
| 农田-滩涂Farmland-Tidal flat | 0.333 | 0.5 | 0 | -1.448 | -2.581 | 0.333 | 0.024 |
| 鱼塘-滩涂 Fishpond-Tidal flat | 0.317 | 0.5 | 0 | -1.176 | -2.466 | 0.365 | 0.024 |
| 12种观测Observation | 0.272 | 0.5 | 0 | -0.434 | -4.276 | 0.455 | 0.0003 |
对不同生境下的鸟类作指示种分析,选出具有显著指示值的种类(表4)。灰斑鸻、黑腹滨鹬和白腰杓鹬都是滩涂生境常见的鸟类,黑腹滨鹬偶尔出现在鱼塘淤泥中。黑尾鸥、红嘴鸥、织女银鸥和苍鹭都是鱼塘生境的常见鸟,织女银鸥偶尔出现在光滩,苍鹭常常在芦苇沼泽及农田生境中觅食,偶尔出现在滩涂潮沟。灰鹤是农田常见的鸟类,偶尔出现在光滩,其对农田生境的指示值比较高,主要原因是该生境具有丰富食物资源。黑水鸡近水生活,分布较广,也常常在鱼塘生境中觅食,偶尔出现在农田及滩涂,然而其对芦苇沼泽生境具有较高的指示性,原因是该生境的水位、植被覆盖比较稳定。
| 种类Species | 生境Habitat | 指示值Indicator Value | 指示值随机化Randomization indicated value | P | |
| 平均值Mean | 标准Stand.Dev. | ||||
| 灰鹤Grus grus | 农田 | 92.7 | 41.9 | 19.37 | 0.0404 |
| 黑尾鸥Larus crassirostris | 鱼塘 | 100 | 34.2 | 17.91 | 0.0178 |
| 红嘴鸥Larus ridibundus | 鱼塘 | 100 | 33.9 | 18.24 | 0.0178 |
| 织女银鸥Larus vegae | 鱼塘 | 76.7 | 36 | 15.33 | 0.0374 |
| 苍鹭Ardea cinerea | 鱼塘 | 73.1 | 40.3 | 12.77 | 0.0178 |
| 黑腹滨鹬Calidris alpina | 滩涂 | 87.2 | 37.1 | 18.6 | 0.0376 |
| 灰斑鸻 Pluvialis squatarola | 滩涂 | 100 | 31.7 | 17.53 | 0.0186 |
| 白腰杓鹬Numenius arquata | 滩涂 | 100 | 31.2 | 18.28 | 0.0186 |
| 黑水鸡Gallinula chloropus | 芦苇沼泽 | 78.4 | 41.1 | 14.63 | 0.0174 |
双向聚类分析法把12种观测条件分为4大类(图2),与观测点所在的生境划分非常吻合。只有Obs11与Obs2观测到的鸟类较少被归为一类。鸟类的剩余信息越大,则在观测点的分布状况越相似,聚类越近。在剩余信息40%的情况下,67种鸟类大致被分为7个类群(图2):1)中华攀雀为一类,这种鸟类冬季成群,喜于茂密芦苇地栖息。2)生活于水面或水生植被的鸟类聚为一类,小、黑水鸡、骨顶鸡、普通翠鸟、白秋沙鸭、红头潜鸭、普通秧鸡、绿鹭、夜鹭、白鹡鸰、震旦鸦雀、三道眉草鹀等可在沼泽不同水深、芦苇植被中觅食。3)适应性较强或喜开阔农田的鸟类为一类群,灰鹤、豆雁、红隼、紫翅椋鸟、环颈雉等具有较长的活跃时段和较广的栖息生境。4)白头鹎、麻雀、田鹀、山斑鸠、珠颈斑鸠、灰椋鸟、灰喜鹊、乌鸫等对农田等人为干扰有较强适应性的鸟类为一类群。5)矶鹬、黑尾鸥、织女银鸥、红嘴鸥、鹤鹬等为一类群,这些鸟类可在开阔水塘觅食。6)白琵鹭为一类,其偏好泥泞水塘,在水中缓慢行进,嘴往两旁甩动以寻找食物,与其它的鱼塘鸟类不同。7)该类群均为冬季滩涂常见鸟类,可再细分为两类,其中普通鸬鹚、白腰草鹬、中杓鹬、灰斑鸻、白腰杓鹬、大杓鹬、黑腹滨鹬等在常见光滩或毗邻水域觅食;而灰头鹀、金翅雀、苇鹀等则多在米草丛中觅食。这7个类群的划分, 符合由于人类活动的不同程度干扰,导致由陆向海方向,形成芦苇沼泽、农田、鱼塘和滩涂4种不同的生境。也体现了生境变化后,鸟类为适应生境而呈现的梯度变化特点。
利用12种观测条件作为变量进行DCA排序(图3),排序结果表明,鸟类类群从轴1上可以分为3个类群,最左边为适应开阔水域环境(鱼塘)的鸟类,包括:白琵鹭、黑尾鸥、红嘴鸥、织女银鸥、大白鹭、小、斑嘴鸭等。中间为依赖水生植被环境(滩涂和芦苇沼泽)的鸟类,包括:常在光滩觅食的灰斑鸻、白腰杓鹬和大杓鹬;常见于芦苇丛的黑水鸡、骨顶鸡等。最右则为依赖陆地植被环境(农田)的鸟类,包括:苍鹰、白头鹎、灰鹤、紫翅椋鸟、麻雀等。从左到右大致为不同生境中水面面积与植被面积比率从高到低的一个连续梯度,对应的是鱼塘到农田。从轴2上看,农田、鱼塘、芦苇沼泽3个生境差异不显著,只有滩涂生境中Obs7、Obs9与Obs8在轴2上分开最为明显。以上表明生境间的差异(Axis1)以及不同的观测时间(Axis2)共同影响着不同生境中鸟类群落结构,并且生境之间的差异起着主导作用,同时也表明了潮汐对滩涂上鸟类群落组成的影响显著。

|
| 图 3 鸟类物种组成的DCA 排序 Fig.3 Detrended Correspondence Analysis for the clustering of bird species composition 图中物种名称缩写同表1; Axis1:生境间的差异;Axis2:不同的观测时间 |
鱼塘生境具有最高的鸟类丰富性,鱼塘生境包含了明水面、淤泥、荒地、建筑物、电线杆以及之间的电线、沟道等成分,组成较为多样化,表现出更强的内部异质性,而可利用资源的多样化有利于提高鸟类丰富度[21],适合多种鸟类栖息。滩涂生境的格局随着潮汐的变化明显,退潮时生境内部的异质性较高,能使鸟类在各自喜好的高程上取食,这种生态位的合理利用导致鸟种分布均匀,多种鸟类共存,因而鸟类群落的均匀性与物种多样性都较高。芦苇沼泽生境内具有明水面与植被,生境异质性较高,这可以为鸟类提供丰富食物和隐蔽条件好,吸引大量鸟类前来觅食和栖息。
虽然农田、鱼塘的鸟类群落都受到较高强度的人为干扰,但是各生境还留存一些草丛、灌丛和芦苇斑块,为鸟类提供了觅食与隐蔽场所而吸引大量鸟类前往,不过这些鸟类大多数为一些适应人类活动干扰的种类,比如环颈雉、白头鹎、麻雀、珠颈斑鸠等。相比之下,干扰强度低的芦苇沼泽具有高于滩涂、等于农田而低于鱼塘的鸟类丰富性,但较多专性的鸟类栖息于此,比如在芦苇层觅食的震旦鸦雀、在水面凋落物上觅食的秧鸡类、在浅水区觅食鹭类、在深水区觅食的鸭类与丹顶鹤等等。
值得提出的是,滩涂生境潮水的涨落对鸟类的生境选择有很大的影响。水鸟常会因为潮水涨落在鱼塘、滩涂、芦苇沼泽间移动。因此,本研究选择了3个时段(均为较低潮位)进行了群落调查,以便在各时段评估各生境鸟类群落的变化。每生境中不同时段鸟类活动变动较大(表2),9:30—11:30是芦苇沼泽和农田各种鸟类的休息时段,各项指数均低于其余两个时段,并且相差很大;鱼塘生境中虽然7:00—9:30时段鸟类个体数量最多,但鸟类群落的多样性指数最低;滩涂3个时段鸟类群落的多样性指数差异不显著,但7:00—9:30时段鸟类个体数量最低;这可能是由于受到冬季早上时段常排水收鱼的影响,收鱼活动会吸引大量滩涂鸟类前来觅食。从这一点看,15:00—17:00时间段的鸟类群落研究可能是最佳时段。
3.2 鸟类指示种对生境具有很好的响应模式鸟类常作为特定生境的指示种,利用鸟类表现出的特性,可对生境做出描述。一些在特定时间出现或在特定生境活动的种类,其种群数量不论大小,都可以较好地代表其栖息的环境特点。排序、聚类和指示种分析的方法可以在不清楚鸟类特性的情况下了解鸟类与其栖息的环境的关系,结合鸟类的生物学特性信息,就可以借此推测其栖息生境的异质性概况。
黑水鸡喜栖息于开阔水面,常于水面浮游植物间翻拣寻食,但其体型较小,水生植被正好可以为其提供保护,所以在四个生境中,芦苇沼泽生境更适合黑水鸡生活。在滩涂调查过程中,发现黑水鸡常在水面凋落物上取食或在沼泽边缘草地觅食,也常见其在明水面上作鸭类样游泳和作鹭类样涉水。与其余沼泽鸟类比较,黑水鸡更适应在不同水深及栖息地类型的芦苇沼泽生境栖息。
灰鹤迁徙时停歇和取食于农耕地。调查过程中发现其集小群在近棉花带小麦田中觅食,但集大群在开阔农田中休息,有人为干扰时灰鹤迅速起飞同时呼叫警报。这表明灰鹤能够很好地响应农田生境不同植被及人为干扰。
红嘴鸥是水域鸟类,调查过程中发现红嘴鸥喜集大群在水面上盘旋飞行或在刚排水收鱼的鱼塘中觅食。同时也记录到织女银鸥与黑尾鸥混群在淤泥中觅食或在荒地上休息。由此可以表明,这3种鸥对鱼塘生境不同栖息地做出响应,由于开阔的空间以及较好的飞行能力,这3种鸥的活动受到人为干扰的影响较小。
灰斑鸻在沿海滩涂及沙滩取食,白腰杓鹬多栖息于河口、海岸及沿海滩涂环境,黑腹滨鹬栖于海滩的近陆地段及内陆湿地。在调查过程中发现前两种鸟类主要是单独活动,退潮时光滩裸出,它们才结成稀疏的群体,仅依靠视觉觅食,而黑腹滨鹬则结成密集的群体依靠触觉觅食。涨潮时未见黑腹滨鹬,只见灰斑鸻和白腰杓鹬栖息于滩涂上互花米草丛中。然而在鱼塘调查过程中未见灰斑鸻和白腰杓鹬,只见小群黑腹滨鹬在淤泥中觅食。由此表明,这3种鸻鹬类对滩涂生境不同水位及食物资源做出响应,与其余两种相比,黑腹滨鹬对滩涂生境指示性较弱。
3.3 鸟类群落结构梯度变化明显鸟类群落结构的差异对不同的环境特点有指示作用[22,23]。芦苇沼泽和农田生境具有较多的相似种,具有相似生态位的鸟类总是栖息在相同的生境内,开垦的农田内保留着一些芦苇斑块,拥有与芦苇沼泽相似的生境类型。沿海鱼塘的开发增加了水面面积,导致一些适应海洋环境的鸟类成为优势种,如红嘴鸥、黑尾鸥和织女银鸥。芦苇沼泽和滩涂两种湿地生境均具有明水面、植被等内部生境因子,其植被密度及水位的差异成为吸引鸟类栖息的重要因素。芦苇沼泽的水面面积与植被面积比率较稳定,吸引了更多偏好该生境的沼泽鸟类。滩涂的水面面积与植被面积比率也较为稳定,但在一天时间内有所变动,涨潮时鸟类群落主要为依赖植被环境的鸟类,而退潮时在光滩、泥滩上觅食的鸟类占优。除了人为干扰强度弱以外,滩涂、芦苇沼泽的水面面积与植被面积比率稳定更有利于维持鸟类群落的稳定性,所以这两种生境鸟类多样性指数以及均匀度都较高,同时也表明这两种生境环境质量较优。事实上,不同生境的鸟类群落变化都受到自然和非自然因素的影响[24,25,26,27],鸟类群落组成以及结构变化影射出来的信息丰富,不只局限于生境异质性与稳定性。环境的好坏以及人为干扰的强弱不能只通过比较鸟类多样性得出定论,反之亦然,但通过观测与分析可知,水面面积与植被面积的比率对鸟类群落结构具有一定的影响。
天然湿地是鸟类栖息和繁殖的良好场所,在保护鸟类群落多样性方面意义重大,尤其在为水鸟提供停歇觅食的场所起着重要的作用。盐城自然保护区天然湿地包括芦苇沼泽和泥滩2种生境类型,但在1995至2010年的15a内丧失的天然湿地面积约12687 hm2,特别是在芦苇沼泽被开发为水产品养殖塘和农田之后,天然湿地消减越发严重。天然湿地面积逐渐减少,湿地生态功能明显下降,生物多样性降低。鉴于人类活动对湿地鸟类影响的不断加剧,建议在开垦农田、鱼塘和在光滩引入互花米草时应考虑到人类活动对鸟类分布和多样性的影响,同时进行科学的功能区划分,将人类的生产、生活等区域进行有效的布局,合理地确定水面面积与植被面积比率来维持鸟类多样性。
| [1] | Gallagher J L, Reimold R J, Linthurst R A, Pfeiffer W J. Aerial production, mortality, and mineral accumulation-export dynamics in Spartina alterniflora and Juncus roemerianus plant stands in a Georgia salt marsh. Ecology, 1980, 61(2): 303-312. |
| [2] | Kauppinen J. Temporal variation in the wetland waterfowl communities of the Finnish lake district. Ornis Fennnica, 1995, 72: 145-153. |
| [3] | Boorman L. Saltmarsh Review: An overview of coastal saltmarshes, their dynamic and sensitivity characteristics for conservation and management. JNCC Report No. 334. UK: Joint Nature Conservation Committee, 2003. |
| [4] | 杨月伟, 夏贵荣, 丁平, 马仁翻, 陈余钊. 浙江乐清湾湿地水鸟资源及其多样性特征. 生物多样性, 2005, 13(6): 507-513. |
| [5] | 钟福生, 颜亨梅, 李丽平, 蒋勇, 姚毅, 邓学建. 东洞庭湖湿地鸟类群落结构及其多样性. 生态学杂志, 2007, 26(12): 1959-1968. |
| [6] | Davis D S, Browne S. Natural History of Nova Scotia: Topics and Habitats. Canada: Nova Scotia Museum, 1996. |
| [7] | Moser M, Prentice C, Frazier S. A global overview of wetland loss and degradation. Technical session B of the 6th Ramsar COP. Wetlands International, Brisbane. 1996. |
| [8] | 李丽平, 王德良, 钟福生, 朱文博. 环境因子对湿地鸟类多样性的影响分析. 四川动物, 2007, 26(3): 588-591. |
| [9] | Fairbairn S E, Dinsmore J J. Local and landscape-level influences on wetland bird communities of the prairie pothole region of Iowa, USA. Wetlands, 2001, 21(1): 41-47. |
| [10] | Naugle D E, Johnson R R, Estey M E, Higgins K F. A landscape approach to conserving wetland bird habitat in the prairie pothole region of eastern South Dakota. Wetlands, 2001, 21(1): 1-17. |
| [11] | 蒋科毅, 吴明, 邵学新, 吕咏. 杭州湾及钱塘江河口南岸滨海湿地鸟类群落多样性及其对滩涂围垦的响应. 生物多样性, 2013, 21(2): 214-223. |
| [12] | 张斌, 袁晓, 裴恩乐, 牛俊英, 衡楠楠, 王天厚. 长江口滩涂围垦后水鸟群落结构的变化——以南汇东滩为例. 生态学报, 2011, 31(16): 4599-4608. |
| [13] | 金杰锋, 刘伯锋, 余希, 鲁长虎. 福建省兴化湾滨海养殖塘冬季水鸟的栖息地利用. 动物学杂志, 2008, 43(6): 17-24. |
| [14] | 华宁, 马志军, 马强, 宋国贤, 汤臣栋, 李博, 陈家宽. 冬季水鸟对崇明东滩水产养殖塘的利用. 生态学报, 2009, 29(12): 6342-6350. |
| [15] | 刘春悦, 张树清, 江红星, 王会, 孙妍. 江苏盐城滨海湿地景观格局时空动态研究. 国土资源遥感, 2009, (3): 78-83. |
| [16] | Howes J, Bakewell D. Shorebird studies manual. Cornell University: Asian Wetland Bureau, 1989, 55: 143-147. |
| [17] | 孙儒泳. 动物生态学原理. 北京: 北京师范大学出版社, 2001: 398-401. |
| [18] | 张海明, 萧前柱, 高中信. 多样性指数公式在鸟类群落中应用的探讨. 生态学杂志, 1990, 9(5): 50-55. |
| [19] | 马克平, 刘玉明. 生物群落的多样性测度方法 Ⅰ.α多样性的测度方法(下). 生物多样性, 1994, 2(4): 231-239. |
| [20] | Bruce M C, James B G. Analysis of Ecological Communities. Oregon: MjM Software Design, 2002: 188-204. |
| [21] | Hurlbert A H. Species-energy relationships and habitat complexity in bird communities. Ecology Letters, 2004, 7(8): 714-720. |
| [22] | Berg . Composition and diversity of bird communities in Swedish farmland-forest mosaic landscapes: The amount of forest (at local and landscape scales) and occurrence of residual habitats at the local scale are shown to be the major factors influencing bird community composition in farmland-forest landscapes in central Sweden. Bird Study, 2002, 49(2): 153-165. |
| [23] | Castelletta M, Thiollay J M, Sodhi N S. The effects of extreme forest fragmentation on the bird community of Singapore Island. Biological Conservation, 2005, 121(1): 135-155. |
| [24] | Faccio S D. Effects of ice storm-created gaps on forest breeding bird communities in central Vermont. Forest Ecology and Management, 2003, 186(1): 133-145. |
| [25] | Lemoine N, Bauer H G, Peintinger M, Bhning-Gaese K. Effects of climate and land-use change on species abundance in a central European bird community. Conservation Biology, 2007, 21(2): 495-503. |
| [26] | Miller J R, Wiens J A, Hobbs N T, Theobald D M. Effects of human settlement on bird communities in lowland riparian areas of Colorado (USA). Ecological Applications, 2003, 13(4): 1041-1059. |
| [27] | Schieck J, Song S J. Changes in bird communities throughout succession following fire and harvest in boreal forests of western North America: literature review and meta-analyses. Canadian Journal of Forest Research, 2006, 36(5): 1299-1318. |