 2015, Vol. 35
2015, Vol. 35文章信息
- 党宏忠, 杨文斌, 李卫, 刘世增, 张友焱
- DANG Hongzhong, YANG Wenbin, LI Wei, LIU Shizeng, ZHANG Youyan
- 新疆杨树干液流的径向变化及时滞特征
- Radial pattern and time lag of sap flow in Populus alba var. pyramidalis
- 生态学报, 2015, 35(15): 5110-5120
- Acta Ecologica Sinica, 2015, 35(15): 5110-5120
- http://dx.doi.org/10.5846/stxb201401060046
-
文章历史
- 收稿日期:2014-01-06
- 网络出版日期:2014-11-03






2. 甘肃省治沙研究所, 兰州 730070
2. Gansu Desert Control Research Institute, Lanzhou 730070, China
树木从庞大的根系吸收的水分都要经边材的传输通道才能到达冠层,满足树木生理活动需求[1],因此,在分析树木的水分循环过程时,对树干液流的直接测定比对土壤水分含量的监测可能更可靠、更简便[2]。然而,调查发现大多数树种树干边材内部不同年轮处的液流传输能力并不相同,一般从外到内表现出“低-高-低”的变化格局[3],如果忽略液流速率在径向的这种差异,仅仅用单点测定的液流速率值与边材面积的乘积来估算单株耗水量,其误差可达-90%—300%[4],因此,确定树木边材输水能力的径向变化特征十分必要。
树干液流受树种、土壤与大气环境因子的共同影响,其中气象因子是驱动液流速率日变化最直接的因素,这与蒸腾作用受大气环境因子紧密驱动相一致。然而,树干液流与气象因子的日变化过程并不完全一致,其中峰值出现时刻上的差异(即时滞)反映了树木调控冠、根间水分传导过程能力的大小[5]。由于树木在午间前后的蒸腾变化主要受周围空气温、湿度驱动下的气孔的开放度所控制,因此液流与主要气象要素间的时滞被认为是评价树木冠层叶片蒸腾与大气耦合效应的良好指标,同时也反映了树体储存水分对蒸腾贡献的相对大小[6, 7, 8]。树木的时滞可达10 —130 min[9, 10],一般树体越高大,时滞时间越长[11],但是时滞的长短仍随树种、生境的差异而存在较大的变动[12]。
新疆杨(Populus alba L.var. pyramidalis)是我国分布最广泛的农田防护林树种之一,在北方具有广泛的适应性,但一直以来也存在着对其可能过度耗水的质疑。要提高对新疆杨的蒸腾耗水量估算的准确性,就必须明确其液流的径向变异并据此提出测定方法的最优选择与安排。对新疆杨液流时滞特征的系统研究也有助于揭示新疆杨蒸腾耗水过程对环境驱动的响应及其能力。为此,本研究将通过对生长在干旱绿洲环境中的新疆杨大树的树干液流开展连续两个生长季的监测,分析液流变化格局及其径向变化特征,揭示液流与主要气象要素间的时滞及其季节变化规律,为建立更完整的液流径向分布函数、寻求更可靠、更简便的单株液流估算方法提供依据,研究结果对于更深入地认识新疆杨个体适应干旱环境的水分利用策略具有重要意义。
1 研究区域与方法 1.1 研究区域概况试验地位于地处腾格里沙漠西缘的民勤沙生植物园附近(103°05′E,38°38′N,海拔1,367 m),属于典型的温带大陆性干旱荒漠气候,日温差大,多年年平均气温7.6℃,日平均气温最高、最低分别可达40.0 ℃和-30.8 ℃;年日照时间为2799.4h,≥10℃的活动积温为3036.4 ℃;年平均降雨量仅113.2 mm,而年平均蒸发量高达2604.3 mm。由于长期超采,地下水位已低于15 m,水资源短缺问题严峻。该地区为典型的风沙区,年均风沙日可达83d,主导风向为西北风,年平均风速2.5 m/s,最大风速可达23.0 m/s[13]。农田防护林以面积为2.5—6.0 hm2的窄林带小网格格局设置,树种以新疆杨(P. alba)和二白杨(Populus. gansuensis C. Wang and H.L. Yang)为主,树种较单一,大部分已达防护成熟期[14]。
1.2 样株的选择及液流测定装置布设于2011年4月开始在长势良好的防护林带中选择生长健康、冠形完整的32年生新疆杨单株3株,测量胸高、树高、枝下高、冠幅等形态学指标。采用严格按照Granier[15]原理制作的热扩散探针,探针长度分别为1、2、3、5 cm和8 cm,上下两探针间距离为10 cm,探针直径1.2 mm(1、2 cm和3 cm)和1.65 mm(5 cm和8 cm),安装、隔热、防水等步骤见文献[16]。在每株样株胸高位置的向北一侧安装1cm探针,随后以水平、垂直方向各10 cm的间距按逆时针螺旋上升的方向分别安装2、3、5 cm和8 cm探针,以避免各组针之间的干扰。五组探针布设在树干圆周的四分之一范围内,方位引起的差异可以忽略不计。采用220 V交流转接直流12 V铅酸蓄电瓶连续供电,加热探针供电量按照探针长度0.2 W/cm的标准执行,整个观测期内记录的上、下探针间温差不大于10 ℃。采用32通道SQ2040数采器(英国Grant公司)按30 s采集10 min记录的步长连续记录数据,采用Granier公式(式(1))计算液流速率(Js,m3 m-2 s-1),其中确定ΔTmax的时间步长为10 d[4, 17]。观测结束后采用生长锥法结合千分尺测量边材、心材等厚度并计算边材面积(表 1)。收集连续两年6—10月份的数据进行分析(删除因停电或传感器损坏期间的数据)。

式中,ΔT为两探针间的温差(℃),ΔTmax为木质部液流为零时的两探针间的温差(℃)。
| 样株号 No. of sample trees | 胸径 Diameter at breast height/cm | 树高 Height/m | 宽度Width/cm | 边材面积/cm2 Sapwood area | |||
| 髓心 Pith | 树皮和形成层 Bark& phloem | 心材 Heartwood | 边材 Sapwood | ||||
| 1号样株 Tree 1 | 30.88 | 15.24 | 0.50 | 0.33 | 7.29 | 7.33 | 527.57 |
| 2号样株 Tree 2 | 31.20 | 15.32 | 0.50 | 0.33 | 7.36 | 7.41 | 538.45 |
| 3号样株 Tree 3 | 30.04 | 15.16 | 0.50 | 0.31 | 7.10 | 7.11 | 498.33 |
简易气象站AR5架在距离样株约50 m外的空地(包括太阳辐射传感器,温、湿度传感器,风速、风向传感器和雨量传感器,以上传感器均来自美国Avalon公司,数采器采用32通道SQ2020,雨量筒上平面距地面高80 cm,其它传感器均安装在离地2 m高处)。气象站的信息采集与记录步长与测定液流时的设置相同。所记录、计算的气象要素包括太阳辐射(Ra)、大气温度(Ta)、大气相对湿度(Rh)、大气水汽压亏缺(VPD)和风速(Ws)等五项。
根据FAO Penman-Monteith方法,在以天为步长计算ET0时,Cn = 900,Cd = 0.34;在以小时步长的计算中,Cn = 37,Cd的取值在白天(6:00—20:00)为0.24,晚间(21:00—5:00)为0.96;土壤热通量G的计算按照以小时为时间尺度计算时的精度要求,以固定系数乘以净辐射计算而得,该系数在白天、晚间分别取0.5和0.1[18]。
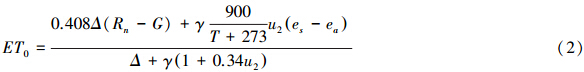
式中,ET0 为大气蒸发潜力 (mm),Δ为饱和水汽压随Ta变化的斜率(kPa/℃),Rn 为净辐射(MJ/m2),G为土壤热通量 (MJ/m2),γ为 空气温湿度常数 (kPa/℃);es为饱和水汽压 (kPa),ea为实际的水汽压 (kPa),u2 为2m高处的平均风速 (m/s)。大气水汽压亏缺VPD定义为es与ea的差。参数的取值及计算过程参照FAO 56执行[18]。
1.4 数据分析方法采用origin 9.0 提供的Peak形式函数中的Guass方程拟合液流速率、大气因子的日变化过程并判识峰值时刻。

根据Guass方程中各变量-参数的数学意义,式中变量Js为液流速率,x为时刻(北京时间,取值0—24),xc为Js达到峰值时的时刻,A,w为方程参数。为便于分析,本文把Js峰值时刻与ET0、 VPD峰值时刻的时间差(时滞)分别记作ΔJ-E和ΔJ-V,当差值为正值时,表示液流峰值滞后于所对应的气象因子的峰值时刻,负值表示液流的峰值时刻提前于气象因子的峰值时刻。
采用Statistica 10.0提供的逐步回归方程(后退式)进行液流速率与气象因子间的回归分析。
2 结果与分析 2.1 新疆杨树干边材不同深度处液流速率的变化测定表明,新疆杨边材沿径向各层处的液流速率日平均值整体上随着深度表现出“低-高-低”的格局,其中液流速率最高位点在边材中间3 cm(21.23—24.65 kg dm-2 d-1) 或5 cm处(23.62 kg dm-2 d-1),最低点在边材两端1 cm处(5.20—3.83 kg dm-2 d-1)或8 cm处(5.81 kg dm-2 d-1)。方差分析(结果略)表明,除了最外层与最里层处边材液流速率间差异不显著外(Tree 1和Tree 2),其它各层液流速率的平均值间均存在显著差异(P = 0.05),反映了新疆杨边材液流速率存在着显著的径向变异(图 1)。

|
| 图1 边材不同深度处液流速率的比较 Fig.1 Comparision of Js at different depths of xylem a,b,…,e 为方差分析Duncan检验多重比较结果标记(P = 0.05) |
以2 cm处(长度为2 cm探针是Granier最初设计的标准长度)液流速率(Js-2)为参考值,建立的其与1、3、5 cm 和8 cm处液流速率(Js-1、Js-3、Js-5和Js-8)间的简单回归分析(图 2),结果表明各层处的液流速率与Js-2间均具有显著的线性相关(R2= 0.81—0.99),表明五个深度处的液流传输具有相近的变化格局与驱动过程。在5个位点中,Js-2与Js-3、Js-5间的回归系数均高于与Js-1、Js-8间的回归系数,5、3 cm位点所在的边材是液流输送的主导部分,其中对于Tree1和Tree 3,Js-5和Js-2间的回归系数为1.35—1.64,高于Js-3和Js-2间的值(1.18—1.24),边材中输水能力最强的位点更接近于5 cm处;而对于Tree 2,Js-3和Js-2的回归系数(1.61)高于Js-5和Js-2间的(0.81),指示着该样株边材液流传输最高的点更接近于3 cm处,5 cm处的液流速率已经明显下降,可见树木液流速率在径向不仅存在着变化,而且这种变化也存在着个体间的不同。对于大树,传统上2 cm长的探针不足以探测到最大的液流速率位点。

|
| 图2 不同深度处液流速率与Js-2间的线性关系(Js-2为边材深度2 cm的液流速率,依次类推) Fig.2 The linear relationships of sap flow velocity at different depths with Js-2(Js-2 was sap flow velocity at depth of 2 cm,and so on) |
树木蒸腾耗水过程受不同气象要素共同驱动的效应非常复杂,为了简化并筛选出主导的气象要素,以2011年2 cm处液流速率(Js-2)为因变量,以同期观测的5项气象要素(Ra、Ta、Rh、VPD和Ws)为自变量进行逐步回归分析,结果显示(表 2),对于3株样树,VPD和Ra均为5个气象要素中首先被引入的变量且通过了差异显著性检验,其校正后的回归系数(b*)明显高于其它变量,表明它们是5项气象要素中驱动该地区树木蒸腾过程的首要、主导因子,二者分别表达了大气水汽状况与热量状况,共同驱动着该地区树木液流的过程。
| 样株 Sample trees | 气象因子 Meteorologic factors | 校正回归系数b* Adjusted regression coefficient | b*的标准误差 Std.Err. of b* | 显著性检验值 t(n-5-1) | P | F |
| VPD: 大气水汽压亏缺the vapor pressure deficit of atmosphere; Ra: 太阳辐射solar radiation; Ws: 风速wind speed; Ta: 大气温度air temperature; Rh : 大气相对湿度relative humidity of atmosphere; *为在逐步回归分析中保留的变量,所有入选的因子均通过显著性检验(P = 0.05) | ||||||
| Tree 1 | VPD* | 0.47 | 0.03 | 15.05 | 0.00 | F (5,1637)=1533.5 |
| Ra* | 0.42 | 0.02 | 26.19 | 0.00 | ||
| Ws* | 0.07 | 0.02 | 4.52 | 0.00 | ||
| Ta | 0.04 | 0.03 | 1.75 | 0.08 | ||
| Rh | 0.00 | 0.02 | 0.21 | 0.84 | ||
| Tree 2 | VPD* | 0.58 | 0.03 | 22.47 | 0.00 | F (5,1587)=2575 |
| Ra* | 0.48 | 0.01 | 39.01 | 0.00 | ||
| Ws* | 0.05 | 0.01 | 4.06 | 0.00 | ||
| Rh | 0.03 | 0.02 | 2.14 | 0.03 | ||
| Ta | -0.04 | 0.02 | -1.75 | 0.08 | ||
| Tree 3 | VPD* | 0.55 | 0.02 | 22.38 | 0.00 | F (5,1587)=2841.7 |
| Ra* | 0.53 | 0.01 | 45.29 | 0.00 | ||
| Ws* | 0.05 | 0.01 | 4.62 | 0.00 | ||
| Ta* | -0.07 | 0.02 | -3.29 | 0.00 | ||
| Rh | 0.03 | 0.02 | 1.95 | 0.05 | ||
上述分析已证实VPD与Ra是驱动干旱区新疆杨液流速率日变化过程的主要气象因子,但它们与液流日变化过程的一致性需要更进一步的分析。以生理活动最为旺盛的7月份为时间片断[19, 20],挑选典型晴天日(2012)并选择主导气象因子Ra、VPD及综合变量ET0,与Js,5一起采用Guass方程拟合其日变化过程。图 3展示了三者与Js之间峰值到达时刻的相对差异关系。可以看出,Js的峰值时刻要比Ra、ET0明显滞后,但比VPD的峰值时刻明显提前。Ra与ET0峰值出现的时刻完全一致(这也是可采用ET0代替Ra来描述液流时滞特征的依据),但明显早于VPD的峰值时刻。在树木生长旺盛的7月份,树干液流在上午随着ET0的上升而增加,在ET0达到峰值时液流速率(Js-5)占峰值的94.16%,仍然在上升,经过一段时间(ΔJ-E)才到达峰值。该季节新疆杨旺盛的生理活动,使得即便午间前后较高的ET0也并不能限制树木液流传输速率的继续升高,以至于在液流达到峰值时的ET0已下降到只占峰值的88.35%,但这时的VPD(-3.23 kPa)的绝对值尚没达到最高值(占峰值的97.61%),等到VPD绝对值达到峰值时,Js-5下降到约占峰值的97.28%。可以看出,在上午,当VPD处于较低水平而Ra逐渐上升时,液流与VPD呈正相关的关系,当VPD上升到某一阈值后(在7月份为-3.23 kPa),液流与VPD呈负相关,经过一段时间(ΔJ-V),二者又呈共同下降的正相关关系。

|
| 图3 典型晴天日液流速率与气象变量间的时滞关系(以2012年7月份数据为例绘图,Js为5 cm处液流速率) Fig.3 Relationships of time lag between Js and meteorological varibles during typical sunny days (graphic is with data in July,2012 and Js is Js-5) |
对于其它位点的峰值时刻,我们一并列于表 3来更进一步比较大气对不同层次边材液流日变化过程驱动的差异。表 3中参数xc代表了在典型晴天日各变量峰值出现的时刻,可以看出,新疆杨大树树干边材除了1 cm 位点外,其它四个位点(2 cm,3 cm,5 cm和8 cm)处液流速率峰值时刻基本一致,均约在14:00之后,且明显晚于Ra与ET0的峰值时刻(72—99 min),早于VPD的峰值时刻(40—59 min)。1 cm 位点处的液流速率较小,且日变化过程较平缓,其峰值较其它层次提前4—14 min(2011)和111—123 min(2012)。Ra与ET0峰值出现的时刻相同,均比VPD峰值出现的时刻早132 min(2011)和139 min(2012)。
| 变量与参数 Varibles and parameters | 2011年7月 June,2012 | 2012年7月 July,2012 | ||||||||
| A | xc | w | H | R2 | A | xc | w | H | R2 | |
| A,xc,w为Guass方程(公式(3))中的参数,H为曲线的峰值,R2 为决定系数 | ||||||||||
| Ra | 7648.45 | 12.84 | 6.16 | 990.86 | 0.97 | 7363.96 | 12.61 | 6.01 | 978.38 | 0.98 |
| VPD | 23.70 | 15.04 | 6.49 | 2.91 | 0.78 | 21.48 | 14.93 | 6.56 | 2.61 | 0.83 |
| ET0 | 4.93 | 12.84 | 5.32 | 0.74 | 0.94 | 4.97 | 12.61 | 5.42 | 0.73 | 0.96 |
| Js-1 | 219.41 | 13.97 | 11.66 | 15.01 | 0.11 | 58.78 | 12.20 | 16.20 | 2.90 | 0.14 |
| Js-2 | 231.53 | 14.11 | 7.90 | 23.37 | 0.93 | 194.15 | 14.11 | 8.00 | 19.37 | 0.83 |
| Js-3 | 304.08 | 14.04 | 8.37 | 28.98 | 0.89 | 245.76 | 14.26 | 7.63 | 25.71 | 0.78 |
| Js-5 | 283.10 | 14.04 | 8.14 | 27.74 | 0.84 | 239.83 | 14.06 | 8.03 | 23.84 | 0.83 |
| Js-8 | 107.68 | 14.21 | 8.76 | 9.81 | 0.73 | 124.40 | 14.19 | 9.04 | 10.98 | 0.57 |
新疆杨液流速率与ET0及VPD在7月份的时滞,是否在不同季节表现出不同?统计分析表明,在主要的生长季(6—10月份),研究地区ET0的季节变化呈现递减的趋势,其中各月ET0日平均值变化区间为5.96—2.80 mm/d(2011)、5.49—2.53 mm/d(2012)。新疆杨树干边材五个深度处的液流速率与ET0间峰值的时滞长度总体上均呈逐渐增加的趋势(图 4)。6月份,随着展叶的完成,新疆杨生理活动逐渐旺盛,此时大气的驱动力最强,时滞ΔJ-E 约为(70±11)min(除Js-1外),此时冠层蒸腾与ET0间仍有较强的耦合效应。之后的7—9月间,随着冠层生长发育的进一步完善与成熟,树木继续维持较高的水分耗散量,气孔对大气蒸发力的调控能力逐渐加强,表现在液流的峰值有所降低而上升期延长,时滞ΔJ-E逐渐增加。到了10月份,大部分叶片生理活动明显下降,树木液流受大气热量驱动的效应明显减弱,各位点(除Js-1外)时滞ΔJ-E平均约为(110±11)min(2011)和(128±20)min(2012)。

|
| 图4 液流速率与ET0间时滞(ΔJ-E)、与VPD间时滞(ΔJ-V)的季节变化 Fig.4 Seasonal variation pattern of time lag between Js and ET0(ΔJ-E),Js and VPD(ΔJ-V) |
与ET0不同,液流速率峰值往往提前于VPD峰值而来到,这种提前量在6月份最大,约为(73±11) min(2011)和(63±15) min(2012)(除Js-1外),在大气水汽压亏缺还没到达非常严峻的局面之前,液流传输过程已经经历了峰值并逐渐下降,避免与大气水汽压亏缺出现峰值叠加效应;之后,随着树木生理能力的逐渐健全和冠层需水量的增加,液流峰值后移,VPD与Js峰值间的时滞缩短,到10月份仅为(20±11) min(2011)和(-8±20) min(2012)(负值表示液流滞后),此时树木较弱的蒸腾生理活动基本受控于大气水汽压亏缺的驱动而表现出与其近一致的变化过程。统计表明,6—10月间,研究地区VPD日平均值的逐月变化呈递减的趋势,变化区间为2.14—0.68 kpa(2011)、1.81—0.65 kPa(2012)。
新疆杨边材1 cm位点处的时滞ΔJ-E与ΔJ-V虽然也表现出与其它位点相近似的季节变化规律,但在2012年的6—8月,Js-1却提前于ET0约16 —32 min,更显著地提前于VPD(157—165 min),新疆杨最外层边材的输水性明显受大气蒸发的控制而下降,这可能表明新疆杨最外层边材与表层根系的水分传导能力直接对应,在夏日强大的大气-根系-土壤水势梯度下,最外层的边材首先响应但更早地降低液流传输。
3 结论新疆杨树干边材液流具有明显的径向差异,各层边材液流具有相一致的变化过程,与Js-2间均具有较强的线性相关关系(R2= 0.81—0.99)。液流速率最高点位于形成层下3—5 cm处,Js-5、Js-3最高分别可达Js-2的1.64倍和1.61倍,常规的2 cm长的TDP探针难以到达大树液流速率最高的位点。在5项常规气象要素中,VPD和Ra是该地区驱动新疆杨液流进程的主导气象因子。综合变量ET0集合了多种气象因子的信息且具有与Ra一致的日变化进程(峰值时刻),在分析大气对液流的驱动过程中更具有优势。
新疆杨边材各位点处不仅液流速率明显不同,而且均具有明显的时滞特征,液流速率的峰值一般滞后于ET0而提前于VPD,从6到10月份,时滞ΔJ-E表现为递增趋势,随着冠层叶片生理特征的成熟及大气蒸发需求的下降,液流传输受太阳辐射的约束逐渐减弱;与之相反,时滞ΔJ-V在6月份最大,之后表现出递减的趋势,Js与VPD的日变化过程渐趋一致。时滞的这种变化表明在新疆杨生长季的早期,液流传输主要受大气热量因子(Ra)的驱动,树木贮存水分对液流日变化的缓冲效应较小;而在后期,这种缓冲效应趋于加强,液流过程主要受大气水汽因子(VPD)的驱动。
4 讨论通过对新疆杨边材3个主要深度处液流速率的观测分析,揭示了新疆杨边材液流速率存在着的径向差异,为今后通过建立完整的径向分布函数来提高单株蒸腾估算的精度提供了基础。液流速率沿径向的分布可能随树种而异,但大多数可归于递减型[11]和单峰型[21, 22]两类,对于新疆杨其径向分布属于典型的单峰型并符合Guassian函数。有研究发现对于胸径达75.6 cm的大树用3.3 cm的探针很难触及液流的主要位点[23],同样发现传统的标准长度的探针(2 cm)也不能到达新疆杨大树液流的最高位点(3—5 cm处),但是各层液流速率与J2间存在较紧密的线性相关性(R2 = 0.81—0.99),不同深处的液流之间具有较一致的变化过程,为根据单点测定来估算径向格局、并进而估算单株耗水量提供了简化途径。
为了尽可能地探测到深处边材的液流传输过程,采用了长度为8 cm的探针并监测到了新疆杨树干深处较小的液流速率,但在观测结束后用生长锥钻取髓芯并结合颜色判断测量的边材厚度为7.11—7.41 cm,这一方面说明对于散孔材新疆杨来说,绝对的边材与心材的“分界线”可能并不存在,液流速率随着深度而下降的过程是渐进式的,另一方面,传统的通过颜色区分心材、边材可能并不十分准确,如果需要估算单株的液流通量,最好是基于液流完整的径向分布函数通过积分求算。
驱动新疆杨液流变化的主要大气因素是VPD和Ra,它们分别代表着树木冠层蒸腾的两类驱动力(大气水汽和大气热量)。太阳辐射对气孔导度具有直接或间接的正效应,树木的蒸腾会随辐射的上升而增高。对我国从南到北的研究结果综合比较来看,比较一致地认为太阳辐射是影响液流变化的首要气象因子[20, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34],树木的整个日蒸腾过程对辐射的响应程度最突出,其原因除了可见光对气孔开闭的直接影响外,可能也与太阳辐射对叶面和空气的增温效应能迅速提高叶片内外的水汽扩散梯度,促进蒸腾的进行有关。
然而,不少研究也报道水汽压亏缺对液流变化的重要性[26, 35, 36, 37, 38, 39],VPD表达了空气实际的水汽压与相同温度下饱和水汽压的差值,因而也受大气温、湿度的共同影响。对于大树,特别是有灌溉条件下的绿洲防护林,树木叶片所含水分一般接近饱和,在叶片周围空气湿度距离饱和水汽压的差值越大,水分从叶片向大气中扩散的速率越高。国外无论是对冷杉林(Picea abies)还是对热带雨林的研究均有树干液流与水汽压亏缺的相关性要高于与太阳辐射的报道[40, 41]。在我国南方,不论是对于阔叶林还是针叶林,树干液流与水汽压亏缺均呈极显著相关,其相关性高于太阳辐射[42, 43]。由于地处干旱荒漠区,太阳辐射普遍强烈,而大气水汽状况差,这可能是VPD对液流影响更灵敏的主要原因。
太阳辐射和水蒸汽压亏缺对液流的影响并不总是正效应,在到达一定值后会呈负效应。这表明两因子并不是独立存在的,而是对植物的水分利用产生协同效应,单因子分析难以准确地反映他们的真实关系。实际上太阳辐射不仅通过对大气温度的影响对树干液流产生直接影响[44],太阳辐射和温度也都影响水汽压亏缺,而叶面气孔蒸腾的驱动力直接和水汽压亏缺相关。因此,在分析驱动液流的环境效应时,综合变量ET0作为表达有效能量的因子[45],要比单一的Ra或VPD更具有可靠性。
虽然太阳辐射和大气水汽压亏缺与液流间有紧密的相关关系,但二者与液流的变化过程并不完全一致,而是存在着时滞。我国有学者通过逐次错位移动两列数据的方法得出107欧美杨(P. × euramericanacv)液流峰值时刻滞后Ra峰值30—60 min,提前VPD峰值120—150 min[46],对于马占相思(Acacia mangium)液流比光合有效辐射滞后40—110 min[47, 48]。 新疆杨在主要的生长季中,时滞大小存在明显的季节差异,Js相对Ra峰值(与ET0相同)的时滞为70—128 min,Js相对于VPD提前8—73 min,Js相对Ra的时滞较VPD明显大,这表明干旱荒漠区树木液流与VPD间的耦合较与Ra的紧密,这与干旱荒漠特征是相一致的。
研究表明,树木时滞的长短与树木的水容有着紧密的联系[49],树木由于存在水容而具备水分储备和释放能力,从而缓解蒸腾过程对根系吸水的压力。同时,树木体内释放的水分在一定程度上可以缓解叶温的升高和气孔的关闭,进而使叶片保持较高的光合速率,因此可以认为水容大的树种一般其与VPD的时滞更加明显,体现了对大气蒸腾驱动的更强的缓冲能力。研究发现新疆杨液流与VPD的时滞在整个生长季中呈逐渐下降的趋势,这与该过程中树体水容能力逐渐下降是一致的。
在环境因子中,土壤水分含量的过高过低都不利于植物对水分的吸收与利用。在较小的时间尺度(如一天或小时),液流对土壤水分含量的变化并不“敏感”,这与树木强大的根系分布有直接关系。由于不同土壤层次间水分动力学特征的差异,及树木根系分布的空间变异性,很难从本质上建立起土壤水分含量与树干液流间的固定关系模式。由于定期灌溉,防护林带内土壤含水量较充足且各个月份间变化不大,土壤含水量对树干液流格局的影响很小可忽略不计,这也在干旱区梭梭(Haloxylon ammodendron)的液流监测中得到证实[50],说明即便在干旱区,土壤水分状况对树木在较长时间内液流总量的影响要比在较小时间尺度上液流过程的影响更明显。
| [1] | Tyree M T,Zimmermann M H.Xylem Structure and the Ascent of Sap.2nd edn.Heidelberg:Springer Verlag,2002:49-88. |
| [2] | Wilson K B,Hanson P J,Mulholland P J,Baldocchi D D,Wullschleger S D.A comparison of methods for determining forest evapotranspiration and its components:sap-flow,soil water budget,eddy covariance and catchment water balance.Agricultural and Forest Meteorology,2001,106(2):153-168. |
| [3] | Phillips N,Oren R,Zimmermann R.Radial patterns of xylem sap flow in non-,diffuse-and ring-porous tree species.Plant Cell and Environment,1996,19(8):983-990. |
| [4] | Lu P,Urban L,Zhao P.Granier's thermal dissipation probe (TDP) method for measuring sap flow in trees:theory and practice.Acta Botanica Sinica,2004,46(6):631-646. |
| [5] | Ford C R,Goranson C E,Mitchell R J,Will R E,Teskey R O.Diurnal and seasonal variability in the radial distribution of sap flow:predicting total stem flow in Pinus taeda trees.Tree Physiology,2004,24(9):951-960. |
| [6] | Phillips N G,Ryan M G,Bond B J,McDowell N G,Hinckley T M,Čermák J.Reliance on stored water increases with tree size in three species in the Pacific Northwest.Tree Physiology,2003,23(4):237-245. |
| [7] | Dawson T E,Burgess S S O,Tu K P,Oliveira R S,Santiago L S,Fisher J B,Simonin K A,Ambrose A R.Nighttime transpiration in woody plants from contrasting ecosystems.Tree Physiology,2007,27(4):561-575. |
| [8] | Moore G W,Cleverly J R,Owens M K.Nocturnal transpiration in riparian Tamarix thickets authenticated by sap flux,eddy covariance and leaf gas exchange measurements.Tree Physiology,2008,28(4):521-528. |
| [9] | Čermák J,Kučera J,Bauerle W L,Phillips N,Hinckley T M.Tree water storage and its diurnal dynamics related to sap flow and changes in stem volume in old-growth Douglas-fir trees.Tree Physiology,2007,27(2):181-198. |
| [10] | Scholz F C,Bucci S J,Goldstein G,Meinzer F C,Franco A C,Miralles-Wilhelm F.Temporal dynamics of stem expansion and contraction in savanna trees:withdrawal and recharge of stored water.Tree Physiology,2008,28(3):469-480. |
| [11] | Horna V,Schuldt B,Brix S,Leuschner C.Environment and tree size controlling stem sap flux in a perhumid tropical forest of Central Sulawesi,Indonesia.Annals of Forest Science,2011,68(5):1027-1038. |
| [12] | Cienciala E,Kučera J,Malmer A.Tree sap flow and stand transpiration of two Acacia mangium plantations in Sabah,Borneo.Journal of Hydrology,2000,236(1/2):109-120. |
| [13] | 常兆丰,邱国玉,赵明,杨自辉,韩富贵,仲生年,李爱德,刘淑娟.民勤荒漠区植物物候对气候变暖的响应.生态学报,2009,29(10):5195-5206. |
| [14] | 张锦春,汪杰,李爱德,俄有浩.民勤绿洲杨树林带生长特性及更新年龄初探.甘肃林业科技,2000,25(4):1-6. |
| [15] | Granier A.A new method of sap flow measurement in tree stems.Annales des Sciences Forestières,1985,42(2):193-200. |
| [16] | 党宏忠,张劲松,赵雨森.应用热扩散技术对柠条锦鸡儿主根液流速率的研究.林业科学,2010,46(3):29-36. |
| [17] | Granier A.Evaluation of transpiration in a Douglas-fir stand by means of sap flow measurements.Tree Physiology,1987,3(4):309-320. |
| [18] | Allen R G,Pereira L S,Raes D,Smith M.Crop evapotranspiration-Guidelines for computing crop water requirements//FAO Irrigation and Drainage Paper 56.Rome:FAO,1998:5-15. |
| [19] | 马长明,管伟,叶兵,袁玉欣,王金凤,李淑梅.利用热扩散式边材液流探针(TDP)对山杨树干液流的研究.河北农业大学学报,2005,28(1):39-43. |
| [20] | 张劲松,孟平,孙惠民,周泽福,于延凤.毛乌素沙地樟子松蒸腾变化规律及其与微气象因子的关系.林业科学研究,2006,19(1):45-50. |
| [21] | Lu P,Muller W J,Chacko E K.Spatial variations in xylem sap flux density in the trunk of orchard-grown,mature mango trees under changing soil water conditions.Tree Physiology,2000,20(10):683-692. |
| [22] | Ford C R,McGuire M A,Mitchell R J,Teskey R O.Assessing variation in the radial profile of sap flux density in Pinus species and its effect on daily water use.Tree Physiology,2004,24(3):241-249. |
| [23] | 蒋文伟,杨广远,赵明水,杨淑贞.天目山柳杉树干液流的昼夜及季节变化.南京林业大学学报:自然科学版,2012,36(5):77-80. |
| [24] | 丁日升,康绍忠,龚道枝.苹果树液流变化规律研究.灌溉排水学报,2004,23(2):21-25. |
| [25] | 于占辉,陈云明,杜盛.黄土高原半干旱区人工林刺槐展叶期树干液流动态分析.林业科学,2009,45(4):53-59. |
| [26] | 于占辉,陈云明,杜盛.黄土高原半干旱区侧柏(Platycladus orientalis)树干液流动态.生态学报,2009,29(7):3970-3976. |
| [27] | 于红博,杨劼,臧春鑫,徐延达.皇甫川流域中国沙棘树干液流日变化及其相关因子.生态学杂志,2008,27(7):1071-1076. |
| [28] | 孙慧珍,孙龙,王传宽,周晓峰.东北东部山区主要树种树干液流研究.林业科学,2005,41(3):36-42. |
| [29] | 段玉玺,秦景,贺康宁,张进虎,赵奎,孙毅.白榆、新疆杨液流动态比较研究.中国沙漠,2008,28(6):1136-1144. |
| [30] | 郭宝妮,张建军,王震,茹豪,黄明.晋西黄土区刺槐和油松树干液流比较.中国水土保持科学,2012,10(4):73-79. |
| [31] | 张友焱,周泽福,党宏忠,李卫.利用TDP茎流计研究沙地樟子松的树干液流.水土保持研究,2006,13(4):78-80. |
| [32] | 蒋文伟,汤富彬,刘志梅,赵明水.天目山柳杉古树的液流特征研究.林业科学研究,2012,25(1):58-64. |
| [33] | 赵奎,丁国栋,原鹏飞,张进虎,孙毅,刘楠.盐池毛乌素沙地白榆树干液流研究.水土保持研究,2008,15(6):85-88. |
| [34] | 桑玉强,刘全军,吴文良,张劲松,孟平,周择福.毛乌素沙地新疆杨生长季节蒸腾耗水规律.东北林业大学学报,2008,36(9):28-30,47-47. |
| [35] | 屈艳萍,康绍忠,夏桂敏,李王成.甘肃石羊河流域人工种植长穗柽柳树干液流量变化规律研究.西北植物学报,2005,25(11):2259-2265. |
| [36] | 熊伟,王彦辉,徐德应.宁南山区华北落叶松人工林蒸腾耗水规律及其对环境因子的响应.林业科学,2003,29(2):1-7. |
| [37] | 吴芳,陈云明,于占辉.黄土高原半干旱区刺槐生长盛期树干液流动态.植物生态学报,2010,34(4):469-476. |
| [38] | 牛丽,岳广阳,赵哈林,张铜会,赵学勇,刘新平,赵玮.利用液流法估算樟子松和小叶锦鸡儿人工林蒸腾耗水.北京林业大学学报,2008,30(6):1-8. |
| [39] | 孙龙,王传宽,杨国亭,张全智,周晓峰.应用热扩散技术对红松人工林树干液流通量的研究.林业科学,2007,43(11):8-14. |
| [40] | Granier A,Huc R,Barigah S T.Transpiration of natural rain forest and its dependence on climatic factors.Agricultural and Forest Meteorology,1996,78(1/2):19-29. |
| [41] | Granier A,Claustres J P.Variations of water relations within a spruce (Picea abies) growing in natural conditions.Acta Oecologica Oecologia Plantarum,1989,10(3):295-310. |
| [42] | 史梅娟,郑怀舟,王健,郑群瑞,朱锦懋.3种优势树种深秋树干液流特征与环境因子的关系.福建师范大学学报:自然科学版,2010,26(3):57-61. |
| [43] | 刘敏,贺康宁,于洋.青海云杉树干液流研究.水土保持应用技术,2009,(1):1-3,6-6. |
| [44] | 李海光,李钢铁,王彦辉,管伟,刘建立.六盘山北侧华北落叶松耗水特性研究.内蒙古农业大学学报:自然科学版,2008,29(3):72-76. |
| [45] | Sun G,Alstad K,Chen J Q,Chen S P,Ford C R,Lin G H,Liu C F,Lu N,McNulty S G,Miao H X,Noormets A,Vose J M,Wilske B,Zeppel M,Zhang Y,Zhang Z Q.A general predictive model for estimating monthly ecosystem evapotranspiration.Ecohydrology,2011,4(2):245-255. |
| [46] | 申李华,张志强,刘晨峰,朱金兆,查同刚,方显锐,陈军.沙地杨树人工林树干液流特征.中国水土保持科学,2007,5(1):88-92. |
| [47] | 赵平,饶兴权,马玲,蔡锡安,曾小平.马占相思(Acacia mangium)树干液流密度和整树蒸腾的个体差异.生态学报,2006,26(12):4050-4058. |
| [48] | 王华,赵平,蔡锡安,马玲,饶兴权,曾小平.马占相思树干液流与光合有效辐射和水汽压亏缺间的时滞效应.应用生态学报,2008,19(2):225-230. |
| [49] | Burgess S S O,Dawson T E.Using branch and basal trunk sap flow measurements to estimate whole-plant water capacitance:a caution.Plant and Soil,2008,305(1/2):5-13. |
| [50] | 许浩,张希明,闫海龙,孙红叶,单立山.塔克拉玛干沙漠腹地梭梭(Haloxylon ammodendron)蒸腾耗水规律.生态学报,2008,28(8):3713-3720. |