 2015, Vol. 35
2015, Vol. 35文章信息
- 王玄, 江红星, 张亚楠
- WANG Xuan, JIANG Hongxing, ZHANG Yanan
- 稳定同位素分析在鸟类食性及营养级结构中的应用
- Application of stable isotope analyses to avian diets and trophic structure
- 生态学报, 2015, 35(16): 5556-5569
- Acta Ecologica Sinica, 2015, 35(16): 5556-5569
- http://dx.doi.org/10.5846/stxb201402120243
-
文章历史
- 收稿日期: 2014-02-12
- 修订日期: 2014-09-28






自然界中,食物、水和隐蔽物是动物生存的3大基本要素[1]。动物食性分析是了解动物与环境的相互关系,掌握其食物来源和营养级位置,以及开展资源管理与物种保护的前提和基础[2, 3]。传统的食性分析方法包括直接观察法[4]、嗉囊胃容物分析法[5, 6]、粪便显微分析法[7]等,均能直接反映一定时间尺度下动物的主要食物来源。随着人类认知能力和研究水平的提高,这些方法表现出一定的时空偶然性[8],不能充分反映食物在能量流动过程中的同化比例[9]。
稳定同位素技术自20世纪70年代引入生态学领域,在确定动物食物来源[10, 11]、构建食物链与食物网[12, 13, 14]以及追踪动物迁徙[15, 16, 17]等方面取得了重要进展。在动物食性研究方面,稳定同位素技术不仅克服了传统方法的局限性,还展示了其独特的优势:(1)可以反映动物与食物之间较长时间的取食关系[18, 19, 20];(2)可以连续测出动物在食物网中的营养位置,真实反映动物在生态系统中的位置和作用[21, 22];(3)可以通过对古化石的研究确定灭绝物种的食性信息[23, 24, 25];(4)还可以通过对濒临灭绝物种的稳定同位素的研究指导动物觅食栖息地的修复与重建[26, 27]。
鸟类是动物界中最为常见、易于观察的物种,通常作为环境质量监测的指示物种[28]。长期以来,鸟类学家一直关注鸟类食性研究。随着稳定同位素技术的发展,鸟类学家在食物来源和营养级研究中极大的丰富和完善了这一技术[9,18,29,30,31,32]。本文从稳定同位素技术原理、常见鸟类稳定同位素食性分析的样品类别、稳定同位素技术在鸟类食性组成和营养级位置研究中的应用等方面,对目前稳定同位素技术在鸟类食性研究中的最新进展进行总结,并提出有待解决的问题及今后的研究方向。
1 常用稳定同位素及其表达方式食性研究中常用的稳定同位素有碳(12C和13C)和氮(14N和15N) [18,29,33]。通常将机体的稳定性同位素的绝对含量与国际标准比对后进行研究,一般用δ13C(13C/12C)和δ15N(15N/14N)来分别表示机体内这两种稳定性同位素的丰度:
δX=[(R样/R标)-1]×103
式中,δX为δ13C或δ15N,R样为所测样品的13C/12C或15N/14N,R标为标准物质的13C/12C或15N/14N[34]。δ13C值是相对于国际标准物质PDB(R标=0.0112372)的丰度,δ15N值是相对于标准大气氮(R标=0.0036765)的丰度[35]。由于同一元素的不同同位素间原子量不同,导致其化学性质和物理性质存在差异,这种现象称为同位素效应(Isotope effect)[36, 37]。由于同位素效应的存在,较重的同位素更容易在机体内得到保留,而较轻的同位素更容易被机体代谢排出体外,稳定同位素的值在捕食者和食物之间产生了一个差值,这一差值称为营养级富集因子(Diet-tissue fractionation factors)[38, 39, 40, 41],若以“Δ”表示,则:
ΔX=δX样本-δX食物
式中,ΔX表示Δ13C或Δ15N,δX样本表示捕食者血液或组织的稳定同位素碳或氮的丰度值,δX食物表示食物中稳定同位素碳或氮的丰度。稳定同位素能够得到广泛应用依赖于以下2点:(1)稳定同位素的值在鸟类机体和食物之间是以一种可预测的方式进行传递;(2)稳定同位素值在鸟类机体不同部位转化速率不同,所以不同部位样本的稳定同位素值反映不同时间尺度的食性信息,如有迁徙或移动行为,则可反映不同空间尺度的食性信息[42]。
2 SIA的工作流程 2.1 确定研究目的,选择合适的稳定同位素受生理、技术等因素的影响,能够用于食性分析的稳定同位素种类并不多,稳定碳同位素(13C/12C)在营养级间传递时分馏效应不明显,富集因子较小,是用于食源组成研究的理想元素[43];稳定氮同位素(15N/14N)在食物与捕食者组织间存在稳定且较大的营养级富集因子,因此常被用于食源组成和营养级结构的分析[43, 44, 45];稳定硫同位素 (33S/32S)较少应用,但为提高分析准确性,稳定硫同位素在食源种类较多时也被使用[21,46]。此外,数据分析时一般要求各食源之间稳定同位素值存在显著差异,否则会低估取食较多的食源,而高估取食较少的食源[12,47]。
2.2 本底样品采集收集可能被研究对象取食的食物种类。为了减少样本分析的数据量、减小食物组成的种类和提高分析的精度,一般需要一些辅助手段,如直接观察法、胃内容物分析法、粪便显微分析法等,然后根据这些传统手段大致确定食物组成范围[12,47,48]。
2.3 鸟类研究样品的采集鸟类的稳定同位素分析样品众多,应根据研究目的选择合适样品,并尽量减少对水鸟造成的伤害,血浆一般反映短期摄入的食物特征,而羽毛可以反映在换羽地点(对候鸟也许是几千公里之外)的食物信息,常用的鸟类样品类别与采集方式见表1。
| 样品 Sample |
采集方式 Collection |
损伤程度 Damage |
实验难易程 Experiment |
代谢周期 Turnover rate |
| 羽毛Feather | 拣,拔,剪 | 轻微 | 易 | 长 |
| 全血Blood | 抽取 | 轻 | 中 | 短 |
| 血细胞Blood cell | 抽取,离心 | 轻 | 中 | 短 |
| 血浆Plasma | 抽取、离心 | 轻 | 中 | 短 |
| 肌肉Muscle | 解剖 | 重 | 中 | 中 |
| 内脏Visceral | 解剖 | 重 | 难 | 短 |
| 骨骼 Bone | 解剖 | 重 | 难 | 长 |
血液样本的采集依据鸟类体型大小,一般有翅静脉和颈静脉等取血方式,取血量1-2ml即可[48, 49]。采集的全血可通过离心分离出血细胞和血浆,获取新的研究样本[50, 51]。
鸟类的换羽有其各自的规律,应根据研究目的选择合适部位的羽毛,一般选择新长出的初级飞羽和尾羽。为避免取整根羽毛对鸟类飞行构成的影响,建议拣取刚脱落的羽毛或剪取所需部位羽毛末端2-4cm[42,52,53]。
肌肉组织含有一定量的脂肪,对稳定碳同位素值会产生较大影响,为减少实验误差,建议采集脂肪含量较少的胸部或大腿上的肌肉组织,1-2g即可达到测定需求[54, 55]。
用于稳定同位素分析的样品还包括:肝脏、肾脏、心脏、皮肤、大脑和骨骼等[49,50,51,55,56],由于对鸟类伤害较大,在食性研究中受到限制。
2.4 样品的保存样本完好的从野外保存至实验室是稳定同位素准确测定的前提,保存的原则是不因添加的保存物质而改变样品的稳定同位素比例,也不因保存方法问题导致样品的轻同位素或重同位素的比例发生变化[57]。血液、肌肉和羽毛等组织样本最常用保存方法是冷冻法,需在采样后2h内保存[58]。若野外无冷冻条件,且需保存时间较长,使用烘箱干燥、空气干燥、70%酒精浸泡等方法也可得到较好的保存效果[8]。样品保存中,冷冻保存不宜超过30d,70%酒精浸泡保存不宜超过60d[58]。对于羽毛,仅常温密封保存即可满足测定要求[59, 60]。
2.5 样品送检前的预处理 2.5.1 干燥干燥的目的是除去样品中的水分,便于研磨成粉。将样品放在透气性好且耐高温的器具或取样袋中,然后在60-70℃的恒温干燥箱中干燥24-48 h,至恒重后及时研磨成粉末,若样品返潮,不仅给磨样造成困难,且影响同位素数据[51]。对于血液,常采用冷冻干燥法[47,51,61]。
2.5.2 脱脂处理脱脂的目的是剔除肌肉、内脏和骨胶原等组织中的脂肪和羽毛上的油脂,因为脂肪的存在使样品中δ13C的值降低[42,62,63,64]。剔除方法是将样品放在装有氯仿和甲醇溶液(比例是2∶1)的索氏提取仪中加热煮沸20min,可用氢氧化钠代替氯仿[65]。因脱脂对稳定氮同位素的测定有负面影响,所以在脱脂前将样品分为两份,一份脱脂处理后测定稳定碳同位素含量,另一份不作脱脂处理,用于测定稳定氮同位素含量[51,62]。
2.5.3 酸化处理酸化的目的是除去样品中的无机碳,消除外源性无机碳对样品测定的影响,所有采集的样品均需要酸化[66]。将样品适当粉碎后,倒入适量浓度为0.5mol/L的盐酸中,每间隔1h用玻璃棒搅拌一次使之充分反应,反应至少6h后,静置沉淀,倒掉上层清夜,再用去离子水搅拌洗涤,静置沉淀,倾倒上层清夜,重复3-4次,充分洗净过量盐酸,然后烘干样品[26,38,44]。
2.5.4 充分研磨、过筛和保存烘干后的样品需粉碎才能进行分析,为了保证样品的均匀,应将样品充分研磨。粉碎可以用研钵、球磨机或混合磨碎机等处理,磨好的样品放于密封锡帽内或干净的离心管内,贴上标签,低温保存[67, 68]。
2.6 稳定同位素测定的技术支持同位素比率质谱仪(Isotope Ratio Mass Spectrometers,IRMS)是近些年发展起来的用于测定某些稳定同位素组成的连续流进样系统,由元素分析仪和质谱分析仪组成[69, 70]。与之前的双路进样系统相比,其特点是测试速度快,样品用量小(微克级),能精确测定元素的同位素比值[37]。
3 稳定同位素数据分析 3.1 营养级富集因子的确定营养级富集因子是准确确定鸟类食源组成和营养级位置的基本前提,为提高分析精度,测定的稳定同位素数据需经过富集因子矫正后输入分析模型[51,71]。实际研究中,营养级富集因子并非都是通过室内控制实验准确获取。大量研究结果表明献,同一科属的鸟类中生境类型、取食习性和个体大小相近的种类,营养级富集因子差异并不显著[40,51,72,73]。鉴于此,Caut等人于2009年[41]收集了66种期刊近300个物种的营养级富集因子,并按种类、生境和组织进行了分类归纳,表2显示鸟类各组织与食物之间的δ13C和δ15N的营养级富集因子的均值和标准误。该均值与Peterson和Fry[18]在1987年的结论相比更接近于真实值,并且得到了研究者的认可,在难以获取准确的营养级富集因子的情况下可采用该数据代替[41,48,74,75,76]。
| 组织Tissues | 营养级富集因子 Diet-tissue fractionation factors |
组织Tissues | 营养级富集因子 Diet-tissue fractionation factors |
||
| Δ 13C | Δ 15N | Δ 13C | Δ 15N | ||
| *表示血液中Δ13C与δ13C存在明显的线性关系;白色区域表示没有显著的线性关系,数据显示不同组织富集因子的均值(±SE);在不便获取研究对象准确的营养级富集因子,且无相关研究成果作为参考的情况下,可使用表中均值或公式近似的取代研究对象真实的营养级富集因子 | |||||
| 所有组织All tissues | 1.15 (0.18) | 2.91(0.16) | 骨胶原Collagen | 1.76 (0.64) | 2.82(0.14) |
| 羽毛Feather | 2.16 (0.35) | 2.25(0.20) | 肝脏Liver | 0.35(0.32) | 2.37(0.47) |
| 肌肉Muscle | 0.92(0.27) | 3.84(0.26) | 血浆Plasma | -0.08(0.38) | 2.60(0.38) |
| 血液Blood | -0.199δ 13C-3.986 * | 1.70(0.43) | |||
用于食源贡献率分析的数据模型有很多,但各有特点,应依据研究目的、同位素种类和数据特点选择合适的模型。常用稳定同位素分析的数据模型及其优缺点详见表3。
对2-3种食源的分析,可使用稳定同位素质量平衡方程(mass-balance mixing model)来实现。稳定同位素在时间尺度上是守恒的,且在不同营养级间以可预测的方式传递,利用质量平衡方程计算食物源的相对贡献比例既简单又快捷[77, 78, 79]:
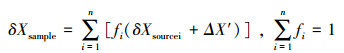
若只测定样本和食源的一种稳定同位素的相对含量(如δ13C),则可组成两个方程,可确定两种食物源的贡献:
δ13CSample=δ13CSource 1×f1+δ13CSource2×f2,f1+f2=1
式中,δSample为经营养级富集因子校正后的样本的稳定同位素值,δSource 1和δSource2为2种食源的稳定同位素值,f1和f2为这2种食源的贡献比例。如将测定的样本和食源的2种稳定同位素值(如δ13C和δ15N)代入质量平衡方程可确定3种食源的贡献,即测定样本和食源的n种稳定同位素可以确定n+1种食源的贡献率。当要解决更多食源问题(n>3种食物源) 时,则需要更为复杂的模型来计算每种食物源的贡献比例范围[80]。近年来,新的混合模型不断涌现,其中由Phillips等提出的线形混合模型是其中讨论较多的一种[81]。Phillips及其同事还开发出了一种相应的统计分析软件IsoSource (http://www.epa.gov/wed/pages/models/stableIsotopes/isosource/isosource.htm)。该软件适用于两种同位素下多种食源(3-10种)比例的确定,主要依据不同步长下的多次积分得到食物的相对比例。当有太多的食物资源而无法确定唯一的解时(n个同位素值却有>n+1种食源),可通过同位素比率信息来确定可能食物资源对混合物(消费者)的贡献范围。每种资源贡献范围(0-100%)的所有可能组合在小范围的增量(increments,如1%)中都被进行检查分析,把各个组合中观测到的混合同位素信息相加,如果之和在一个较小的忍受范围内(tolerance,如±0.01),那么这个组合就被认为是合适的解,从而可以确定潜在食物资源贡献的频率和范围[77,81,82,83]。
稳定同位素分析中可能存在很多不确定因素(如标准差、富集因子、同位素值的多变性等),这些不确定因素对数据的分析有很大影响。贝叶斯混合模型(Bayesian isotope mixing model)[86]通过综合先验信息,整合不确定因素,来明确比较竞争模型或参数值的支持强度。同时,该模型引入“取样-重要性-再取样(sampling-importance-resampling,SIR)”的运算法则,进行食源贡献率的后验概率检验,提高了数据分析的精度,是对IsoSource软件的丰富和发展[85]。考虑到复合食源、富集因子和同位素比值等的不确定性,模型运行过程中,可以通过调整变量数量、迭次量级和食源先验信息等来提高不同食源贡献率的估计精度。目前,该模型得到了广泛地应用,并取得了较好的效果[4,74,87,88,89,90]。可供选择的贝叶斯模型有MixSIA(http://conserver.iugo-cafe.org/user/brice.semmens/MixSIR)[85]和SIAR(http://cran.r-project.org/web /packages/siar/index.html)[87]。
3.3 确定营养级结构的数据分析方法一般情况下,稳定氮同位素(δ15N)在相邻两个营养级间所产生的富集因子(Δδ15N)在3‰-5‰之间,较稳定碳同位素产生的富集因子大且较为恒定[18]。根据相邻营养级间动物组织的δ15N和生态系统初级消费者(或初级生产者)的δ15N就可以划分动物在该生态系统的营养级位置:
TP=λ+δ15N消费者-δ15N基线/Δδ15N
式中,TP代表特定物种的营养位置,λ表示食物链基底生物的营养级位置(若δ15N基线取初级生产者的氮同位素比率,λ取1;若以初级消费者为基线,λ取2),δ15N消费者表示消费者的稳定氮同位素比率,δ15N基线表示基线生物的稳定氮同位素比值,Δδ15N是该生态系统的营养级富集因子[91]。δ15N基线和Δδ15N值随地理位置和生态系统(陆地或海洋生态系统)的改变而改变[91]。因δ13C值在相邻营养级间富集因子(Δδ13C)并不明显,仅为0.4‰-1.0‰,使δ13C值在动物营养级研究中不被广泛应用[91]。从营养级位置的计算公式可以看出,生态系统内基线生物的营养级富集因子对计算结果有一定影响。目前对基线营养级富集因子的获取方法有两种:一种是在室内严格控制的条件下,测量实验对象与其单一饵料间稳定氮同位素的差值;另一种方法是选用野外生态系统中食源相对简单的生物,测量其组织与食源间的稳定氮同位素的差值。室内实验控制了实验对象和饵料组成,但室内条件下生物的代谢活动、消化吸收率和饵料组成等均与其自然生态系统下存在较大差异,很难获取动物在自然状态下真实的营养级富集因子[92],针对这种情况,许多研究者采用基线营养级富集因子的统计平均值或几个统计平均值并用[93]。
4 SIA在鸟类食性及营养级中的研究现状 4.1 食源组成成分的确定动物取食食物后,通过代谢作用,机体的稳定性同位素δ13C值和δ15N值会因同化了不同同位素比率的食物而发生变化[38],稳定同位素比值在机体与组织间存在一个常量(即营养级富集因子),借助分析模型即可得到各食源的贡献比例[9,26,38]。稳定同位素技术是目前公认的用于研究迁徙鸟类食性时空变化的最佳手段[48, 94]。
4.1.1 不同时间尺度上的食性研究鸟类不同组织的代谢速度不同[95, 96],不同组织的稳定同位素分馏水平和转化周期也不尽相同,摄食后代谢较快的组织反映的是短时间尺度的食性信息,代谢较慢的组织反映的是长时间尺度的食性信息,这是稳定同位素技术在确定鸟类食源组成上区别于其他方法的最大优点[15,29,56,97]。
Hobson等[49,98]通过对人工饲养的日本鹌鹑(Coturnix japonic)血液和组织的稳定同位素研究发现,血清中稳定碳同位素值反映的是日本鹌鹑1周左右的食性信息,血细胞能够反映其两个月左右的食性信息,全血和脱脂后胸肌则能够反映其3周左右的食性信息,而代谢较慢的羽毛和骨胶原等反映的却是其几百天到几年的食性信息,在短嘴鸦(Corvus brachyrhyncho)[49]、黑腹滨鹬(Calidris alpina pacifica)[51]、黄腹鹪莺(Dendroica coronata)[60]和园莺(Sylvia borin)[99]等鸟类研究中均得到相似的转化规律。
鸟卵、雏鸟和正羽(飞羽和尾羽)是用于稳定性同位素时间特征分析的理想材料[52]。鸟卵的营养成分全部由产卵的雌鸟的食物成分转化而来,直接地反映了雌鸟在产卵繁殖期间的食性信息[100]。雏鸟的血液及组织完全由繁殖地所获得的食物转化而来,反映了雏鸟在育雏期的食性信息[11,48]。鸟类通常有换羽的习性,换羽方式可分为完全换羽和不完全换羽,初级飞羽、次级飞羽和尾羽的更换都有一定的规律,且更换的时间尺度也长短不一[101],因此,采集研究羽毛样本前须了解研究对象的换羽情况。Cherel等[52]在了解黑眉信天翁尾羽更换顺序后采集了尾羽最外侧和中间羽毛,分别代表了黑眉信天翁(Diomedea melanophrys)在冬羽换羽开始到换羽结束的食性信息。
4.1.2 不同空间尺度上的食性研究世界上现存鸟类有9755种[102],其中候鸟有5000多种[103],每年随季节的转变,候鸟在繁殖地和越冬地之间进行有规律的迁居,迁徙途中会选择补充能量的停歇地。繁殖地、越冬地和停歇地的食性反映了候鸟食性在空间尺度上的变化,对候鸟迁徙通道的保护有很大的作用,受到鸟类学家的关注[53]。
准时到达繁殖地是迁徙鸟类繁殖成功的重要保证,Marra等[104]在1995-1996年利用稳定碳同位素技术对美洲红尾鸥(Setophaga ruticilla)繁殖地和越冬地的食性进行研究,发现红尾鸥冬季栖息地食物资源非常有限,且越冬地的质量与春季迁回繁殖地的时间有密切关系。如果越冬地质量较差,红尾鸥到达繁殖地的时间就会提前或推迟,进而影响到它次年的繁殖活动。Minami等[105]利用稳定同位素技术对灰鹱(Puffinus griseus)和短尾鹱(P. tenuirostris)从南到北迁徙过程中的食性研究中发现,这两个物种的同位素组成随着他们从南半球到北太平洋的迁移而增加。且在日本北部的沿海海域灰鹱的δ13C和δ15N值均显著高于短尾鹱,然而在北太平洋海域两种鹱的δ15N并无显著差异,灰鹱的δ13C却显著高于短尾鹱。近海与远洋地理种群之间的稳定同位素的差异,反映出它们食性变化上的差异。
4.2 生态系统营养级结构分析稳定性碳、氮同位素技术被应用于分析生态系统中的营养级关系,是较为实用、准确和简单的一种方法,只需测定消费者组织和初级生产者的同位素比值,就可以推断它们在生态系统中的营养级位置。
4.2.1 确定研究对象在生态系统中的营养级位置稳定同位素技术在营养级结构研究中提出了新的划分方法,传统的营养级结构研究只能得到物种的整数点营养级,而新的营养级结构是连续的,这种表现方式更准确,因为研究对象在食物网中并非取食单一食物种类,杂食性鸟类既取食植物性食物又取食动物性食物,传统的营养级划分方法则不能准确的反映这一现象[80, 106]。这种新的营养级位置不仅可以反映各物种在系统中的营养位置和取食关系,而且也可以反映物种对物质的代谢水平以及环境因素差异所引起的营养状况和营养模式的改变[107,108, 109]。
Becker等[23]2006年通过对比加利福尼亚沿海岸100a前和现在斑海雀(Brachyramphus marmoratus)羽毛中的稳定碳氮同位素比值(δ13C和δ15N)发现,随着人类对沿海海洋生态系统渔业资源的过度捕捞,斑海雀食物资源显著降低,导致其在生态系统中的营养级位置显著下降。Procházka等[14]利用稳定同位素技术对非洲4种取食不同食物鸟类的营养级位置研究中发现,红嘴黑顶梅花雀(Estrilda nonnula)和查氏扇尾莺(Cisticola chubbi)的生态位较窄,而北非双领花蜜鸟(Cinnyris reichenowi)和西绿鹎(Andropadus tephrolaemus)却有较宽的取食生态位。
4.2.2 确定鸟类所在生态系统的营养级结构稳定同位素确定的营养级结构与传统方法相比较,能够准确反映能量沿营养级的流动和在物种之间的分配,间接地反映物种之间尤其是相近种之间取食生态位的分离和重叠程度[109]。在物种种内与种间关系的研究上都有很深远的意义[8, 110]。
Thompson等[111]通过稳定同位素分析,确定了冰岛上6种海鸟各自的食物组成和营养关系,Forero等[100]在对地中海地区海鸟的研究中建立了该地区海鸟及食源之间的营养级结构,系统阐述了稳定碳氮同位素在鸟类生态学研究中的广泛应用前景。杨月琴等[109]于2008年4-6月对青海湖的8种优势水鸟进行了稳定同位素分析,将其划分为3个明显的营养级群体,普通鸬鹚(Phalacrocorax carbo)、棕头鸥(Larus brunnicephalus)、渔鸥(L. ichthyaetus)和凤头(Podiceps cristatus)主要取食各种鱼类,稳定同位素值最高,属于最高的营养级群体;赤麻鸭(Tadorna ferruginea)、红脚鹬(Tringa totanus)和普通燕鸥(Sterna hirundo)取食鱼类、浮游动物和昆虫,稳定同位素值处在中间,属于第二营养级群体;而斑头雁(Anser indicus)主要取食植物性食物,稳定同位素值较低,属于最低的营养级群体。因此,通过该研究可以了解高低能量的食物在生态系统中的流动及分配情况。人类活动对野生动物的营养结构也产生了较大影响。易现峰等[112]在2002年4-7月间利用稳定同位素技术研究海北高寒草甸生态系统内主要生物的捕食关系,确定了高寒草甸生态系统中5条主要的食物链,21世纪初,由于当地居民大规模灭鼠,导致以田鼠为主要食物的大鵟(Buteo hemilasius)的食物结构发生了较大变化,主要食物来源由原来的小型哺乳类转变为雀形目鸟类。
5 SIA在鸟类保护与管理方面的应用稳定碳氮同位素分析不仅在鸟类食性及营养级的基础研究中起重要作用,而且在鸟类生态学的相关领域展现了强大发展潜力[68,113,114]。对于鸟类,尤其是濒危候鸟,其食性随着生境在时间和空间尺度上的变化而发生变化,利用稳定同位素技术结合相关研究手段可以从食性的角度,探究和解释食性与繁殖和种群动态之间的关系,进而从食性角度为物种保护和觅食生境恢复提供指导[24]。
5.1 评估栖息地变化对种群繁殖的影响栖息地(或生境)指动物生活的周围环境,对鸟类而言,栖息地就是个体、种群或群落在其某一生活史阶段(比如繁殖期、越冬期)所占据的环境类型,是其各种生命活动的场所[101]。然而,不断的资源开采和土地覆被的改变对鸟类栖息环境造成了较大的影响,其中食物资源减少和繁殖成功率降低是物种长久发展的最大障碍[101]。
稳定同位素技术结合相关研究手段能够合理的评估生境变化对种群繁殖的影响。Ramírez等[48]利用同位素和遥感技术,研究了自然和人工栖息地的变化对细嘴鸥(Chroicocephalus genei)种群大小和繁殖参数的影响。结果显示,在自然湿地减小的情况下,盐场作为细嘴鸥的替代栖息地,扮演了越来越重要的角色;而渔场等人工湿地的建立,对细嘴鸥自然栖息地丧失起到了一定的缓冲作用,但食源结构的改变降低了其繁殖产出。Ma等[74]于2007-2008年利用同位素技术对上海崇明东滩外来物种互花米草(Spartina alterniflora)群落中最新记录到的斑背大尾莺(Locustella pryeri)的食性、栖息地利用和繁殖成功率进行了研究。结果表明,在芦苇和互花米草混合群落中,斑背大尾莺主要营巢和取食于互花米草;其种群数量也随着互花米草的快速扩展而增加。这说明互花米草的入侵,不仅改变了原生盐沼湿地生态系统,同时促进了非本土物种斑背大尾莺的集聚与繁衍。
5.2 探究种群数量波动的原因人类对自然资源的过度索取使许多鸟类丧失栖息地甚至发生食物链的断裂,最终导致其种群数量锐减或灭绝[115],然而一些鸟类在短期内也有种群爆发的现象[63, 116]。探究种群数量波动的原因对鸟类保护计划的制定以及保护区管理都有指导意义[24]。
20世纪20年代到70年代加拿大西海岸的鸥类种群数量翻了3.5倍[116],Hobson[63]采集了灰翅鸥(L. glaucescens)和西美鸥(L. occidentalis)的骨胶原蛋白,同位素分析表明,灰翅鸥和西美鸥食物的主要来源于人类丢弃的可食废弃物,从食性角度解释了其种群爆发的原因。Thompson等[117]通过同位素分析,认为暴风鹱(Fulmarus glacialis)在英国上世纪种群数量的爆发与捕鲸业的发展有关,因为当时暴风鹱主要取食捕鲸遗弃的内脏。然而上世纪在欧洲种群数量同样增长的普通鸬鹚却对淡水渔业产生了较大的冲击,同位素研究表明,普通鸬鹚种群增长的原因是其食源主要依赖淡水渔业[118]。
稳定同位素技术还为重现灭绝鸟类的食性提供了可能,生活在北大西洋沿岸的大海雀(Pinguinus impennis)20世纪中叶灭绝了,Hobson等[119]挖掘大海雀曾生活过的海岛上的骨骼样本,通过分析其中的稳定氮同位素(δ15N)值,并与该生态系统中其他6种海雀的稳定同位素相比较,最终重现其食性信息和在生态系统中的营养级位置。Norris等[24]通过同位素技术研究了斑海雀(Brachyramphus marmoratus)过去100a的食性组成的变化和种群动态之间的关系,认为食物的质量和食物的可获得性是种群动态变化的主要原因。通过历史食性和种群波动的研究,能为濒危鸟类保护计划的制定提供更好的参考价值。
5.3 评估越冬地和繁殖地对繁殖的能量贡献鸟类繁殖营养学是稳定同位素技术的比较新的研究领域。对于迁徙鸟类,其繁殖的能量通常有内源性和外源性能量两部分组成[43, 76],内源性能量表示候鸟在到达繁殖地之前(即在越冬地)所积累的能量,外源性能量则表示候鸟到达繁殖地后于繁殖地短期快速获取的能量。通过稳定碳氮同位素(δ13C和δ15N)对繁殖能量的示踪,可以准确评估越冬地的生境质量对繁殖的贡献情况,为越冬地和种群的保护提供科学依据[43,120,121,122]。
传统研究认为内源性能量对恶劣环境下繁殖的大型鸟类贡献率大[123, 124]。Gauthier等[123]对雪雁(Chen caerulescens atlantica)的肌肉组织、卵和繁殖地食物的稳定碳同位素进行了分析,由于雪雁在越冬区主要取食玉米(C4植物),而到达繁殖区后主要食物是禾本科与非禾本科的杂草(C3植物),因此内源性能量和外源性能量之间的稳定碳同位素(δ13C)存在显著差异。同位素数据显示,内源性的能量对卵的贡献比外源性的能量贡献率低,从同位素的角度证实了传统观念的局限。然而,通常情况下内源性和外源性的能量对繁殖卵的贡献是复杂的。Sénéchal等[124]于2010年研究了极地繁殖的欧绒鸭(Somateria mollissima)内源性和外源性营养对鸟卵的贡献情况,结果显示,外源性能量对卵的蛋白贡献较大,内源性能量对卵黄的脂类贡献较大。Hobson等[125]对巴氏鹊鸭(Bucephala islandica)的研究表明,内源性能量对第1每卵的卵黄和蛋白的贡献显著高于第4和第8枚卵,然而5a前获取的样本显示,内源性营养对卵黄的贡献并不明显。他们认为巴氏鹊鸭内源性和外源性能量对繁殖的贡献表现为一种混合的策略,因不同个体、产卵策略和年份等差异而变化。
6 SIA在鸟类食性研究中的注意事项与展望稳定同位素技术是目前揭示鸟类食源,探究湿地生态系统内物质循环、能量流动和营养级结构等的重要方法。但是在食源组成分析和迁徙鸟类食性转变的研究中还有很多需要思考的问题,这些问题也许会给我们提供一些新的研究和发展方向。
6.1 稳定同位素在机体内的分馏及影响因素稳定同位素技术在应用时需充分考虑引起稳定同位素在机体内分馏的因素,如成鸟与幼鸟的差异、雌雄差异、繁殖鸟与非繁殖鸟差异、雌雄二型与雌雄同型的差异以及温度对分馏的影响等。Shealer[126] 研究粉红燕鸥(Sterna dougallii)和黄嘴凤头燕鸥(Thalasseus sandvicensis)发现,幼鸟因飞行能力有限,难以吞咽较大块食物等因素的影响,所取食的食物与成鸟有很大差异。Niebuhr[127] 在繁殖期研究银鸥(L. argentatus)哺育幼鸟的成鸟比非繁殖成鸟获取高能量的食物,在产卵期雌鸟为了促进蛋壳的形成,会偏向于取食贝类等。Lavoie等[4]研究加拿大海鸟食性发现,二形的雄性成体要比雌性成体的稳定性同位素变化幅度大;相反,单形的雌性成体的同位素变化幅度比雄性成体大。有研究表明,迁徙鸟类和伤残病鸟等未及时获取水分(水分胁迫)和足够食物(营养胁迫)时,稳定氮同位素值会发生很大变化。受水分胁迫时,浓缩的含氮废物中氮同位素的分馏比稀释的含氮废物中的高,受营养胁迫时,机体蛋白质的分解代谢提高了15N值,因此,在样本采集时为提高实验准确性,应避免采集受水分胁迫或营养胁迫的个体[51]。
6.2 营养级富集因子的获取及意义虽然稳定同位素在机体各组织内富集规律相同,甚至可使用其他物种的富集因子均值代替研究对象的营养级富集因子,但不同的物种、不同的采样部位和不同栖息环境下稳定同位素的值都存在差异[8,72,128,129,130]。因此,需要更多的室内试验来得到具体动物种类的特定营养级富集因子,以更好地解释野外实验获得的稳定同位素数据,从而更好地估算该种类的食源组成[98]。然而,被捕的鸟类被关进一个封闭的笼内,其活动空间较野外自由状态下小很多,因此其代谢速度也较低,那么稳定同位素在体内的转化速率同样会降低,这与野外生活鸟类的真实稳定同位素值存在一定差异,但是在野生状态下无法确保同位素含量稳定的食物和持续供给,因此,通过被捕鸟类的稳定同位素研究,可提供一种最接近富集因子真实状态的近似值[51]。
6.3 稳定同位素在机体内的转化周期稳定同位素在机体内含量的高低受机体代谢速率的影响,且不同组织的代谢速率不同,通过采集不同代谢速率的组织样本反映不同时间尺度的食性信息。值得注意的是,迁徙中的鸟类代谢更旺盛,稳定同位素的转化周期则更短[42,60];由于实验中的转化周期都是通过对捕捉鸟类的研究得出的,鸟类在野外的代谢速率可能比捕捉后饲养条件下的高[98];此外,血液中分离出的各成分比例与鸟的种类和体内的激素水平有关,对稳定同位素的含量和转化周期都有影响[95,131]。另外,通过稳定同位素的转化周期可以来解释一些与机体代谢相关的其他生理反应。
6.4 同位素印记对食源分析的影响同位素印记是指不同同位素比值的食物在进入动物组织时,会分别进入动物特定的组织或部位,并非平均分配到动物不同组织或组织的不同部分[99,132]。虽然同位素印记现象还没有得到更多研究的确认,但已有的研究表明同位素印记现象确实存在[99,132,133],其结果是动物某一组织并不能完全反映其整体的食物组成[72]。这一问题的提出对利用稳定同位素分析鸟类食性的准确性提出了质疑,也将影响稳定同位素的长远发展,为解决这一难题,需要更深入的研究稳定同位素在动物组织与其食物成分之间的转化关系,特别是当杂食性动物食性发生转变时,动物组织同位素组成的动态平衡过程又是怎样的变化特征[72,134]。
作为一门新技术,稳定同位素技术已向研究者展示了强大的活力,但在应用中仍然需要注意一些细节。不断的对未知生物体内碳氮稳定同位素富集因子进行探索和总结,也许能发现一些新的规律。稳定碳氮同位素的应用领域还有很大的发展空间,可以去解释一些鸟类生理上的问题,也可与其他技术手段相结合解释一些更为复杂的问题,如探讨动物种间种内竞争、协同进化、污染生态学、生态保护与恢复等等。此外,由于在地表降水中δD(2H/1H)和δ18O(18O/16O)与纬度、海拔高度以及距海岸线距离等有密切关系,因此δD和δ18O作为地理位置的标记被鸟类学家广泛应用于迁徙研究中[59,104,135,136,137]。随着技术手段的不断进步和越来越多的鸟类学家的加入,稳定同位素分析在鸟类生态学研究中一定会得到长足的发展。
| [1] | 陈化鹏, 高中信. 野生动物生态学. 哈尔滨: 东北林业大学出版社, 1992. |
| [2] | 郑光美. 我国鸟类生态学的回顾与展望. 动物学杂志, 1981, (1): 63-68. |
| [3] | 郑荣泉, 鲍毅新. 有蹄类食性研究方法及研究进展. 生态学报, 2004, 24(7): 1532-1539. |
| [4] | Lavoie R A, Rail J F, Lean D R S. Diet composition of seabirds from Corossol island, Canada, using direct dietary and stable isotope analyses. Waterbirds, 2012, 35(3): 402-419. |
| [5] | Jobling M, Breiby A. The use and abuse of fish otoliths in studies of feeding habits of marine piscivores. Sarsia, 1986, 71(3): 265-274. |
| [6] | Ryan P G, Jackson S. Stomach pumping: is killing seabirds necessary? The Auk, 1986, 103(2): 427-428. |
| [7] | Hobbs N T. Fecal indices to dietary quality: a critique. The Journal of Wildlife Management, 1987, 51(2): 317-320. |
| [8] | Hobson K A, Piatt J F, Pitocchelli J. Using stable isotopes to determine seabird trophic relationships. Journal of Animal Ecology, 1994, 63(4): 786-798. |
| [9] | Tieszen L L, Boutton T W, Tesdahl K, Slade N A. Fractionation and turnover of stable carbon isotopes in animal tissues: implications for δ13C analysis of diet. Oecologia, 1983, 57(1): 32-37. |
| [10] | Estrada J, Lutcavage M, Thorrold S. Diet and trophic position of Atlantic bluefin tuna (Thunnus thynnus) inferred from stable carbon and nitrogen isotope analysis. Marine Biology, 2005, 147(1): 37-45. |
| [11] | Girard J, Baril A, Mineau P, Fahrig L. Foraging habitat and diet of Song Sparrows (Melospiza melodia) nesting in farmland: a stable isotope approach. Canadian Journal of Zoology, 2012, 90(11): 1339-1350. |
| [12] | 易现峰, 张晓爱. 稳定性同位素技术在生态学上的应用. 生态学杂志, 2005, 24(3): 306-314. |
| [13] | Hobson K A. Trophic relationships among high Arctic seabirds: insights from tissue-dependent stable-isotope models. Marine Ecology Progress Series, 1993, 95: 7-18. |
| [14] | Procházka P, Reif J, Hořák D, Klvaňa P, Lee R W, Yohannes E. Using stable isotopes to trace resource acquisition and trophic position in four Afrotropical birds with different diets. Ostrich, 2010, 81(3): 273-275. |
| [15] | Hobson K A, Wassenaar L I. Linking breeding and wintering grounds of neotropical migrant songbirds using stable hydrogen isotopic analysis of feathers. Oecologia, 1996, 109(1): 142-148. |
| [16] | Hobson K A. Tracing diets and origins of migratory birds using stable isotope techniques Biology and Conservation of Forest Birds, 1999, 1: 21-41. |
| [17] | Ashley P, Hobson K, Van Wilgenburg S, North N, Petrie S. Linking Canadian harvested American black ducks to their natal areas using stable isotope analysis of δD, δ13C, and δ15N. Avian Conservation and Ecology, 2010, 5(2): 7. |
| [18] | Peterson B J, Fry B. Stable isotopes in ecosystem studies. Annual Review of Ecology and Systematics, 1987, 18: 293-320. |
| [19] | 蔡德陵, 张淑芳, 张经. 天然存在的碳、氮稳定同位素在生态系统研究中的应用. 质谱学报, 2003, 24(3): 434-440. |
| [20] | Hobson K, Van Wilgenburg S, Wassenaar L, Powell R, Still C, Craine J. A multi-isotope (δ13C, δ15N, δ2H) feather isoscape to assign Afrotropical migrant birds to origins. Ecosphere, 2012, 3(5): 44-44. |
| [21] | 万祎, 胡建英, 安立会, 安伟, 杨敏, 伊藤光明, 服部达也, 陶澍. 利用稳定氮和碳同位素分析渤海湾食物网主要生物种的营养层次. 科学通报, 2005, 50(7): 708-712. |
| [22] | Boecklen W J, Yarnes C T, Cook B A, James A C. On the use of stable isotopes in trophic ecology. Annual Review of Ecology, Evolution, and Systematics, 2011, 42: 411-440. |
| [23] | Becker B H, Beissinger S R. Centennial decline in the trophic level of an endangered seabird after fisheries decline. Conservation Biology, 2006, 20(2): 470-479. |
| [24] | Norris D, Arcese P, Preikshot D, Bertram D F, Kyser T. Diet reconstruction and historic population dynamics in a threatened seabird. Journal of Applied Ecology, 2007, 44(4): 875-884. |
| [25] | Touzeau A, Amiot R, Blichert-Toft J, Flandrois J P, Fourel F, Grossi V, Martineau F, Richardin P, Lécuyer C. Diet of ancient Egyptians inferred from stable isotope systematics. Journal of Archaeological Science, 2014, 46: 114-124. |
| [26] | Barrett R T, Camphuysen K C, Anker-Nilssen T, Chardine J W, Furness R W, Garthe S, Hüppop O, Leopold M F, Montevecchi W A, Veit R R. Diet studies of seabirds: a review and recommendations. ICES Journal of Marine Science: Journal du Conseil, 2007, 64(9): 1675-1691. |
| [27] | Ramírez F, Abdennadher A, Sanpera C, Jover L, Wassenaar L I, Hobson K A. Assessing waterbird habitat use in coastal evaporative systems using stable isotopes (δ13C, δ15N and δD) as environmental tracers. Estuarine, Coastal and Shelf Science, 2011, 92(2): 217-222. |
| [28] | 王强, 吕宪国. 鸟类在湿地生态系统监测与评价中的应用. 湿地科学, 2007, 5(3): 274-281. |
| [29] | DeNiro M J, Epstein S. Influence of diet on the distribution of carbon isotopes in animals. Geochimica et Cosmochimica Acta, 1978, 42(5): 495-506. |
| [30] | Hobson K A, Welch H E. Determination of trophic relationships within a high Arctic marine food web using δ13C and δ15N analysis. Marine Ecology Progress Series, 1992, 84: 9-18. |
| [31] | Phillips R A, McGill R A, Dawson D A, Bearhop S. Sexual segregation in distribution, diet and trophic level of seabirds: insights from stable isotope analysis. Marine Biology, 2011, 158(10): 2199-2208. |
| [32] | Van-Wilgenburg S L, Hobson K A, Brewster K R, Welker J M. Assessing dispersal in threatened migratory birds using stable hydrogen isotope (δD) analysis of feathers. Endangered Species Research, 2012, 16(1): 17-29. |
| [33] | DeNiro M J, Epstein S. Influence of diet on the distribution of nitrogen isotopes in animals. Geochimica et Cosmochimica Acta, 1981, 45(3): 341-351. |
| [34] | McKinney C R, McCrea J M, Epstein S, Allen H, Urey H C. Improvements in mass spectrometers for the measurement of small differences in isotope abundance ratios. Review of Scientific Instruments, 1950, 21(8): 724-730. |
| [35] | Fry B. Stable Isotope Ecology. New York: Springer, 2006: 1-308. |
| [36] | Farquhar G D, Ehleringer J R, Hubick K T. Carbon isotope discrimination and photosynthesis. Annual Review of Plant Biology, 1989, 40(1): 503-537. |
| [37] | Hoefs J. Stable Isotope Geochemistry. 6th ed. Berlin, Heidelberg: Springer, 2009: 1-292. |
| [38] | Duffy D C, Jackson S. Diet studies of seabirds: a review of methods. Colonial Waterbirds, 1986, 9: 1-17. |
| [39] | Therrien J F, Fitzgerald G, Gauthier G, Bêty J. Diet-tissue discrimination factors of carbon and nitrogen stable isotopes in blood of Snowy Owl (Bubo scandiacus). Canadian Journal of Zoology, 2011, 89(4): 343-347. |
| [40] | Caut S, Angulo E, Courchamp F. Discrimination factors (δ15N and δ13C) in an omnivorous consumer: effect of diet isotopic ratio. Functional Ecology, 2008, 22(2): 255-263. |
| [41] | Caut S, Angulo E, Courchamp F. Variation in discrimination factors (δ15N and δ13C): the effect of diet isotopic values and applications for diet reconstruction. Journal of Applied Ecology, 2009, 46(2): 443-453. |
| [42] | Bearhop S, Waldron S, Votier S C, Furness R W. Factors that influence assimilation rates and fractionation of nitrogen and carbon stable isotopes in avian blood and feathers. Physiological and Biochemical Zoology, 2002, 75(5): 451-458. |
| [43] | Inger R, Bearhop S. Applications of stable isotope analyses to avian ecology. Ibis, 2008, 150(3): 447-461. |
| [44] | Kelly J F. Stable isotopes of carbon and nitrogen in the study of avian and mammalian trophic ecology. Canadian Journal of Zoology, 2000, 78(1): 1-27. |
| [45] | Bond A L, Jones I. A practical introduction to stable-isotope analysis for seabird biologists: approaches, cautions and caveats. Mar Ornithol, 2009, 37(3): 183-188. |
| [46] | Nehlich O, Fuller B T, Márquez-Grant N, Richards M P. Investigation of diachronic dietary patterns on the islands of Ibiza and Formentera, Spain: evidence from sulfur stable isotope ratio analysis. American Journal of Physical Anthropology, 2012, 149(1): 115-124. |
| [47] | 张璇, 华宁, 汤臣栋, 马强, 薛文杰, 吴魏, 马志军. 崇明东滩黑腹滨鹬(Calidris alpina)食物来源和组成的稳定同位素分析. 复旦学报: 自然科学版, 2013, 52(1): 112-118. |
| [48] | Ramírez F, Navarro J, Afán I, Hobson K A, Delgado A, Forero M G. Adapting to a changing world: unraveling the role of man-made habitats as alternative feeding areas for slender-billed gull (Chroicocephalus genei). PloS One, 2012, 7(10): e47551. |
| [49] | Hobson K A, Clark R. Turnover of 13C in cellular and plasma fractions of blood: implications for nondestructive sampling in avian dietary studies. The Auk, 1993, 110(3): 638-641. |
| [50] | Barquete V, Strauss V, Ryan P G. Stable isotope turnover in blood and claws: A case study in captive African Penguins. Journal of Experimental Marine Biology and Ecology, 2013, 448: 121-127. |
| [51] | Ogden L J E, Hobson K A, Lank D B, Martínez del Rio C. Blood isotopic (δ13C and δ15N) turnover and diet-tissue fractionation factors in captive dunlin (Calidris alpina pacifica). The Auk, 2004, 121(1): 170-177. |
| [52] | Cherel Y, Hobson K A, Weimerskirch H. Using stable-isotope analysis of feathers to distinguish moulting and breeding origins of seabirds. Oecologia, 2000, 122(2): 155-162. |
| [53] | Hobson K A, Bowen G J, Wassenaar L I, Ferrand Y, Lormee H. Using stable hydrogen and oxygen isotope measurements of feathers to infer geographical origins of migrating European birds. Oecologia, 2004, 141(3): 477-488. |
| [54] | Bosley K L, Wainright S C. Effects of preservatives and acidification on the stable isotope ratios (15N/14N, 13C/12C) of two species of marine animals. Canadian Journal of Fisheries and Aquatic Sciences, 1999, 56(11): 2181-2185. |
| [55] | Schmutz J A, Hobson K A. Geographic, temporal, and age-specific variation in diets of glaucous gulls in western Alaska. Condor, 1998, 100(1): 119-130. |
| [56] | Hobson K A, Clark R G. Assessing avian diets using stable isotopes I: turnover of 13C in tissues. Condor, 1992, 94(1): 181-188. |
| [57] | Gearing J N. The study of diet and trophic relationships through natural abundance 13C//Coleman D C, Fry B. Carbon Isotope Techniques. San Diego: Academic Press, 1991: 201-218. |
| [58] | Barrow L M, Bjorndal K A, Reich K J. Effects of preservation method on stable carbon and nitrogen isotope values. Physiological and Biochemical Zoology, 2008, 81(5): 688-693. |
| [59] | Van Dijk J G, Meissner W, Klaassen M. Improving provenance studies in migratory birds when using feather hydrogen stable isotopes. Journal of Avian Biology, 2014, 45(1): 103-108. |
| [60] | Podlesak D W, McWilliams S R, Hatch K A. Stable isotopes in breath, blood, feces and feathers can indicate intra-individual changes in the diet of migratory songbirds. Oecologia, 2005, 142(4): 501-510. |
| [61] | Hobson K A, McLellan B N, Woods J G. Using stable carbon (δ13C) and nitrogen (δ15N) isotopes to infer trophic relationships among black and grizzly bears in the upper Columbia River basin, British Columbia. Canadian Journal of Zoology, 2000, 78(8): 1332-1339. |
| [62] | DeNiro M J, Epstein S. Mechanism of carbon isotope fractionation associated with lipid synthesis. Science, 1977, 197(4300): 261-263. |
| [63] | Hobson K A. Use of stable-carbon isotope analysis to estimate marine and terrestrial protein content in gull diets. Canadian Journal of Zoology, 1987, 65(5): 1210-1213. |
| [64] | Post D M, Layman C A, Arrington D A, Takimoto G, Quattrochi J, Montana C G. Getting to the fat of the matter: models, methods and assumptions for dealing with lipids in stable isotope analyses. Oecologia, 2007, 152(1): 179-189. |
| [65] | Ambrose S H. Isotopic analysis of paleodiets: methodological and interpretive considerations. Investigations of Ancient Human Tissue, 1993, 10: 59-130. |
| [66] | Jaschinski S, Hansen T, Sommer U. Effects of acidification in multiple stable isotope analyses. Limnol Oceanogr: Methods, 2008, 6: 12-15. |
| [67] | Oppel S, Federer R N, O'Brien D M, Powell A N, Hollmén T E. Effects of lipid extraction on stable isotope ratios in avian egg yolk: is arithmetic correction a reliable alternative? The Auk, 2010, 127(1): 72-78. |
| [68] | Doucette J L, Wissel B, Somers C M. Cormorant-fisheries conflicts: stable isotopes reveal a consistent niche for avian piscivores in diverse food webs. Ecological Applications, 2011, 21(8): 2987-3001. |
| [69] | Brand W A. Mass spectrometer hardware for analyzing stable isotope ratios. Handbook of Stable Isotope Analytical Techniques, 2004, 1: 835-857. |
| [70] | Werner R A, Brand W A. Referencing strategies and techniques in stable isotope ratio analysis. Rapid Communications in Mass Spectrometry, 2001, 15(7): 501-519. |
| [71] | Auerswald K, Wittmer M H, Zazzo A, Schäufele R, Schnyder H. Biases in the analysis of stable isotope discrimination in food webs. Journal of Applied Ecology, 2010, 47(4): 936-941. |
| [72] | Martínez del Rio C, Wolf N, Carleton S A, Gannes L Z. Isotopic ecology ten years after a call for more laboratory experiments. Biological Reviews, 2009, 84(1): 91-111. |
| [73] | Caut S, Angulo E, Courchamp F. Caution on isotopic model use for analyses of consumer diet. Canadian Journal of Zoology, 2008, 86(5): 438-445. |
| [74] | Ma Z, Gan X, Choi C Y, Li B. Effects of invasive cordgrass on presence of marsh grassbird in an area where it is not native. Conservation Biology, 2013, 28(1): 150-158. |
| [75] | Cherry S G, Derocher A E, Hobson K A, Stirling I, Thiemann G W. Quantifying dietary pathways of proteins and lipids to tissues of a marine predator. Journal of Applied Ecology, 2011, 48(2): 373-381. |
| [76] | Hobson K A. Isotopic ornithology: a perspective. Journal of Ornithology, 2011, 152(1): 49-66. |
| [77] | Phillips D L, Gregg J W. Uncertainty in source partitioning using stable isotopes. Oecologia, 2001, 127(2): 171-179. |
| [78] | Saito L, Johnson B M, Bartholow J, Hanna R B. Assessing ecosystem effects of reservoir operations using food web-energy transfer and water quality models. Ecosystems, 2001, 4(2): 105-125. |
| [79] | Del Rio C M, Wolf B O. Mass-balance models for animal isotopic ecology//Starck J M, Wang T. Physiological and Ecological Adaptations to Feeding in Vertebrates. Enfield, New Hampshire: Science Publishers, 2005: 141-174. |
| [80] | 冯建祥, 黄敏参, 黄茜, 郭婕敏, 林光辉. 稳定同位素在滨海湿地生态系统研究中的应用现状与前景. 生态学杂志, 2013, 32(4): 1065-1074. |
| [81] | Phillips D L, Gregg J W. Source partitioning using stable isotopes: coping with too many sources. Oecologia, 2003, 136(2): 261-269. |
| [82] | Phillips D L. Mixing models in analyses of diet using multiple stable isotopes: a critique. Oecologia, 2001, 127(2): 166-170. |
| [83] | Phillips D L, Newsome S D, Gregg J W. Combining sources in stable isotope mixing models: alternative methods. Oecologia, 2005, 144(4): 520-527. |
| [84] | Moreno R, Jover L, Munilla I, Velando A, Sanpera C. A three-isotope approach to disentangling the diet of a generalist consumer: the yellow-legged gull in northwest Spain. Marine Biology, 2010, 157(3): 545-553. |
| [85] | Moore J W, Semmens B X. Incorporating uncertainty and prior information into stable isotope mixing models. Ecology Letters, 2008, 11(5): 470-480. |
| [86] | Jackson A L, Inger R, Bearhop S, Parnell A. Erroneous behaviour of MixSIR, a recently published Bayesian isotope mixing model: a discussion of Moore & Semmens (2008). Ecology Letters, 2009, 12(3): e1-e5. |
| [87] | Parnell A C, Inger R, Bearhop S, Jackson A L. Source partitioning using stable isotopes: coping with too much variation. PloS One, 2010, 5(3): e9672. |
| [88] | Semmens B X, Ward E J, Parnell A C, Phillips D L, Bearhop S, Inger R, Jackson A, Moore J W. Statistical basis and outputs of stable isotope mixing models: Comment on Fry (2013). Marine Ecology Progress Series, 2013, 490: 285-289. |
| [89] | Bond A L, Diamond A W. Recent Bayesian stable-isotope mixing models are highly sensitive to variation in discrimination factors. Ecological Applications, 2011, 21(4): 1017-1023. |
| [90] | Weiser E L, Powell A N. Evaluating gull diets: a comparison of conventional methods and stable isotope analysis. Journal of Field Ornithology, 2011, 82(3): 297-310. |
| [91] | Post D M. Using stable isotopes to estimate trophic position: models, methods, and assumptions. Ecology, 2002, 83(3): 703-718. |
| [92] | France R, Peters R. Ecosystem differences in the trophic enrichment of 13C in aquatic food webs. Canadian Journal of Fisheries and Aquatic Sciences, 1997, 54(6): 1255-1258. |
| [93] | 林光辉. 稳定同位素生态学. 北京: 高等教育出版社, 2013. |
| [94] | Ramos R, Ramírez F, Carrasco J L, Jover L. Insights into the spatiotemporal component of feeding ecology: an isotopic approach for conservation management sciences. Diversity and Distributions, 2011, 17(2): 338-349. |
| [95] | Waterlow J C, Garlick P J, Mill Ward D. Protein Turnover in Mammalian Tissues and in the Whole Body. Amsterdam: North-Holland Biomedical Press, 1978. |
| [96] | Schoenheimer R. The Dynamic State of Body Constituents. Cambridge: Harvard University Press, 1946. |
| [97] | Hilderbrand G V, Farley S, Robbins C, Hanley T, Titus K, Servheen C. Use of stable isotopes to determine diets of living and extinct bears. Canadian Journal of Zoology, 1996, 74(11): 2080-2088. |
| [98] | Hobson K A, Clark R G. Assessing avian diets using stable isotopes II: factors influencing diet-tissue fractionation. Condor, 1992, 94(1): 189-197. |
| [99] | Hobson K A, Bairlein F. Isotopic fractionation and turnover in captive Garden Warblers (Sylvia borin): implications for delineating dietary and migratory associations in wild passerines. Canadian Journal of Zoology, 2003, 81(9): 1630-1635. |
| [100] | Forero M G, Hobson K A. Using stable isotopes of nitrogen and carbon to study seabird ecology: applications in the Mediterranean seabird community. Scientia Marina, 2003, 67(S2): 23-32. |
| [101] | 郑光美. 鸟类学 (第二版). 北京: 北京师范大学出版社, 2012. |
| [102] | 郑光美. 世界鸟类分类与分布名录. 北京: 科学出版社, 2002. |
| [103] | Gretel H S, Sheila K S. Animal Behavior: Animal Migration. Chelsea House, 2009: 1-120. |
| [104] | Marra P P, Hobson K A, Holmes R T. Linking winter and summer events in a migratory bird by using stable-carbon isotopes. Science, 1998, 282(5395): 1884-1886. |
| [105] | Minami H, Minagawa M, Ogi H. Changes in stable carbon and nitrogen isotope ratios in sooty and short-tailed shearwaters during their northward migration. Condor, 1995, 97(2): 565-574. |
| [106] | 蔡德陵, 李红燕, 唐启升, 孙耀. 黄东海生态系统食物网连续营养谱的建立: 来自碳氮稳定同位素方法的结果. 中国科学: C辑, 2005, 35(2): 123-130. |
| [107] | Wada E, Terazaki M, Kabaya Y, Nemoto T. 15N and13C abundances in the Antartic Ocean with emphasis on the biogeochemical structure of the food web. Deep Sea Research Part A. Oceanographic Research Papers, 1987, 34(5): 829-841. |
| [108] | Layman C A, Arrington D A, Montaña C G, Post D M.Can stable isotope ratios provide for community-wide measures of trophic structure? Ecology, 2007, 88(1): 42-48. |
| [109] | 杨月琴, 易现峰, 李宁. 利用稳定同位素技术分析青海湖优势水鸟的营养级结构. 动物学研究, 2009, 30(4): 418-422. |
| [110] | Thompson D R, Furness R W. Stable-isotope ratios of carbon and nitrogen in feathers indicate seasonal dietary shifts in northern fulmars. The Auk, 1995, 112(2): 493-498. |
| [111] | Thompson D R, Lilliendahl K, Sólmundsson J, Furness R W, Waldron S, Phillips R A. Trophic relationships among six species of Icelandic seabirds as determined through stable isotope analysis. Condor, 1999, 101: 898-903. |
| [112] | Yi X F, Li L X, Zhang X A, Li M C. Diet shift of upland buzzards (Buteo hemilasius): evidence from stable carbon isotope ratios. Acta Zoologica Sinica, 2003, 49(6): 764-768. |
| [113] | Casey M M, Post D M. The problem of isotopic baseline: reconstructing the diet and trophic position of fossil animals. Earth-Science Reviews, 2011, 106(1): 131-148. |
| [114] | Brittain R A, Schimmelmann A, Parkhurst D F, Craft C B. Habitat use by coastal birds inferred from stable carbon and nitrogen isotopes. Estuaries and Coasts, 2012, 35(2): 633-645. |
| [115] | 杨勇, 温俊宝, 胡德夫. 鸟类栖息地研究进展. 林业科学, 2011, 47(11): 172-180. |
| [116] | Conover M R. Recent changes in Ring-billed and California gull populations in the western United States. The Wilson Bulletin, 1983, 95(3): 362-383. |
| [117] | Thompson D R, Furness R W, Lewis S A. Diets and long-term changes in δ13C and δ15N value in northern fulmars (Fulmarus glacialis) form two northest Altlantic colonies. Marsh Ecololgy, 1995, 125: 3-11. |
| [118] | Bearhop S, Thompson D R, Waldron S, Russell I, Alexander G, Furness R W. Stable isotopes indicate the extent of freshwater feeding by cormorants Phalacrocorax carbo shot at inland fisheries in England. Journal of Applied Ecology, 1999, 36(1): 75-84. |
| [119] | Hobson K A, Montevecchi W A. Stable isotopic determinations of trophic relationships of great auks. Oecologia, 1991, 87(4): 528-531. |
| [120] | Bond J C, Esler D. Nutrient acquisition by female Harlequin Ducks prior to spring migration and reproduction: evidence for body mass optimization. Canadian Journal of Zoology, 2006, 84(9): 1223-1229. |
| [121] | Hobson K A, Sharp C M, Jefferies RL, Rockwell RF, Abraham KF. Nutrient allocation strategies to eggs by Lesser Snow Geese (Chen caerulescens) at a sub-Arctic colony. The Auk, 2011, 128(1): 156-165. |
| [122] | Langin K, Norris D, Kyser T, Marra P P, Ratcliffe L. Capital versus income breeding in a migratory passerine bird: evidence from stable-carbon isotopes. Canadian journal of Zoology, 2006, 84(7): 947-953. |
| [123] | Gauthier G, Bêty J, Hobson K A. Are greater snow geese capital breeders? New evidence from a stable-isotope model. Ecology, 2003, 84(12): 3250-3264. |
| [124] | Sénéchal É, Bêty J, Gilchrist H G, Hobson K A, Jamieson S E. Do purely capital layers exist among flying birds? Evidence of exogenous contribution to arctic-nesting common eider eggs. Oecologia, 2011, 165(3): 593-604. |
| [125] | Hobson K A, Thompson J E, Evans M R, Boyd S. Tracing nutrient allocation to reproduction in Barrow's Goldeneye. Journal of Wildlife Management, 2005, 69(3): 1221-1228. |
| [126] | Shealer D A. Differences in diet and chick provisioning between adult Roseate and Sandwich Terns in Puerto Rico. Condor, 1998, 100: 131-140. |
| [127] | Niebuhr V. Feeding strategies and incubation behaviour of wild herring gulls: an experiment using operant feeding boxes. Animal Behaviour, 1983, 31(3): 708-717. |
| [128] | Mizutani H, Kabaya Y, Wada E. Nitrogen and carbon isotope compositions relate linearly in cormorant tissues and its diet. Isotopenpraxis Isotopes in Environmental and Health Studies, 1991, 27(4): 166-168. |
| [129] | Gannes L Z, O'Brien D M, del Rio C M. Stable isotopes in animal ecology: assumptions, caveats, and a call for more laboratory experiments. Ecology, 1997, 78(4): 1271-1276. |
| [130] | Robbins C T, Felicetti L A, Sponheimer M. The effect of dietary protein quality on nitrogen isotope discrimination in mammals and birds. Oecologia, 2005, 144(4): 534-540. |
| [131] | Sturkie P D. Avian Physiology. 2nd ed. Ithaca NY: Cornell University Press, 1965: 1-766. |
| [132] | Schwarcz H P. Some theoretical aspects of isotope paleodiet studies. Journal of Archaeological Science, 1991, 18(3): 261-275. |
| [133] | Gannes L Z, Del Rio C M, Koch P. Natural abundance variations in stable isotopes and their potential uses in animal physiological ecology. Comparative Biochemistry and Physiology Part A: Molecular & Integrative Physiology, 1998, 119(3): 725-737. |
| [134] | Wassenaar L I, Hobson K A. A stable-isotope approach to delineate geographical catchment areas of avian migration monitoring stations in North America. Environmental Science & Technology, 2001, 35(9): 1845-1850. |
| [135] | Kelly J F, Finch D M. Tracking migrant songbirds with stable isotopes. Trends in Ecology & Evolution, 1998, 13(2): 48-49. |
| [136] | Chabot A A, Hobson K A, Van Wilgenburg S L, McQuat G J, Lougheed S C. Advances in linking wintering migrant birds to their breeding-ground origins using combined analyses of genetic and stable isotope markers. PLoS One, 2012, 7(8): e43627. |
| [137] | Reichlin T S, Hobson K A, Van Wilgenburg S L, Schaub M, Wassenaar L I, Martín-Vivaldi M, Arlettaz R, Jenni L. Conservation through connectivity: can isotopic gradients in Africa reveal winter quarters of a migratory bird? Oecologia, 2013, 171(2): 591-600. |