 2015, Vol. 35
2015, Vol. 35文章信息
- 王艳廷, 冀晓昊, 张艳敏, 吴玉森, 安萌萌, 张芮, 王立霞, 张晶, 刘文, 李敏, 李文燕, 陈学森
- WANG Yanting, JI Xiaohao, ZHANG Yanmin, WU Yusen, AN Mengmeng, ZHANG Rui, WANG Lixia, ZHANG Jing, LIU Wen, LI Min, LI Wenyan, CHEN Xuesen
- 自然生草对黄河三角洲梨园土壤物理性状及微生物多样性的影响
- Effects of self-sown grass on soil physical properties and microbial diversity of pear orchards in yellow river delta
- 生态学报, 2015, 35(16): 5374-5384
- Acta Ecologica Sinica, 2015, 35(16): 5374-5384
- http://dx.doi.org/10.5846/stxb201402220306
-
文章历史
- 收稿日期: 2014-02-22
- 修订日期:






2. 临沂大学生命科学学院, 临沂 276005;
3. 济南军区黄河三角洲生产基地, 东营 257231
2. College of Life Science, Linyi University, Linyi 276005, China;
3. Production base of Jinan Military Region in the Yellow River Delta, Dongying 257231, China
In summary, our study shows that the culture of naturally grown grass in the orchard significantly increases microbial biomass C, N, and microbe diversity, and that uncultured species constitute the major part of the increased bacterial concentration. Moreover, sustained grass culture in the orchard benefits the up-regulation of soil microbial activity and respiration and the increasing amount of active microbes and total phospholipid fatty acids. It is also effective at stimulating the balanced utilization of six carbons by microbes.
近几年研究表明,果园生草利于蓄水保肥、改善土壤结构、增加有机质、提高土壤酶活、提升果品品质、减少环境污染以及有效防治病虫害等[1, 2, 3, 4],是我国当今果业可持续高效发展及生态农业建设的重要组成部分[5];在意大利、法国、美国及日本等果品生产先进国家的考察调研也发现,果园生草能够全面综合提高果园生产能力和效率,实现果园土壤可持续发展的良好生态和经济效益,在这些国家很难见到不生草果园[6, 7, 8, 9, 10, 11]。而土壤微生物作为土壤重要组成部分,参与土壤中约90%的土壤反应过程[12],其中微生物量多少、微生物群落组成、活性等可以用来衡量土壤质量好坏、肥力高低以及物质代谢的旺盛程度,一定程度反映了作物对养分的吸收利用与生长发育状况[13, 14, 15];同时,微生物作为土壤重要“源”和“汇”,还参与有机质分解、pH变化、温室气体产生、毒素降解、土壤结构稳定、功能维持、腐殖质和团粒结构形成以及C、N、P、S等养分的循环与转化,是土壤质量评价指标体系中不可缺少的组成部分[16, 17, 18, 19, 20]。因此,探讨生草果园土壤微生物多样性变化,对果园生草推广及全面评价果园生草改良土壤效果具有重要意义。
前人就不同生草草种、生草模式对不同树种果园土壤微生物影响进行了一定研究,吴家森等[21]研究生草山核桃林土壤微生物影响发现,生草4a后土壤微生物量碳(MBC)增加了138.61%—159.68%,3种生草处理土壤微生物碳源利用AWCD显著高于免耕;潘学军等[22]对盆栽纽荷尔脐橙土壤微生物数量及酶活性的变化进行了测定,结果表明柑橘生育期内,土壤微生物以细菌为主,真菌最少,间种绿肥提高了细菌和放线菌数量,自然生草则明显提高了土壤中真菌数量,相关分析还表明微生物数量与土壤氮素及有机质含量之间成显著正相关;陈伟等[23]研究清耕苹果园微生物特性发现,高产果园微生物数量多、多样性好;吴红英等[24]对间作不同芳香作物、自然生草条件下沙地梨园土壤微生物特性研究发现,间作芳香植物有效调节了土壤微生物数量及其群落中细菌、真菌和放线菌比例,且梨园间作香矢车菊和柠檬罗勒较自然生草调节效果更好。属滨海盐化潮土的山东东营济南军区黄河三角洲生产基地,含盐量达0.3%—0.4%,曾采取“大坑深栽,清耕除草”方式发展果业,出现建园成活率低等问题;1993年从国外引进白三叶、黑麦草、高羊茅等10余种草种,进行果业发展-人工种草结合试探,因白三叶引发红蜘蛛大爆发,黑麦草、高羊茅等根系发达,易引起表层土壤干燥等原因导致试验又遭失败;2003年梨园尝试自然生草,割草3—4次/a覆盖于地表,并撒施7.5—10kg尿素/667m2,取得了省力、优质、高效的初步成效,得到了周围果农的认可[5, 6]。梁博等[25]研究自然生草对黄河三角洲梨园土壤pH的影响,发现生草能显著降低株间和行间各土层土壤pH值,且随生草年限的增加效果更好;本课题组吴玉森等[2]研究发现,自然生草降低黄河三角洲梨园0—40cm土层土壤含盐量及增加土壤有机质含量的同时,随着生草年限增加有效提高了参试梨园耕作层(0—20cm)土壤酶活性及土壤矿质营养元素含量,还提升了果实鲜食品质,但有关黄河三角洲生产基地自然生草对梨园土壤物理性状和微生物多样性影响,至今未见研究报道。为此,本试验以山东东营济南军区黄河三角洲生产基地自然生草4a、6a和9a的梨园耕作层土壤为试材,以清耕为对照,探讨自然生草对梨园耕作层土壤物理性状、微生物量、土壤微生物呼吸、活性、活跃微生物量、磷酯脂肪酸总量及土壤微生物碳源利用情况等的影响,旨在丰富生草果园土壤微生物多样性理论研究体系,并为我国果园生草培肥地力技术的推广应用提供理论支撑和科学依据。
1 材料与方法 1.1 试验材料本试验于2012—2013年在山东农业大学作物生物学国家重点实验室和东营市黄河三角洲济南军区基地三团十三分场进行,试材为17年生的黄金梨品种[Pyrus pyrifolia (Burm.f.).cv. Nakai],砧木为杜梨,栽植密度为2m×4m。试验梨园面积14.7hm2,土壤为滨海盐化潮土,2011年测得含盐量0.3%,pH8.5,有机质12.8mg/kg,全氮0.52g/kg,全磷0.20g/kg,全钾21.58g/kg,碱解氮135mg/kg,速效磷20 mg/kg,速效钾152 mg/kg,立地条件一致,常规管理[2]。
1.2 试验处理试验设4个处理:连续自然生草4a、6a、9a和清耕;处理分别开始于2009年、2007年和2004年的春季进行全园自然生草,试验小区面积分别为2.7、3.3、7.3 hm2,清耕小区为1.3 hm2。随机区组设计,3次重复。各处理小区土壤及植株管理条件一致,以清耕为对照;草区刈割3—4次/a,就地覆盖,每次割草后撒施7.5—10 kg尿素/667m2,以促进草的生长(给草施肥),清耕区采用传统的方法施入同量尿素。
1.3 自然生草地被植物群落及产草量见表 1。
|
处理 Control | 优势草种 Dominant grasses | 鲜草量 Fresh weight/ (kg/667m2) | 生物量 Biomass/ (kg/667m2) |
|
自然生草4a Natural grass4 years | 蒲公英Herba taraxaci,伴生种有狗尾草Setaira viridis (L.)Beauv,稗子Echinochloa crusgalli (L.) Beauv.,虎尾草Chloris virgata Swartz等15种 | 2545 | 612. |
|
自然生草6a Natural grass 6 years | 虎尾草 Chloris virgata Swartz,伴生种有打碗花Calystegia hedercea Wall.,稗子Echinochloa crusgalli (L.) Beauv.)等6种 | 4983 | 1245 |
|
自然生草9a Natural grass 9 years | 马唐 Digitaria sanguinalis (L.) Scop,伴生种有虎尾草Chloris virgata Swartz,稗子Echinochloa crusgalli (L.) Beauv.等7种 | 5255 | 1318 |
试材随机区组设计,3次重复。土壤样品于2013年5月21日在各处理小区按5点取样法,用土钻取耕作层(0—20cm)土层土样,各小区5个样点混合均匀后作为1个土壤样品,共12份土样,剔除植物残根等杂物后用无菌塑料袋装好,带回实验室后分为3部分,一部分阴干、研磨、过筛保存;另一部分于-20℃冰箱保存,用于各类微生物指标的测定;第3部分在-80℃冰箱保存,用于细菌遗传多样性测定。
1.5 试验测定指标及方法 1.5.1 土壤物理特性的测定土壤含水量测定采用烘干法;容重测定采用环刀法;pH测定采用水土比2.5∶1酸度计测定;电导率采用水土比5∶1电导率仪测定;土壤孔隙度测定采用常规方法进行[26]。
1.5.2 微生物量及活性等的测定(1)微生物量的测定
微生物量C测定采用(氯仿)熏蒸提取-容量分析法,微生物量N采用(氯仿)熏蒸提取-茚三酮比色法[27]。
(2)土壤活跃微生物量、微生物活性及土壤呼吸的测定
土壤活跃微生物量、微生物活性及土壤呼吸的测定采用王芸等及高云超等的方法进行[28, 29]。其中土壤活跃微生物量测定采用呼吸曲线数学分析法,用葡萄糖培养基,其配方为葡萄糖3g、酵母膏0. 75g、NH4Cl 1.125g、MgSO47H2O0.3g、KH2PO40.25g、水1000mL,培养方法是1g新鲜土样中添加0.4mL培养基,20℃培养10h,测定CO2 量,计算公式为:X0= 0.283YCO2×10h/干土百分率,X0为活动生物量(mg生物量干重/(kg土))。YCO2/10h为10h培养期内CO2产生量〔mg-CO2 kg-1干土 10h- 1)。另外,44mgCO2=22.4mL CO2;土壤微生物活性测定采用土壤呼吸CO2测定法,5g鲜土于310mL试剂瓶中,22℃、24h测定CO2释放量;土壤呼吸测定采用CO2释放量法:取5 g鲜土于试剂瓶中,28℃培养24 h后测CO2呼吸量。上述3个指标土壤均以干土重计算,测定仪器均为Portable Gas Analyser(ADC BioSientific Ltd)。
(3)微生物磷酯脂肪酸(PLFA)的测定
微生物磷酯脂肪酸(PLFA)的测定采用土壤微生物分析——FAME法(fatty acid methyl esters)[30]进行。具体为:将15mL的0.2mol/L KOH甲醇溶液和4g新鲜土壤加到50mL(或者35mL)离心管中,加盖,在37℃下恒温培育1h(脂肪酸释放,并且甲酯化,每10min样品涡悬20s/1次);加3mL 1.0mol/L醋酸溶液中和pH值,充分摇匀。加10mL正己烷,(涡旋30s)混匀,使FAMEs转到正己烷相中,800r/min离心15min将正己烷相转到干净试管中,在N2气流下挥发掉溶剂,将提取的FAMEs溶解于0.8 mL 1∶1 的正己烷∶甲基叔丁基醚(methyl-tertbutyl ether)溶液中,充分溶解3—5 min,转入GC小瓶,同时加入10μL浓度为1mg/mL的内标C19:0并上机测定,作GC-MS(Shimadzu GC/MS-QP 2010)分析。以十九烷酸(19:0为内标)甲酯内标进行定量计算。
1.5.3 土壤细菌遗传多样性的测定(1)DGGE变性凝胶梯度电泳
①土壤DNA提取。采用OMEGA(Soil DNA Kit)试剂盒,具体按照试剂盒说明书进行。
②16SrDNA基因V8区PCR反应。以(1)中提取的DNA为模板,反应体系为50μL:2μL 10μmol/L 341F(5′-
③将各处理3平行PCR产物采用OMEGA公司Cycle-Pure Kit试剂盒回收纯化为50μL,具体步骤参照说明书进行。
④采用Bio-Rad公司Dcode型梯度胶制备装置,制备变性剂浓度从40%—70%的8%聚丙烯酰胺凝胶。待胶凝固后,加入PCR产物45μL和10×Loading Buffer 15μL的混合物,在80V电压、60℃条件下电泳16h。
⑤电泳结束后将凝胶放在DuRed核酸染液(泡染法)染色40—50min。采用UVP(Gel Doc-It Imaging System)成像系统拍照,并用(Bio-Rad)软件进行图像分析。
(2)DGGE部分条带测序
①选择DGGE电泳图中生草4—9a处理与对照相比亮度明显增加的条带(1—7共7条)进行回收测序。首先在紫外灯下快速切胶、捣碎置于1.5mL离心管(含900μL无菌水)中过夜;回收后进行PCR扩增并进行纯化,扩增采用TIANGEN公司2×Taq PCR Master Mix试剂,方法及条件严格按其说明书进行。扩增引物为:341F(5′-CCTACGGGAGGCAGCAG-3′),907R(5′-CCGTCAATTCCTTTGAGTTT-3′);
②扩增完成后同样采用1.5.2中(3)的方法进行纯化,然后采用TIAN GEN公司Zero Back Fast Ligation Kit及DH5α感受态细胞进行连接转化并挑选单克隆体,具体步骤参照试剂盒说明书进行,特征条带测序在英潍捷基(上海)贸易有限公司进行测定。
1.5.4 微生物碳源利用的测定微生物碳源利用的测定采用Biolog-Eco方法进行[32]。称取10g新鲜土样,在超净工作台中将土壤加至带有玻璃珠,内90mL灭菌0.85%NaCl溶液的三角瓶中,加盖振荡40min(转速为250r/min)。静置5min后按逐步稀释法,依次稀释为10-2、10-3土壤悬浮液.用八通道移液器向ECO板每孔中加入10-3的土壤悬浮液,接种量为150μL,将接种好的微孔板放在25℃下连续培养10d,每隔24h在590nm处用酶标仪(PerkinElmer USA)测定其光密度值,连续测定7d,选取96h(快速生长期)数据进行碳源利用分析。
1.6 数据处理与分析(1)本文所有试验数据均为3次重复,并采用Excel 2003和DPS7.05数据分析软件进行统计分析,并用邓肯氏法进行多重比较,差异显著性用不同大小写字母表示;DGGE条带使用quantity one-4.6.2 软件分析,聚类分析采用非加权组平均法(UPGMA);主成分分析采用SAS9.0软件分析。
(2)使用Shannon指数(H)、Margalef丰富度指数(D)和Pielou均匀度指数(E)来评价细菌群落遗传多样性,其计算公式为: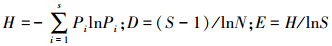 。式中Pi为某一条带的强度与同泳道中所有条带总强度的比值,即Pi= ni/N;S为每一泳道总的条带数[33]。
。式中Pi为某一条带的强度与同泳道中所有条带总强度的比值,即Pi= ni/N;S为每一泳道总的条带数[33]。
自然生草不同年限梨园土壤物理性状测定结果见表 2。由表 2可以看出,自然生草4—9a的总孔隙度是清耕对照的1.27倍,而土壤容重和电导率(可溶性含盐量)分别仅是清耕对照的85%和59%,差异均达极显著水平,各生草处理间差异不显著;自然生草9a的土壤PH7.67,是清耕对照(7.95)的96%,差异达显著水平,其他处理间差异不显著,而所有处理间的土壤含水量差异均不显著。
|
处理 Treatment | 总孔隙度/% Total porosity | 容重 Capacity Weight | 含水量/% Water content | 电导率/(μs/cm) Conductivity | pH |
| 同列中不同小写字母表示差异显著(P<0.05),不同大写字母表示差异极显著(P<0.01) | |||||
| 清耕对照Control | 35.46%bB | 1.71aA | 13.30%aA | 900.33aA | 7.95aA |
| 自然生草4a Natural grass 4 years | 44.84aA | 1.46bB | 12.76aA | 542.67bB | 7.76abA |
| 自然生草6a Natural grass 6 years | 44.33aA | 1.46bB | 14.00aA | 606.33bB | 7.75abA |
| 自然生草9a Natural grass 9 years | 45.81aA | 1.44bB | 13.27aA | 453.67bB | 7.67bA |
以上结果表明,自然生草明显提高了梨园土壤孔隙度、降低了土壤容重和电导率(可溶性含盐量),而对土壤含水量没有影响。
2.2 自然生草不同年限对梨园土壤微生物多样性影响 2.2.1 土壤微生物量变化自然生草不同年限梨园耕作层土壤微生物量测定结果见图 1。由图 1可以看出,清耕处理微生物量C、N含量均为最低,随着生草年限增加微生物量C、N逐渐增加,生草处理微生物量C含量分别为清耕处理的1.40、1.76和2.86倍,微生物量N为清耕处理的1.25、1.53和2.07倍,其中生草4a微生物量C、N与清耕对照差异均不显著,生草6—9a处理与清耕对照差异均达显著水平。

|
| 图 1 微生物量C、N Fig.1 Microbial biomass C,N 不同小写字母表示差异显著(P<0.05) |
本试验进一步通过DGGE-PCR方法对土壤细菌整体群落特点进行了测定。DGGE电泳结果见图 2。由图 2可以看出,与清耕对照相比,生草处理条带数量增多,亮度明显增亮。

|
| 图 2 16SrDNA 基因V8区DNA片段的DGGE分离 Fig.2 16SrDNA gene V8 region separated by DGGE |
DGGE条带聚类分析结果见图 3。由图 3可以看出,4种不同生草年限处理共聚为3类,首先清耕对照单独聚为一类,其次生草4a处理聚为一类,最后生草6a和9a聚为一类。

|
| 图 3 细菌DGGE胶片条带的聚类分析 Fig.3 Cluster dendrogram analysis of DGGE bands in gel |
选择生草4—9a处理与对照相比亮度明显增加的条带(1—7共7条)进行测序比对,条带测序比对结果(表 3)。由表 3可知,比对结果与已发表序列的相似性达到91%—100%,7条带共测定出16个序列,其中有13个序列为未培养菌类,占到总序列数的81%,即增加的多为未培养菌类。其中有球形杆菌科(Sphaerobacteraceae)、浮霉状菌目(Planctomycetales)、浮霉状菌(planctomycete)、(Chitinophagaceae)鞘脂杆菌目、(Comamonadaceae)丛毛单胞菌科及(Chitinophaga)泉发菌科等细菌菌类。
|
条带 Band | 序列号 Accession number | 序列比对结果 Most similar sequence | 相似度 similarity/% |
| 1 | AM935838.1 | Uncultured Sphaerobacteraceae bacterium partial 16S rRNA gene,clone AMAH5 | 99 |
| JN825587.1 | Uncultured Planctomycetales bacterium clone Alchichica_AL522_1B160 small subunit ribosomal RNA gene, | 96 | |
| JF703505.1 | Uncultured Chitinophagaceae bacterium clone L1-10 16S ribosomal RNA gene, | 99 | |
| 2 | AB231417.1 | Uncultured bacterium gene for 16S ribosomal RNA,partial sequence,clone: SBR-Sludge-27 | 92 |
| GQ202634.1 | Uncultured bacterium clone C51 16S ribosomal RNA gene, | 95 | |
| EU589311.1 | Uncultured soil bacterium clone 2_F1 16S ribosomal RNA gene, | 99 | |
| 3 | AB545742.1 | Comamonadaceae bacterium Gu-R-8 gene for 16S ribosomal RNA, | 100 |
| AB656423.1 | Uncultured bacterium RNA for 16S rRNA,partial sequence,clone: B0423R002_K08 | 96 | |
| KC331435.1 | Uncultured bacterium clone lp217 16S ribosomal RNA gene, | 99 | |
| 4 | FJ949399.1 | Uncultured planctomycete clone MS-C156 16S ribosomal RNA gene, | 97 |
| HM556976.1 | Uncultured bacterium clone BICH805 16S ribosomal RNA gene, | 97 | |
| HM186028.1 | Uncultured bacterium clone HDB_SION1004 16S ribosomal RNA gene, | 97 | |
| 5 | JN178907.1 | Uncultured planctomycete clone 29G 16S ribosomal RNA gene, | 91 |
| 6 | KC464820.1 | Chitinophaga sp. M43 16S ribosomal RNA gene, | 99 |
| 7 | KC555026.1 | Uncultured bacterium clone K3-295. 16S ribosomal RNA gene, | 100 |
| GQ162414.1 | Bacterium enrichment culture clone 1.89 16S ribosomal RNA gene, | 96 |
细菌遗传多样性指数结果见表 4。由表 4可以看出,生草4—9a处理条带数量增多,多样性指数、丰富度指数逐渐提高,如生草4a多样性指数和丰富度指数分别是清耕的1.14倍和1.35倍,生草9a是清耕的1.22倍和1.44倍;但生草处理对均匀性指数E影响不明显。
|
处理 Treatment | 条带数量(s) Number of bands | Shannon 多样性指数(H) Shannon diversity index | margalef丰富度指数(D) Margalef richness index) | Pielou 均匀度指数(E) Pielou evenness index |
|
清耕对照 Control | 21 | 2.70 | 3.72 | 0.89 |
|
自然生草4a Natural grass 4 years | 38 | 3.09 | 5.02 | 0.85 |
|
自然生草6a Natural grass 6 years | 41 | 3.00 | 5.63 | 0.81 |
|
自然生草9a Natural grass 9 years | 40 | 3.30 | 5.36 | 0.89 |
PCR-DGGE结果总体说明,自然生草有利于增加黄河三角洲梨园土壤未培养细菌种类和数量,随着生草年限的延长,细菌群落向丰富性、多样性发展。
2.2.3 土壤微生物呼吸、活性、活跃微生物量及微生物磷酯脂肪酸种类、总量变化自然生草不同年限梨园耕作层土壤微生物呼吸、活性、活跃微生物量、微生物磷酯脂肪酸(PLFA)种类及总量测定结果见表 5。由表 5可以看出,生草4a微生物活性、活跃微生物量及微生物磷酯脂肪酸总量与清耕对照差异不显著,但生草6—9a处理土壤微生物呼吸、活性、活跃微生物量及微生物磷酯脂肪酸总量分别为清耕对照的2.26倍、2.75倍、1.86倍和1.57倍,差异达显著或极显著,但生草6a和9a处理二者间比较差异均不显著;而较清耕对照,生草处理对微生物磷酯脂肪酸(PLFA)种类影响较小,相互间差异均不显著。
|
处理 Treatment | 土壤微生物呼吸 Soil microbial respiration/ (mg kg-1 h-1) | 活性 Active/ (mg kg-1 h-1) | 活跃微生物量 Active microbial biomass/ (mg/kg干土) | plfa种类 plfa Species | plfa总量 plfa Total/ (μg/g) |
| 清耕对照Control | 2.90bB | 1.99bB | 50.76bC | 15 aA | 16.65bA |
| 自然生草4a Natural grass 4 years | 5.72aA | 3.56bAB | 61.77bBC | 17 aA | 20.95abA |
| 自然生草6a Natural grass 6 years | 6.38aA | 5.32aA | 89.83aAB | 17 aA | 25.92aA |
| 自然生草9a Natural grass 9 years | 6.75aA | 5.63aA | 99.28aA | 17 aA | 26.47aA |
综上,持续多年自然生草有利于梨园耕作层土壤微生物呼吸、活性、活跃微生物量及微生物磷酯脂肪酸总量等的提高。
2.3 自然生草不同年限对梨园土壤微生物碳源利用的影响Biolog-ECO板的碳源共为6类31种,不同年限自然生草梨园土壤微生物对6类碳源的利用情况见图 4。由图 4可知,不同处理对6类碳源利用情况差异明显。清耕土壤微生物对聚合物类利用程度很高,但对碳水化合物类和胺类利用程度很低,而生草处理对碳水化合物利用程度很高,对胺类利用程度却很低;各处理间清耕碳源利用程度最低,生草4a处理6类碳源利用程度最高,但各类碳源利用差异最大;生草6a处理6类碳源利用更为均衡,除聚合物类与胺类之间利用差异显著外,其余任两种碳源之间利用差异都不显著;而生草9a处理6类碳源利用程度较小,但碳源利用最为均衡,6类碳源两两之间利用差异均不显著。

|
| 图 4 同一生草年限土壤微生物对不同碳源的利用情况 Fig.4 Microbial utilization of different carbon sources in grass soil of the same years |
上述结果表明,自然生草改变了土壤微生物碳源利用结构,提高了6类碳源利用能力,其中以生草4a为最大,随着生草年限增加,6类碳源利用更加均衡。
2.4 板孔平均颜色变化率动态变化吸光值平均颜色变化率(AWCD值)某种程度上反映了土壤中快速生长微生物对单一碳源利用能力,能真实反映出快速生长微生物同化分解有机碳的能力。连续培养10d,每隔24h测得AWCD值见图 5。

|
| 图 5 不同生草年限处理土壤微生物碳源利用AWCD值动态变化 Fig.5 The different grass life processing carbon utilization of soil microorganismsdynamic change of AWCD value |
由图 5可知,总体上不同处理都表现为AWCD值随培养时间的延长而不断提高。其中,前期(24—72h)增长比较缓慢,紧接着(72—120h)增长迅速,后期(120h后)则增长缓慢并且逐渐趋于平稳。在连续培养10d中微生物群落代谢的AWCD值随时间变化曲线的形状符合一般微生物利用基质的规律,即存在较明显的适应期、对数期和稳定期等阶段。在培养10d时间里,生草4—9a处理AWCD值均明显高于清耕对照;生草处理间,吸光值生草4a最高、生草6a与其接近,生草9a次之。
上述结果表明,与清耕对照相比,自然生草能有效提高黄河三角洲梨园快速生长微生物碳源利用能力(AWCD值),其中生草4a提高幅度最大,生草6a与之接近,生草9a次之。
2.5 主成分分析BIOLOG的主成分分析显示了微生物群落对碳源利用的整体响应,是反映土壤微生物群落结构特征的有效手段。选择96h(快速生长期)测定的吸光值进行主成分分析(PCA)。前3个主成分累计方差贡献率达到了100%,根据提取的主成分个数一般要求累计方差贡献率达到85%,提取主成分特征值大于1的原则,共提取了2个主成分,评价各处理土壤微生物群落在碳源利用上的整体差异。第一和第二主成分累计贡献率达93.01%。因此本文对前2个主成分进行分析(图 6)。其中,第一主成分贡献率为69.45%,第二主成分贡献率为23.56%。

|
| 图 6 不同生草年限处理土壤微生物碳源利用主成分分析 Fig.6 The different grass life treatment soil microbial carbon source using principal component analysis |
由图 6可以看出,不同处理间土壤微生物群落对碳源利用能力差异明显,表现在它们在PC1和PC2的得分系数差异巨大,即位于不同象限。整体上看,与清耕对照相比,各生草处理空间位置都在向PC1和PC2正方向移动,即生草处理后微生物碳源利用变化趋势一致。
3 讨论 3.1 自然生草对梨园土壤微生物量及碳源利用等的影响土壤微生物量是指土壤中除植物根茬等残体和大于5×103μm3土壤动物以外具有生命活力的有机物质的量[34],其代表参与土壤能量和养分循环以及有机物质转化所对应微生物的数量,主要类群为细菌、真菌、放线菌、藻类和原生动物等。主要有两方面的功能:一方面微生物量是土壤有机质及土壤养分(N、P、S等)转化和循环的动力,参与土壤有机质分解、腐殖质形成、土壤养分转化、循环的各个过程;另一方面土壤微生物量是土壤养分(N、P、S)的贮备库,是植物生长所需速效养分的重要来源[35]。本试验发现生草处理后土壤微生物量C、N逐年增加,微生物量C含量分别为清耕处理的1.40、1.76和2.86倍,微生物量N为清耕处理的1.25、1.53和2.07倍,生草6—9a处理与清耕对照差异均达显著水平,这与前人[21, 22]研究结果类似,微生物量的明显提高,表明土壤肥力的不断改善。
本试验进一步对微生物碳源利用情况进行了测定,结果(图 4)发现生草4—9a处理碳源利用程度均高于清耕对照,这与前人研究结果基本一致[21]。分析是生草后大量凋落物归还土壤,这些凋落物及土壤中根系分泌物富含营养物质,再加上生草后昆虫、土壤动物数量增加从而形成了碳源代谢能力较强的微生物群落结构,而清耕土壤,由于清耕除草导致有机物质归还量较低、土壤团聚体破坏、有机碳矿化加快,同时由于地面裸露导致水土流失严重、耕作层温度变化幅度大等原因,不利于微生物生长繁殖。各处理间,生草4a碳源利用程度最高,但6类碳源利用差异巨大,而生草6—9a处理6类碳源利用却更为均衡;分析认为生草4a处理植物残体、根系分泌物等猛然积累提供了微生物所需各类营养物质,土壤温度变化幅度减小,适于微生物生存繁殖,土壤中原有优势菌群数量迅速变化,导致生草4a碳源利用为最大。同时,各种类草、昆虫及土壤动物等多因素竞争加剧,导致生草4a处理6类碳源利用情况差异巨大,而在生草6—9a草、根系分泌物种类、数量等等因素逐渐趋于动态平衡,从而生草6—9a 6类碳源利用情况比较均衡、稳定;另外,尽管Biolog-ECO板技术测定微生物碳源利用情况已广泛用于微生物的研究,但其仅适合快速生长微生物,且碳源种类也具有很大人为选择性,难以反映生长缓慢及不可培养微生物碳源利用情况[36, 37]。因此,有必要结合新方法从不同角度对自然生草土壤微生物碳源利用做进一步研究,从而得到更为全面结果。
PCR-DGGE技术是从微生物基因多样性角度研究微生物群落结构的方法,DGGE条带数量基本体现了微生物种群数量,而条带亮度则反映了该菌类数量的多少[38]。本试验DGGE图谱可以看出,生草4—9a处理较清耕对照土壤细菌数量、种类都明显增加,进一步对生草后明显增亮条带通过回收、转克隆及测序技术得出81%为未培养菌类,其中包括球形杆菌科(Sphaerobacteraceae)、浮霉状菌目(Planctomycetales)、鞘脂杆菌目(Chitinophagaceae)、丛毛单胞菌科(Comamonadaceae)、浮霉状菌(planctomycete)及泉发菌科(Chitinophaga)等细菌菌类。有研究表明(Comamonadaceae)丛毛单胞菌科细菌对COD和氨氮具有很好去除能力,对好氧颗粒污泥除污能力具有很好的强化作用[39];浮霉状菌目(Planctomycetales) 是细菌域中分化较早的一个分支,该目包括2个科9个属,其中4个属归入 (Planctomycetaceae) 浮霉状菌科,为化能异养型好氧菌[40],本试验测序结果中也检测到了 (Planctomycetaceae) 浮霉状菌科序列,这也说明生草后土壤通气状况的改善。与此同时,由于测序结果多是未培养菌类,难以获得其在土壤中的生态功能,想进一步了解这些优势菌群的结构功能和种属特性,需做更深入的研究。未培养菌类可能在自然生草改良梨园土壤过程中起主要作用,是下一步研究的重点和切入点。
3.2 自然生草对梨园土壤微生物呼吸、活跃微生物量及微生物总量的影响罗海峰等[41]认为可培养微生物种类占土壤微生物种类总数的0.1%—1%,且分离培养方法难以全面反映土壤微生物多样性的原始状态等,本试验对梨园土壤微生物呼吸、活跃微生物量及微生物磷酯脂肪酸总量(PLFA总量)等进行了测定。土壤呼吸主要是由土壤微生物的活动引起,代表土壤碳素的周转速率及微生物的整体活性[42];活跃微生物量是土壤中存在的少部分生理功能活跃微生物,这部分微生物一般不到总量的1/3[28]。本试验结果(表 5)发现生草4a活跃微生物量及微生物磷酯脂肪酸总量与清耕对照差异不明显,但生草6—9a土壤微生物呼吸、活跃微生物量及微生物磷酯脂肪酸总量分别为清耕对照的2.26倍、2.75倍、1.86倍和1.57倍,差异均达显著或极显著。分析认为土壤呼吸的提高一方面是由于活跃微生物量、微生物总量(plfa总量)的增加引起,这在表 5中可以看出;另一方面土壤呼吸、活跃微生物量的变化主要受土壤温度和土壤通透性影响,有关试验证据表明,几乎所有研究都显示土壤温度强烈地影响土壤微生物活性和呼吸[43];对麦田和玉米田的土壤呼吸研究得出土壤呼吸受5cm地温的影响最大[44, 45];高云超[29]等于不同年份免耕试验中发现影响土壤活跃微生物量的主要因素是土壤通透性或营养因素,并且李会科等[46]研究得出果园生草能有效调节5—20cm土层土壤温度变化,明显减小了温度波动范围;进一步分析认为温度影响土壤微生物呼吸、活跃微生物量的机理是温度与土壤酶活性存在密切关系,温度过高或过低均会造成生物酶失活或活性降低,致使微生物活性下降。这也从微生物角度说明自然生草有助于增加土壤营养状况,改善土壤通透性,减小土壤温度波动范围。
4 结论(1)自然生草优化了黄河三角洲梨园耕作层土壤物理性状,增加了微生物量C、N,其中细菌主要增加的是未培养菌类;
(2)持续多年自然生草,有利于参试梨园土壤微生物呼吸、活性、活跃微生物量及磷酯脂肪酸总量的提高,并对微生物均衡利用6类碳源作用明显。
| [1] | 李会科, 张广军, 赵政阳, 李凯荣. 渭北黄土高原旱地果园生草对土壤物理性质的影响. 中国农业科学, 2008, 41(7): 2070-2076. |
| [2] | 吴玉森, 张艳敏, 冀晓昊, 张芮, 刘大亮, 张宗营, 李文燕, 陈学森. 自然生草对黄河三角洲梨园土壤养分、酶活性及果实品质的影响. 中国农业科学, 2013, 46(1): 99-108. |
| [3] | 赵政阳, 李会科. 黄土高原旱地苹果园生草对土壤水分的影响. 园艺学报, 2006, 33(3): 481-484. |
| [4] | 霍颖, 张杰, 王美超, 姚允聪. 梨园行间种草对土壤有机质和矿质元素变化及相互关系的影响. 中国农业科学, 2011, 44(7): 1415-1424. |
| [5] | 陈学森, 王艳廷. 力推果园生草, 建设生态文明. 烟台果树, 2014, (1): 1-4. |
| [6] | 陈学森, 高东升, 辛力, 原永兵, 刘成连, 李文燕. 给果园小草一点阳光, 苹果更灿烂——苹果果实品质提升的途径与关键技术. 落叶果树, 2012, 44(3): 1-4. |
| [7] | 陈学森, 苏桂林, 姜远茂, 毛志泉. 可持续发展果园的经营与管理——再谈果园生草培肥地力及其配套技术. 落叶果树, 2013, 45(1): 1-3. |
| [8] | 陈学森, 韩明玉, 苏桂林, 刘凤之, 过国南, 姜远茂, 毛志泉, 彭福田, 束怀瑞. 当今世界苹果产业发展趋势及我国苹果产业优质高效发展意见. 果树学报, 2010, 27(4): 598-604. |
| [9] | 陈学森, 郝玉金, 杨洪强, 毛志泉, 姜远茂, 彭福田, 沈向, 张继祥, 陈晓流, 束怀瑞. 我国苹果产业优质高效发展的10项关键技术. 中国果树, 2010, (4): 65-67. |
| [10] | Whitelaw-Weckert M A, Rahman L, Hutton R J, Coombes N. Permanent swards increase soil microbial counts in two Australian vineyards. Applied Soil Ecology, 2007, 36(2/3): 224-232. |
| [11] | Moreno B, Garcia-Rodriguez S, Cańizares R, Castro J, Benítez E. Rainfed olive farming in south-eastern Spain: Long-term effect of soil management on biological indicators of soil quality. Agriculture, Ecosystems and Environment, 2009, 131(3/4): 333-339. |
| [12] | Coleman D C, Crossley D A. Fundamentals of Soil Ecology. London: Academic Press, 1996. |
| [13] | Knight B P, McGrath S P, Chaudri A M. Biomass carbon measurements and substrate utilization patterns of microbial populations from soil amended wish cadmium, copper, or zinc. Applied and Environmental Microbiology, 1997, 63(1): 39-43 |
| [14] | 赵彤, 闫浩, 蒋跃利, 黄懿梅, 安韶山. 黄土丘陵区植被类型对土壤微生物量碳氮磷的影响. 生态学报, 2013, 33(18): 5615-5622. |
| [15] | 郭天财, 宋晓, 马冬云, 查菲娜, 岳艳军, 张煜, 李耀昭. 氮素营养水平对小麦根际微生物及土壤酶活性的影响. 水土保持学报, 2006, 20(3): 129-131,140-140. |
| [16] | Green J L, Holmes A J, Westoby M, Oliver I, Briscoe D, Dangerfield M, Gillings M, Beattie A J. Spatial scaling of microbial eukaryote diversity. Nature, 2004, 432(7018): 747-750. |
| [17] | Nachimuthu G, King K, Kristiansen P, Lockwood P, Guppy C. Comparison of methods for measuring soil microbial activity using cotton strips and a respirometer. Journal of Microbiological Methods, 2007, 69(2): 322-329. |
| [18] | 孙波, 赵其国, 张桃林, 俞慎. 土壤质量与持续环境-Ⅲ. 土壤质量评价的生物学指标. 土壤, 1997, 29(5): 225-234. |
| [19] | 孔维栋, 朱永官, 傅伯杰, 陈保冬, 童依平. 农业土壤微生物基因与群落多样性研究进展. 生态学报, 2004, 24(12): 2894-2900. |
| [20] | 申卫收, 林先贵, 张华勇, 尹睿, 段增强, 施卫明. 不同施肥处理下蔬菜塑料大棚土壤微生物活性及功能多样性. 生态学报, 2008, 28(6): 2682-2689. |
| [21] | 吴家森, 张金池, 钱进芳, 黄坚钦. 生草提高山核桃林土壤有机碳含量及微生物功能多样性. 农业工程学报, 2013, 29(20): 111-117. |
| [22] | 潘学军, 张文娥, 樊卫国, 蓬桂华, 罗国华. 自然生草和间种绿肥对盆栽柑橘土壤养分、酶活性和微生物的影响. 园艺学报, 2010, 37(8): 1235-1240. |
| [23] | 陈伟, 姜中武, 胡艳丽, 束怀瑞. 苹果园土壤微生物生态特征研究. 水土保持学报, 2008, 22(3): 168-171. |
| [24] | 吴红英, 孔云, 姚允聪, 毕宁宁, 亓丽萍, 付占国. 间作芳香植物对沙地梨园土壤微生物数量与土壤养分的影响. 中国农业科学, 2010, 43(1): 140-150. |
| [25] | 梁博, 刘成连, 王永章, 李文燕, 原永兵. 黄河三角洲梨园自然生草对土壤PH的影响. 中国农学通报, 2014, 30(1): 143-148. |
| [26] | 杜森, 高祥照. 土壤分析技术规范. 北京: 中国农业出版社, 2006. |
| [27] | 李振高, 骆永明, 滕应. 土壤与环境微生物研究法. 北京: 科学出版社, 2008. |
| [28] | 王芸, 李增嘉, 韩宾, 史忠强, 宁堂原, 江晓东, 郑延海, 白美, 赵建波. 保护性耕作对土壤微生物量及活性的影响. 生态学报, 2007, 27(8): 3384-3390. |
| [29] | 高云超, 朱文珊, 陈文新. 秸秆覆盖免耕土壤细菌和真菌生物量与活性的研究. 生态学杂志, 2001, 20(2): 30-36. |
| [30] | Johnson C K, Wienhold B J, Doran J W, Drijber R A, Wright S F. Linking microbial-scale findings to farm-scale outcomes in a dryland cropping system. Precision Agriculture, 2004, 5(4): 311-328. |
| [31] | Muyzer G, Brinkhoff T, Nubel U, Santegoeds C, Schfer H, Wawer C. Denaturing gradient gel electrophoresis (DGGE) in microbial ecology//Kowalchuk G A, de Bruijn F J, Head I M, Akkermans A D, van Elsas J D. Molecular Microbial Ecology Manual. 2nd ed. 2004, 1(1/2): 743-769. |
| [32] | 董立国, 袁汉民, 李生宝, 袁海燕, 潘占兵. 玉米免耕秸秆覆盖对土壤微生物群落功能多样性的影响. 生态环境学报, 2010, 19(2): 444-446. |
| [33] | 秦华, 李国栋, 叶正钱, 徐秋芳, 曹志洪. 集约种植雷竹林土壤细菌群落结构的演变及其影响因素. 应用生态学报, 2010, 21(10): 2645-2651. |
| [34] | Jenkinson D S, Ladd J N. Microbial biomass in soil: measurement and turnover//Soil Biochemistry. New York: Marcel Dekker, 1981, (5): 451-471. |
| [35] | 张海燕, 张旭东, 李军, 王冬梅. 土壤微生物量测定方法概述. 微生物学杂志, 2005, 25(4): 95-99. |
| [36] | Konopka A, Oliver L, Turco R F Jr. The use of carbon substrate utilization patterns in environmental and ecological microbiology. Microbial Ecology, 1998, 35(2): 103-115. |
| [37] | 李春格, 李晓鸣, 王敬国. 大豆连作对土体和根际微生物群落功能的影响. 生态学报, 2006, 26(4): 1144-1150. |
| [38] | 胡元森, 吴坤, 刘娜, 陈红歌, 贾新成. 黄瓜不同生育期根际微生物区系变化研究. 中国农业科学, 2004, 37(10): 1521-1526. |
| [39] | 李建婷, 纪树兰, 刘志培, 秦振平, 刘缨, 杨媛媛. 16SrDNA克隆文库方法分析好氧颗粒污泥细菌组成. 环境科学研究, 2009, 22(10): 1218-1223. |
| [40] | 郑平, 张蕾. 厌氧氨氧化菌的特性与分类. 浙江大学学报: 农业与生命科学版, 2009, 35(5): 473-481. |
| [41] | 罗海峰, 齐鸿雁, 薛凯, 张洪勋. PCR-DGGE技术在农田土壤微生物多样性研究中的应用. 生态学报, 2003, 23(8): 1570-1575. |
| [42] | Su H M, Li X Y, He B H, Ouyang Y. Rainfall effects on soil microbial biomass and soil respiration under different land use conditions. Journal of Soil and Water Conservation, 2011, 25(6): 92-95. |
| [43] | 肖辉林, 郑习健. 土壤变暖对土壤微生物活性的影响. 土壤与环境, 2001, 10(2): 138-142. |
| [44] | 王立刚, 邱建军, 李维炯. 黄淮海平原地区夏玉米农田土壤呼吸的动态研究. 土壤肥料, 2002, (6): 13-17. |
| [45] | 张文佺, 王磊, 颜一青, 李艳丽, 王红丽, 贾建伟. 不同农业耕作模式下崇明盐碱土壤低碳化改良效应的模型评价. 农业环境科学学报, 2010, 29(5): 1006-1014. |
| [46] | 李会科, 梅立新, 高华. 黄土高原旱地苹果园生草对果园小气候的影响. 草地学报, 2009, 17(5): 615-620. |