 2015, Vol. 35
2015, Vol. 35文章信息
- 熊莉, 徐振锋, 杨万勤, 殷睿, 唐仕姗, 王滨, 徐李亚, 常晨晖
- XIONG Li, XU Zhenfeng, YANG Wanqin, YIN Rui, TANG Shishan, WANG Bin, XU Liya, CHANG Chenhui
- 川西亚高山粗枝云杉人工林地上凋落物对土壤呼吸的贡献
- Aboveground litter contribution to soil respiration in a subalpine dragon spruce plantation of western Sichuan
- 生态学报, 2015, 35(14): 4678-4686
- Acta Ecologica Sinica, 2015, 35(14): 4678-4686
- http://dx.doi.org/10.5846/stxb201312042891
-
文章历史
- 收稿日期:2013-12-04
- 修订日期:2014-09-09






地表凋落物是森林生态系统的基本组成部分,在保持水土、涵养水源、保育生物多样性、幼苗更新以及物质循环中具有十分重要的地位[1]。据估算,全球土壤呼吸释放CO2通量约为68 Pg/a,其中有50 Pg/a来自凋落物和土壤有机质分解[2]。地表凋落物是森林土壤呼吸的重要来源,其随着森林类型、气候带和季节而变化[3, 4]。凋落物数量及其分解速率在很大程度上影响着土壤有机质的形成和对植物养分的供应,或通过影响土壤水热因子,土壤微环境以及微生物种类和数量,直接或间接地影响土壤CO2通量[4, 5, 6, 7]。可见,凋落物对了解森林土壤碳库循环以及土壤呼吸的源汇问题具有重要意义。凋落物去除试验是一种控制土壤碳输入来源及速率的野外试验,主要研究凋落物是如何影响土壤有机质和养分的积累和动态[8]。目前,国内关于凋落物对土壤呼吸的影响多集中在低海拔地区森林[9, 10, 11, 12, 13, 14],而对高寒森林的研究鲜见报道。
川西亚高山森林地处青藏高原东缘和长江上游,海拔2458—4619 m,是我国第二大林区的主体,在区域气候调节、水土保持、水源涵养和生物多样性保育方面具有不可替代的作用和地位[15],并作为长江上游重要的生态屏障。然而,近几十年来川西亚高山森林被过度采伐形成了大量采伐迹地,为恢复生态平衡的同时,积极恢复形成了大面积的人工针叶林,其中云杉林是构成该区亚高山针叶林的主体[16]。粗枝云杉(Picea asperata)是我国特有树种,四季常绿,在涵养水源,保持长江中下游水土流失等方面具有重要作用,是川西亚高山人工林典型的优势树种。有研究表明,其地表凋落物层约为1.2—10.8 cm,枯枝落叶储量约为76.9×103 kg/hm2,碳储量高[17]。川西亚高山粗枝云杉人工林地上凋落物可能是该林区土壤呼吸极为重要的来源,但至今仍没有相关研究量化粗枝云杉人工林地上凋落物对土壤呼吸的贡献。
鉴于此,本研究以川西亚高山粗枝云杉人工林为研究对象,动态监测两种处理(对照和去除凋落物)下粗枝云杉人工林土壤呼吸速率及其相关环境因子,估算了地上凋落物对土壤呼吸的贡献。本研究不仅有助于深入理解地表凋落物对土壤呼吸的贡献及其影响机制,而且可为粗枝云杉人工林土壤碳库管理提供理论依据。
1 材料与方法 1.1 研究区域和研究样地概况本研究区域位于四川省阿坝州理县毕棚沟自然保护区(102°53′—102°57′E,31°14′—31°19′N,海拔2458—4619 m)。地处青藏高原四川盆地的过渡地带,四姑娘山北麓,属丹巴-松潘半湿润气候,区域内年平均温度为2—4 ℃,最高气温23.7 ℃,最低温度-18.1 ℃。年均降水量850 mm,降雨主要分布在生长季节。研究区域土壤类型为有机层较厚的雏形土,主要林型有岷江冷杉(Abies faxoniana)原始林、岷江冷杉-红桦(Betula albo-sinensis)混交林和岷江冷杉次生林;林下灌木主要有箭竹(Fargesia spathacea)、高山杜鹃(Rhododendron delavayi)、三颗针(Berberis sargentiana)、红毛花楸(Sorbus rufopilosa)、沙棘(Hippophae rhamnoides)和扁刺蔷薇(Rosa sweginzowii)等;草本主要有蟹甲草(Cacalia forrestii)、高山冷蕨(Cystopteris montana)、苔草(Vittaria flexuosa)和莎草(Schizaea digitata)等。
本研究选取粗枝云杉人工林(102°56′E,31°18′N,海拔3035 m)为试验样地,乔木层的郁闭度一般为0.8,云杉为绝对的优势树种,占乔木个体数的70%以上。其树高在17 m左右,胸径在18—25 cm,林下植物以小檗(Berberis)为主,地被物以禾草(Agrostis)类、莎草(Cyperes)类植物为主,盖度在10%—80%之间不等[16]。样地土层浅薄,土壤为多为山地棕壤、山地暗棕壤和山地灰褐土。枯枝落叶层约为2—5 cm,腐殖质层平均厚度约为10 cm。0—20 cm土壤有机碳、全氮、全磷分别为159.1,7.8,0.9 g/kg,pH为5.8。
1.2 试验样地设计2012年11月中旬,选择粗枝云杉人工林内地表凋落物、树体长势、大小等相对均匀的地方设置3个20 m×20 m样地,每个样地按着对角线均匀设置5个正常允许凋落物进入的PVC环(直径20 cm,高6 cm,平行地表插入土壤约3 cm),为对照处理(CK);同时在旁边设置5个去除凋落物(NL)的5个PVC环。去除凋落物(NL)第一次处理时,移走样地内地表凋落物,安装PVC环后,在环上部设置50 cm×50 cm、网格为1 mm×1 mm的尼龙网,阻止凋落物进入。
1.3 土壤呼吸、土壤温度和水分测定土壤呼吸通量采用Li-8100土壤碳通量自动测量系统 (Li-Cor Inc,NE,USA)测定。土壤呼吸测定从2012年11月中旬开始至2013年11月中旬,大约每个月观测1次,每次测定在9:00—12:00完成。且在每次测定前一天,在不扰动土壤和PVC环的前提下,从根部齐地剪除地上绿色植物。测定土壤呼吸的同时,土壤5 cm深度的温度和水分(体积含水量,%)分别采用Li-8100自带温度探针和水分传感器观测。另外,试验地5 cm处土壤温度用纽扣式温度传感器(DS1921-F5#,Maxim/Dallas Semiconductor Inc.,USA)连续监测,每2 h记录1次数据。
1.4 相关计算 1.4.1 土壤呼吸温度敏感性及土壤呼吸通量土壤呼吸指数模型,Rs=aebT,Q10=e10b,式中Rs和T分别为土壤呼吸速率和土壤温度,a和b都是拟合参数,Q10为土壤呼吸速率对温度的敏感性。
两种处理测定的5 cm土壤温度T0与试验地连续监测的5 cm土壤温度Ti有显著的线性相关性(Ti=aT0+b,NL: R2=0.94,P<0.0001; CK: R2=0.95,P<0.0001)。将非观测日试验地连续土壤温度Ti代入方程Rs=aebT可得非观测日土壤呼吸速率Rsi;观测日Rsi为对应处理每2 h的土壤呼吸速率平均值。每日土壤呼吸(CO2-C排放量)的计算公式为: Di= (Rs1+ Rs2+ Rs3+……+ Rs12) ×3600×2×12×10-6 式中,Di为每日土壤呼吸(gC/m2),Rsi为每日2 h土壤呼吸速率(μmol m-2 s-1)。12为CO2-C的摩尔质量(g/mol),3600、2和10-6均为换算系数。最后,每日土壤呼吸速率累加得年累计土壤呼吸。
1.4.2 凋落物对土壤呼吸的贡献对照与去除凋落物处理日土壤呼吸速率观测值的差异及其与对照比值作为地表凋落物的贡献,用于评价地表凋落物贡献的季节变异性:
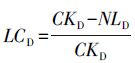
所有的统计分析在SPSS 17.0软件中进行。采用Repeated measures ANOVE进行方差分析和多重比较分析处理之间土壤温度、水分和土壤呼吸差异性;采用Student-t检验各时期各处理间土壤呼吸速率的差异。采用Pearson相关系数评价5 cm土壤水分和土壤呼吸的相关性,利用指数回归曲线拟合土壤呼吸速率与土壤温度的关系。
2 结果与分析 2.1 凋落物去除对土壤温度和水分的影响试验期间,凋落物去除和对照处理5 cm土壤温度变化规律基本一致,均表现为先下降后升高再下降的变化趋势,且存在显著的季节变化(P<0.01)。但两种处理之间5 cm土壤温度的差异性不显著(P>0.05,图 1)。去除凋落物和对照处理的土壤温度最大值分别为14.75 ℃和14.35 ℃,均出现在8月,最小值分别为-0.44 ℃和-0.35 ℃,均出现在1月底,全年平均温度分别为5.27 ℃和5.21 ℃。

|
| 图1 川西亚高山粗枝云杉人工林去除凋落物 (NL) 和对照 (CK) 处理下5 cm土壤温度和水分动态变化 Fig.1 Dynamics of soil temperature and soil water content at 5 cm depth of different treatments with litter exclusion and control in a subalpine dragon spruce (Picea asperata) plantation of western Sichuan NL和CK 分别表示凋落物去除(no litter)和对照(control) |
粗枝云杉人工林凋落物去除和对照处理下5 cm土壤水分相对土壤温度变化较为复杂,但两种处理的土壤水分变化存在相似的曲线格局,且季节性变化显著(P<0.01,图 1)。分析表明,两种处理之间5 cm土壤水分差异不显著(P>0.05)。凋落物去除和对照处理土壤水分分别在2.65%—44.50%和3.31%—45.14%范围变动,最高值和最低值均分别出现在6月和1月,平均值分别为26.48%和27.79 %。凋落物去除使土壤水分降低了4.95%。
2.2 凋落物去除对土壤呼吸速率的影响在季节动态上,凋落物去除和对照两种处理的土壤呼吸速率与以上分析的土壤温度变化规律基本一致,具有相似的‘S’型曲线格局,土壤呼吸速率存在显著季节性变化(图 2)。二者土壤呼吸速率变化范围分别为0.35—4.39 μmol m-2 s-1和0.40—5.15 μmol m-2 s-1,年均土壤呼吸速率分别1.99 μmol m-2 s-1和2.32 μmol m-2 s-1。总体上来看,去除凋落物的土壤呼吸速率比对照处理土壤呼吸速率低14.21%,且二者之间土壤呼吸速率的差异表现为生长季节明显高于非生长季节。统计分析表明,二者之间土壤呼吸速率差异不显著(P>0.05),但在去年的11月和翌年的5月差异显著(P<0.05)。

|
| 图2 川西亚高山粗枝云杉人工林去除凋落物和对照处理下土壤呼吸速率季节动态 Fig.2 Dynamics of soil respiration rate at 5 cm depth of different treatments with no litter and control in a subalpine dragon spruce (Picea asperata) plantation of western Sichuan NL和CK 分别表示凋落物去除(no litter)和对照(control),*表示差异显著 |
凋落物去除明显降低了土壤呼吸速率,且凋落物对土壤呼吸速率的贡献存在显著的季节性变化(P<0.05),变化范围为6.68%—25.52%(图 3)。在凋落物去除处理中,第1个月对土壤呼吸的影响很大(可能是第一次凋落物去除处理使土壤受到较大的干扰或由其他误差造成),第2年5月次之,其他变化幅度较为平稳,影响最小的是第2年的11月。另外,统计分析表明,凋落物去除和对照土壤呼吸年碳通量分别为761.41 gC/m2和875.15 gC/m2,凋落物对土壤呼吸的年均贡献率为14.93%(图 4)。

|
| 图3 凋落物去除对土壤呼吸速率影响的变化幅度 Fig.3 Percent changes in soil respiration induced by litter exclusion in a subalpine dragon spruce (Picea asperata) plantation of western Sichuan |

|
| 图4 凋落物去除和对照处理条件下土壤呼吸的年碳通量 Fig.4 Annual soil respiration between litter exclusion and control treatment |
回归分析表明,凋落物去除和对照处理的土壤呼吸速率与5 cm土壤温度之间存在显著指数关系(P<0.01,图 5)。凋落物去除和对照处理的5 cm土壤温度可分别解释土壤呼吸变异的70%和71%(表 1)。凋落物去除明显降低了土壤呼吸的温度敏感性(Q10),凋落物去除和对照处理的Q10分别为3.84和4.09(表 1)。相关分析表明,二者的土壤呼吸速率的对数值分别与5 cm土壤水分的对数值呈显著线性相关(P<0.05,图 6)。

|
| 图5 川西亚高山粗枝云杉人工林去除凋落物和对照处理下土壤呼吸与土壤温度的关系 Fig.5 Relationship between soil respiration and soil temperature at 5 cm depth of different treatments with litter exclusion and control in a subalpine dragon spruce (Picea asperata) plantation of western Sichuan |
| 处理 Treatment | a | b | Q10 | N | P | R2 |
| 去除凋落物No little | 0.84(0.2) | 0.13(0.003) | 3.84(0.15) | 55 | <0.001 | 0.703 |
| 对照Control | 0.71(0.1) | 0.14(0.009) | 4.09(0.25) | 55 | <0.001 | 0.708 |
| 数值=平均数(标准偏差) | ||||||

|
| 图6 川西亚高山粗枝云杉人工林去除凋落物和对照处理下土壤呼吸与土壤水分的关系 Fig.6 Relationship between soil respiration and soil moisture at 5 cm depth of different treatments with litter exclusion and control in a subalpine dragon spruce (Picea asperata) plantation of western Sichuan |
地表凋落物作为土壤生态系统独特的结构层次,是土壤有机质输入的主要来源,并成为土壤呼吸的一个重要组成部分[3, 4],而凋落物对土壤呼吸的贡献并不完全来源于凋落物自身分解释放的CO2,凋落物对土壤呼吸的贡献是一个非常复杂的生物学过程。森林系统中,通过去除凋落物可以改变土壤根系和微生物所需有机碳的供应而影响土壤呼吸。有研究表明,地表凋落物量与土壤呼吸速率呈极显著线性相关[18],去除凋落物会显著降低土壤呼吸速率[19, 20, 21]。凋落物去除明显降低了川西亚高山粗枝云杉人工林土壤呼吸速率,但差异不显著。这与王光军等[9]和Sulzman等[4]的研究结果一致。去除凋落物对土壤呼吸速率的影响总体上不显著,但去除凋落物所导致的土壤呼吸速率下降幅度均是生长季节大于非生长季节。这可能是该林区凋落物储量和分解主要与林木生长等因素有关,其次受温度和水分等因子的调控[16]。分析表明,凋落物对土壤呼吸速率的年均贡献率为14.93%,接近于陈光水等分析国内62个森林样地土壤呼吸及其相关因子数据得出凋落物对土壤呼吸的贡献率为20.2%[22],明显低于Raich等测定的凋落物对全球森林生态系统中土壤呼吸速率的平均贡献率为33%[18]。这说明凋落物对土壤呼吸的贡献率存在明显的空间变异。这可能主要是因为,云杉凋落物质量较低,可分解性不高;另外,云杉凋落物物理保护(云杉针叶外壳坚硬)使其很难被微生物直接利用[23],针叶凋落物往往含有更多难分解碳组分[24];再者,高海拔、低温度、低气压环境条件不利于凋落物的分解,因此粗枝云杉凋落物层对土壤呼吸的贡献相对较小。
凋落物对土壤呼吸的贡献机制是一个非常复杂的生物学过程,主要通过自身的分解和凋落物分解过程中可溶性有机物质通过淋溶进入土壤对土壤有机质分解产生影响[10, 19],一方面作为真菌或微生物生命代谢活动的物质基础[25],促进土壤微生物呼吸[26]。去除凋落物可以通过减少土壤养分的有效性或改变土壤微环境来影响微生物数量及其活性,进而降低土壤呼吸速率。且有研究表明,去除凋落物会引起土壤微生物生物量和活性降低[7]。另一方面可作为养分促进植物的萌发和生长[27, 28],使植物通过光合作用使可利用的土壤有机碳底物数量增加以及植物根系的伸长生长,进而促进土壤有机碳的化学氧化和植物根系呼吸,凋落物去除势必会降低根系呼吸,进而使土壤呼吸下降。本研究中,凋落物对土壤呼吸的贡献率相对较低,甚至有研究表明凋落物对土壤呼吸的贡献出现负值[11]。这可能与凋落物数量、土地利用方式、土壤水热因子有关,也可能取决于凋落物分解层自身释放的CO2与凋落物层对土壤呼吸CO2排放的屏蔽作用之间的平衡结果[11, 29]。
3.2 凋落物对土壤呼吸水热因子的影响温度是影响土壤呼吸的一个非常重要环境因子[30],主要通过影响微生物活性及植物根系生长进而影响土壤呼吸[9]。本研究表明,川西亚高山粗枝云杉人工林两种处理土壤呼吸速率同样受控于土壤温度,并均与5 cm土壤温度呈显著的指数相关关系,其变异系数约71%,这与前人的研究结果一致[4, 13]。凋落物对土壤温度的影响主要通过阻隔土壤与外界空气的热交换,抑制土壤降温(保温),且凋落物量越多对土壤温度的影响越大[31];反之,温度越高,凋落物分解的越快,则向土壤输入有机碳越多,土壤呼吸速率越高。本研究中,凋落物去除使土壤温度升高了1.2%,这与周小刚[14]等的研究结果一致,而王光军等[9]研究却指出凋落物去除降低了杉木人工林土壤温度。这可能与凋落物去除使土壤直接裸露被太阳直射有关。此外,由于土壤呼吸速率与温度指数呈显著正相关,凋落物去除使土壤温度升高,而实际观测的结果却是凋落物去除使土壤呼吸速率降低,这可能由于凋落物去除使土壤温度发生的变化对土壤呼吸速率的影响很小[9],又或者是土壤呼吸速率在一定程度上还受到其他因子的综合调控。
水分是影响土壤呼吸速率的另一关键环境因子,主要通过影响根系和微生物的生理过程以及底物和氧气的扩散进而调控土壤呼吸。通常认为,水分对土壤呼吸的影响较为复杂。有研究表明,只有当水分过高或过低才会对土壤呼吸有显著影响[32, 33]。本研究中,两种处理中土壤水分的对数值均与土壤呼吸速率呈显著线性正相关,这主要与川西亚高山森林明显的季节动态特征有关。而凋落物去除使土壤水分降低了4.95%,这与前人研究结果相符[9, 14]。因凋落物具有吸持、拦截水量,减少地表蒸发,改善土壤结构等作用[34],而凋落物去除势必会使土壤含水量下降,不利于可矿化底物的扩散和微生物生命代谢活动,导致土壤呼吸速率降低。
此外,本研究发现,凋落物去除和对照两种处理的土壤呼吸均具有显著的季节变化规律,并呈现相似的曲线格局,且凋落物对土壤呼吸速率的贡献也具有明显的季节变化。有研究表明,这主要受控于与季节变化明显相关的土壤水热因子[12, 20],且本研究已证实土壤呼吸速率与土壤温度、水分显著相关,从而使土壤呼吸具有明显的季节变异。
3.3 凋落物去除对Q10的影响Q10值是衡量土壤呼吸对温度变化响应敏感程度的一个重要指数[35],与气候变暖密切相关,对预测未来气候变化下的土壤碳平衡具有重要意义[36, 37]。土壤呼吸中组分不同,表现出对温度的敏感性不同[13]。本研究发现,凋落物去除使土壤呼吸温度敏感性Q10降低,与邓琦等[12]和Boone等[38]的研究结果一致。首先,由于凋落物是土壤生态系统独特的结构层次,覆盖于土壤表面,对土壤具有可塑性和缓冲作用,能降低土壤温度受气温波动的影响,进而减缓土壤环境对外界环境尤其是全球变暖响应的敏感性[12],反之,凋落物去除使土壤对外界环境变化的抵抗能力减弱,土壤温度敏感性降低;其次,凋落物去除使土壤水分降低,而土壤水分下降,不利于可矿化土壤碳底物的扩散,并降低底物的可利用性,从而降低土壤呼吸温度敏感性;最后,凋落物去除可通过直接或间接的途径使土壤有机碳输入减少,而通常情况下,Q10与底物的可利用性成正比[37],进而使土壤温度敏感性降低。此外,本研究还发现,凋落物去除处理和对照的Q10值分别为3.84和4.09,明显高于中国森林土壤呼吸Q10的平均值2.65[22],Erland 等指出全球Q10值变化范围为1.8—4.1[39]。这在一定程度上说明了川西亚高山海拔高、温度低,土壤呼吸的Q10值偏高,与陈宝玉等[40]研究结果一致。其森林土壤可能对未来气候变暖更为敏感。
综上所述,川西亚高山粗枝云杉人工林土壤呼吸具有显著的季节变化规律,与土壤温度呈显著指数相关,与土壤水分对数值呈显著线性相关。短期去除凋落物使土壤水分、土壤呼吸速率、土壤温度敏感性(Q10)均明显降低。本研究只有1年的试验数据,具有一定的局限性,凋落物去除对土壤呼吸的机制(特别是微生物方面)还有待进一步研究。
| [1] | 杨万勤, 邓仁菊, 张健. 森林凋落物分解及其对全球气候变化的响应. 应用生态学报, 2007, 18(12): 2889-2895. |
| [2] | Raich J W, Schlesinger W H. The global carbon dioxide flux in soil respiration and its relationship to vegetation and climate. Tellus B: Chemical and Physical Meteorology, 1992, 44(2): 81-99. |
| [3] | Wang Y D, Wang H M, Ma Z Q, Wen X F, Li Q K, Liu Y F, Sun X M, Yun G R. Contribution of aboveground litter decomposition to soil respiration in a subtropical coniferous plantation in Southern China. Asia Pacific Journal of Atmospheric Science, 2009, 45(2): 137-147. |
| [4] | Sulzman E W, Brant J B, Bowden R D, Lajtha K. Contribution of aboveground litter, belowground litter, and rhizosphere respiration to total soil CO2 efflux in an old growth coniferous forest. Biogeochemistry, 2005, 73(1): 231-256. |
| [5] | Prescott C E. Do rates of litter decomposition tell us anything we really need to know? Forest Ecology and Management, 2005, 220(1-3): 66-74. |
| [6] | Wan S Q, Norby R J, Ledford J, Weltzin J F. Responses of soil respiration to elevated CO2, air warming, and changing soil water availability in a model old-field grassland. Global Change Biology, 2007, 13(11): 2411-2424. |
| [7] | Li Y Q, Xu M, Sun O J, Cui W C. Effects of root and litter exclusion on soil CO2 efflux and microbial biomass in wet tropical forests. Soil Biology and Biochemistry, 2004, 36(12): 2111-2114. |
| [8] | Holub S M, Lajtha K, Spears J D H. Organic matter manipulations have little effect on gross and net nitrogen transformations in two temperate forest mineral soils in the USA and central Europe. Forest Ecology and Management, 2005, 214(1-3): 320-330. |
| [9] | 王光军, 田大伦, 闫文德, 朱凡, 项文化, 梁小翠. 改变凋落物输入对杉木人工林土壤呼吸的短期影响. 植物生态学报, 2009, 33(4): 739-747. |
| [10] | 武传胜, 沙丽清, 张一平. 哀牢山中山湿性常绿阔叶林凋落物对土壤呼吸及其温度敏感性的影响. 东北林业大学学报, 2012, 40(6): 37-40. |
| [11] | 王丽丽, 宋长春, 郭跃东, 刘德燕, 杨桂生. 三江平原不同土地利用方式下凋落物对土壤呼吸的贡献. 环境科学, 2009, 30(11): 3130-3135. |
| [12] | 邓琦, 刘世忠, 刘菊秀, 孟泽, 张德强. 南亚热带森林凋落物对土壤呼吸的贡献及其影响因素. 地球科学进展, 2007, 22(9): 967-986. |
| [13] | 王光军, 田大伦, 闫文德, 朱凡, 李树战. 去除和添加凋落物对枫香和樟树林土壤呼吸的影响. 生态学报, 2009, 29(2): 643-652. |
| [14] | 周小刚, 郭胜利, 车升国, 张芳, 邹彦军, 南雅芳, 李泽, 蒲辉. 黄土高原刺槐人工林地表凋落物对土壤呼吸的贡献. 生态学报, 2012, 32(7): 2150-2157. |
| [15] | Yang W Q, Wang K Y, Kellomäki S, Zhang J. Annual and monthly variations in litter macronutrients of three subalpine forests in western China. Pedosphere, 2006, 16(6): 788-798. |
| [16] | 刘庆, 吴彦, 陈庆恒. 亚高山针叶林生态学研究. 成都: 四川大学出版社, 2002. |
| [17] | 林波, 刘庆, 吴彦, 何海, 庞学勇. 川西亚高山人工针叶林枯枝落叶及苔藓层的持水性能. 应用与环境生物学报, 2002, 8(3): 234-238. |
| [18] | Raich J W, Nadelhoffer K J. Belowground carbon allocation in forest ecosystems: global trends. Ecology, 1989, 70(5): 1346-1354. |
| [19] | Subke J A, Hahn V, Battipaglia G, Linder S, Buchmann N, Cotrufo M F. Feedback interactions between needle litter decomposition and rhizosphere activity. Oecologia, 2004, 139(4): 551-559. |
| [20] | Rey A, Pegoraro E, Tedeschi V, De Parri I, Jarvis P G, Valentini R. Annual variation in soil respiration and its components in a coppice oak forest in Central Italy. Global Change Biology, 2002, 8(9): 851-866. |
| [21] | Ngao J, Epron D, Brechet C, Granier A. Estimating the contribution of leaf litter decomposition to soil CO2 efflux in a beech forest using 13C-depleted litter. Global Change Biology, 2005, 11(10): 1768-1776. |
| [22] | 陈光水, 杨玉盛, 吕萍萍, 张亿萍, 钱小兰. 中国森林土壤呼吸模式. 生态学报, 2008, 28(4): 1748-1761. |
| [23] | Shirato Y, Yokozawa M. Acid hydrolysis to partition plant material into decomposable and resistant fractions for use in the rothamsted carbon model. Soil Biology and Biochemistry, 2006, 38(4): 812-816. |
| [24] | Rustad L E, Fernandez I J. Soil warming: consequences for foliar litter decay in a Spurce-fir forest in Maine, USA. Soil Science Society of America Journal, 1998, 62(4): 1072-1080. |
| [25] | 张东秋, 石培礼, 张宪洲. 土壤呼吸主要影响因素的研究进展. 地球科学进展, 2005, 20(7): 778-785. |
| [26] | Lajtha K, Crow S E, Yano Y, Kaushal S S, Sulzman E, Sollins P, Spears J D H. Detrital controls on soil solution N and dissolved organic matter in soils: a field experiment. Biogeochemistry, 2005, 76(2): 261-281. |
| [27] | Matsushima M, Chang S X. Vector analysis of understory competition, N fertilization, and litter layer removal effects on white spruce growth and nutrition in a 13-year-old plantation. Forest Ecology and Management, 2006, 236(2-3): 332-341. |
| [28] | Haeussler S, Kabzems R. Aspen plant community response to organic matter removal and soil compaction. Canadian Journal of Forest Research, 2005, 35(8): 2030-2044. |
| [29] | 陈四清, 崔骁勇, 周广胜, 李凌浩. 内蒙古锡林河流域大针茅草原土壤呼吸和凋落物分解的CO2排放速率研究. 植物学报, 1999, 41(6): 645-650. |
| [30] | Luo Y Q, Wan SQ, Hui D F, Wallace L L. Acclimatization of soil respiration to warming in a tall grass prairie. Nature, 2001, 413(6856): 622-625. |
| [31] | 潘开文, 何静, 吴宁. 森林凋落物对林地微生境的影响. 应用生态学报, 2004, 15(1): 153-158. |
| [32] | Davidson E A, Verchot L V, Cattânio J H, Ackerman I L, Carvalho J E M. Effects of soil water content on soil respiration in forests and cattle pastures of eastern Amazonia. Biogeochemistry, 2000, 48(1): 53-69. |
| [33] | Maier C A, Kress L W. Soil CO2 evolution and root respiration in 11 year-old loblolly pine (Pinus taeda) plantations as affected by moisture and nutrient availability. Canadian Journal of Forest Research, 2000, 30(3): 347-359. |
| [34] | Atkin O K, Edwards E J, Loveys B R. Response of root respiration to changes in temperature and its relevance to global warming. New Phytologist, 2000, 147(1): 141-154. |
| [35] | Ohashi M, Gyokusen K, Saito A. Contribution of root respiration to total soil respiration in a Japanese cedar (Cryptomeria japonica D. Don) artificial forest. Ecological Research, 2000, 15(3): 323-333. |
| [36] | 周萍, 刘国彬, 薛萐. 草地生态系统土壤呼吸及其影响因素研究进展. 草业学报, 2009, 18(2): 184-193. |
| [37] | 杨庆朋, 徐明, 刘洪升, 王劲松, 刘丽香, 迟永刚, 郑云普. 土壤呼吸温度敏感性的影响因素和不确定性. 生态学报, 2011, 31(8): 2301-2311. |
| [38] | 陈宝玉, 刘世荣, 葛剑平, 王辉, 常建国, 孙甜甜, 马姜明, 施恭暕. 川西亚高山针叶林土壤呼吸速率与不同土层温度的关系. 应用生态学报, 2007, 18(6): 1219-1224. |
| [39] | Bååth E, Wallander H. Soil and rhizosphere microorganisms have the same Q10 for respiration in a model system. Global Change Biology, 2003, 9(12): 1788-1791. |
| [40] | Chen B Y, Liu S R, Ge J P, Wang H, Chang J G, Sun T T, Ma J M, Shi G J. The relationship between soil respiration and the temperature at different soil depths in sub-alpine coniferous forest of western Sichuan Province. Chinese Journal of Applied Ecology, 2007, 18(6): 1219-1224. |