 2015, Vol. 35
2015, Vol. 35文章信息
- 刘磊, 梁昌聪, 曾迪, 杨腊英, 覃和业, 王国芬, 郭立佳, 黄俊生
- LIU Lei, LIANG Changcong, ZENG Di, YANG Laying, QIN Heye, WANG Guofen, GUO Lijia, HUANG Junsheng
- 香蕉枯萎病田间分布型及病原菌在植株上的分布
- Spatial distribution pattern for the Fusarium wilt disease in banana field and the Fusarium oxyporum f. sp. cubense in different parts of banana plants
- 生态学报, 2015, 35(14): 4742-4753
- Acta Ecologica Sinica, 2015, 35(14): 4742-4753
- http://dx.doi.org/10.5846/stxb201311162741
-
文章历史
- 收稿日期:2013-11-16
- 修订日期:2014-12-04






2. 海南大学儋州校区应用科技学院, 儋州 571737;
3. 热作两院种苗组培中心, 儋州 571737
2. College of Applied Science and Technology, Hainan University, Danzhou 571737, China;
3. Seedlings Cultivation Center, Chinese Academy of Tropical Agricultural Sciences, Danzhou 571737, China
植物病害流行是病害在时空两个方面的扩展过程,空间扩散作用导致大田病株产生一定的分布类型[1, 2, 3]。作为种群的重要生态学属性之一,空间分布由物种的遗传恃性和生态环境条件所决定[4, 5, 6],其分布型可分为随机分布(可用泊松分布理论公式表示)[7]、聚集分布(可用负二项分布、奈曼分布等理论公式表示)[8, 9, 10]、均匀分布(可用二项分布理论公式表示)[14, 15]3种[16]。国内外学者多从空间图式角度研究聚集指标(平均拥挤度m*[17]、Bell扩散系数C、丛生指数I、Lloyd′s聚集度指数m*/m、久野Ca指标CA、负二项分布K值[18]等),建立空间分布图式、抽样技术研究等的数学模型[10, 14, 16, 19, 20]。前人已经对水稻纹枯病[21]、稻瘟病[22]、黑粉病[23]、紫鞘病[24],小麦白粉病[25]、纹枯病[26]、散黑穗病[27],小白菜花叶病[3]、棉花黄萎病[28]、番木瓜环斑病[29]、胡麻枯萎病[30]、香蕉束顶病[31]、杨树冰核细菌溃疡病[5]、草坪草条锈病[32]、白三叶草病毒病[33]、番茄花叶病[34]、蚕豆枯萎病[35]、西瓜枯萎病[6]、芦笋茎枯病[36]、西葫芦灰霉病[37]、白菜霜霉病及软腐病[38]、番茄黄化曲叶病[39]、玉米粗缩病[40]、油菜菌核病[41, 42]等病害的大田病株空间分布结构及抽样技术进行了研究。
香蕉枯萎病,又称巴拿马病、黄叶病,是由于尖孢镰刀菌古巴专化型(Fusarium oxyporum f. sp. cubense)破坏植株维管束导致植株死亡的一种毁灭性病害,目前,除地中海、印度洋、南太平洋一些岛屿国家外,几乎所有香蕉种植区均有遭受该病害危害的报道[43, 44, 45]。在我国广东[46]、广西[47]、福建、海南、云南和台湾等主要香蕉种植区也出现该病害,且发病加重趋势明显[48, 49]。该病害大田病株的空间分布型及抽样技术相关研究尚未见有报导,为此,本研究探讨了该典型土传病害大田植株的空间分布型及植株体内尖孢镰刀菌的分布情况,以期为香蕉枯萎病的监测检测和综合防控提供理论依据。
1 材料与方法 1.1 大田病株调查2013年8月在海南省儋州市中国热带农业科学院环境与植物保护研究所铺仔基地随机选择发病程度不同的6块田进行调查,每块田面积约为1334m2,种植香蕉品种为巴西蕉(Musa AAA Cavendish subgroup ‘Brazil’),对每块田进行棋盘式随机取样(根据地形取横向6个点,纵向10个点),以单株香蕉为样方,共取60株,分别记录枯萎病大田病株的株数和发病情况(表 1),计算病情指数。
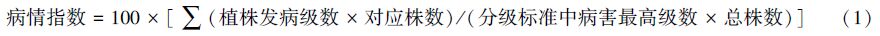
| 发病级别Grade | 分级标准Standards |
| 0 | 植株健康,无病状 |
| 1 | 植株球茎被枯萎病菌侵染,25%以内褐变(≤25%),但仍可正常生长 |
| 2 | 植株球茎被枯萎病菌侵染,25%—50%褐变(包括50%),半数叶片枯萎 |
| 3 | 植株球茎被枯萎病菌侵染,50%—75%褐变(包括75%),大部分叶片枯萎 |
| 4 | 植株球茎被枯萎病菌侵染,超过75%褐变(>75%),全株枯萎死亡 |
将田间调查所取得的数据分别进行二项分布、泊松分布、负二项分布、奈曼分布拟合。对实测频次分布和理论频次分布作X2适合性检验,根据是否适合确定属于何种分布概率类型[16, 50, 51]。

式中,o为观测值,c为理论值。
1.2.2 聚集度指标检验计算每块田的大田病株病情等级均值m、样本方差S2和平均拥挤度m*,根据Bell扩散系数C、丛生指标I、Lloyd′s聚集度指数m*/m、久野Cassie指标CA和负二项分布的K值来分析大田病株的聚集强度[16, 50, 51]。
1.2.3 Taylor幂法则Taylor分析了大量生物种群统计资料后,发现样本平均数m与方差S2的对数值之间存在回归关系公式[52]:

式中,α是一个与样本大小和计算方法有关的因子,受环境异质性的影响;β为种群聚集对密度依赖性的一个测度。lgα=0,β=1,随机分布;lgα>0,β=1,种群在一切密度下均是聚集分布,但不具聚集度对密度的依赖性;lgα>0,β>1,种群为聚集分布,但具聚集度对密度的依赖性;lgα<0,β<1,种群密度越高越均匀。
1.2.4 Iwao的m*-m模型及改进的m*-m回归分析法
当m*与m成线性关系时,a、b可揭示种群的分布特征。
a为分布的基本成分的平均拥挤度,a=0时分布的基本成分为单个个体;a>0时个体间相互吸引,分布的基本成分是个体群,即小聚集团;a<0时个体之间相互排斥。b为基本成分的空间分布型,b<1时为均匀分布,b=1时为随机分布,b>1时为聚集分布。

式中,α′是每个基本成分中个体数的分布的平均拥挤度,β′是在低密度下基本成分的相对聚集度,γ′+2δm是基本成分的相对聚集度随种群密度而变化的速率,2δ是基本成分的相对聚集度随种群密度变化的加速率。α′=0时分布的基本成分为单个个体;α′>0时个体间相互吸引,分布的基本成分是个体群,即小聚集团;α′<0时个体间相互排斥。β′<1时基本成分为均匀分布,β′=1时为随机分布,β′>1时为聚集分布。
| 聚集度指标 Aggregation index | 公式 Formulas | 判断标准 Standards | 备注 Remark |
| 平均拥挤度Mean crowing | m*=m+S2/m-1 | (3) | |
| 扩散系数Diffusion coefficient | C=S2/m | C<1时为均匀分布,C=1时为随机分布,C>1时为聚集分布 | (4) |
| 丛生指标Moore | I=S2/m-1 | I<0时为均匀分布,I=0时为随机分布,I>0时为聚集分布 | (5) |
| 聚集度指数Cluster index | m*/m=1+S2/m2-1/m | m*/m<1时为均匀分布,m*/m=1时为随机分布,m*/m>1时为聚集分布 | (6) |
| 久野Ca指标Cassie index | CA=(S2-m)/m2 | CA<0时为均匀分布,CA=0时为随机分布,CA>0时为聚集分布 | (7) |
| 负二项分布的K值Negative binomial distribution | K=m2/(S2-m) | K<0时为均匀分布,K→+∞时为随机分布,K>0时为聚集分布。 | (8) |

式中,λ为种群聚集均数,K为负二项分布的参数,γ为具有自由度为2K的X20.5分布的函数值,若2K为小数,可用图或比例的内插估值计算X20.5。当λ<2时,种群的聚集可能由某些环境作用所引起,而不是由于大田病株本身有聚集特性;当λ≥2时,种群的聚集是由于上述2个因素或其中的1个因素所引起的[16]。
1.3 抽样技术研究 1.3.1 最适理论抽样模型Iwao提出,当种群m*-m成线性关系时,可利用聚集度参数a、b来建立理论抽样数模型,确定不同病情等级、不同误差条件下的最适理论抽样数[53, 57]。

式中,n为最适抽样数,t为一定置信度p下的分布临界值(当p=95%时,t=1.96),a、b为m*-m回归模型中的截距、斜率值,D为允许相对误差(D=Sx/m)。
同理,改进的m*-m模型抽样公式为:
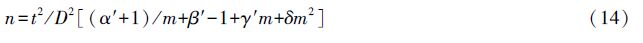
各参数同前述。
1.3.2 序贯抽样分析法模型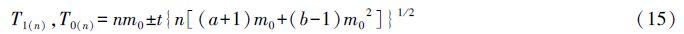
式中,T1(n),T0(n)为抽样结论的判据,a、b为m*-m回归模型中的截距、斜率值,m0为临界平均病情等级(防治指标),t为一定置信度p下的分布临界值(当p=95%时,t取1.96),n为累积抽样数。
1.3.3 抽样方法比较分别应用五点、单对角线、双对角线、平行线、棋盘式、“Z”字型6种方法进行抽样,采用曼-惠特尼U测验比较各种取样方法的误差率,选择最佳抽样方法[30, 36, 60]。
1.4 数据代换统计分析中,数据符合泊松分布、奈曼分布和负二项分布时,方差与均数存在函数关系,需按其分布型进行资料代换,以稳定方差[6, 50]。
TayIor幂法则资料代换:

式中,X′为代换后数据,β为TayIor幂函数参数。
Iwao回归模型资料代换:

式中,X′为代换后数据,a、b为m*-m模型回归参数。
1.5 枯萎病菌在大田病株体内分布在不同发病程度(轻、中、重)的样本田块5、3、6随机选择健康株(0级)和病株(1—4级)各10株,用打孔取样器在不同部位(根、球茎、假茎中部、假茎顶端、老叶、心叶)的病斑处、病健交界处和健康组织分别取样,装入贴有标签的自封袋带回实验室。称取样品质量后用75%的酒精处理2s,之后经0.1%升汞处理2—3min,再用无菌水冲洗3遍,研磨后加入100mL无菌水,制成匀浆[61, 62, 63]。将梯度稀释的匀浆液用微量移液器接入9cm的PDA平板(含50μg/mL潮霉素和200μg/mL硫酸链霉素)上,每皿0.1mL,均匀涂布。在28℃培养箱中培养3d后,采用平板菌落计数法统计菌落形成单位(cfu),计算相应部位每克鲜重样品组织的平均含菌量。试验以健康株为对照,每个处理重复3次。
1.6 数据分析采用Excel进行数据整理和绘图,运用SAS9.0软件计算均值和方差,采用Duncan氏新复极差法进行多重比较。
2 结果与分析 2.1 样本田块病株发病情况所调查的6块田地发病级别均值范围为1.0667—1.7833,其中样本田块6、4病情较重,显著高于其它样地,其病情指数分别为44.58%、40.42%;样本田块5发病较轻,显著低于其它样地,病情指数为26.67%。建立病情指数(X)与发病级别均值m(Y)之间的线性最小二乘回归方程,检测其相关关系极显著,因此可应用此方程估算不同病情指数下的发病级别均值(表 3)。
| 样本田块 Field No. | 病情等级均值 Mean of grade/m | 病情指数/% Disease index | 回归方程 Regression equation | 相关系数R |
| 1 | 1.2667bc | 31.67bc | ||
| 2 | 1.4833b | 37.08b | ||
| 3 | 1.4167b | 35.42b | Y=-0.0013+4.0037X | R=0.9999 |
| 4 | 1.6167ab | 40.42ab | ||
| 5 | 1.0667c | 26.67c | ||
| 6 | 1.7833a | 44.58a | ||
| 同列数据后不同小写字母表示经Duncan氏新复极差法检验差异显著(P<0.05) | ||||
表 4结果表明:6组样本田地的频次分布数据均不符合二项分布,样本田块1、5同时符合负二项分布和奈曼分布,样本田块2仅符合奈曼分布,样本田块3、6同时符合泊松分布、负二项分布和奈曼分布,样本田块4同时符合泊松分布和奈曼分布。因此,香蕉枯萎病大田病株在田间的频次分布主要为聚集分布,田间表现为以发病株为核心,呈嵌纹状或放射状扩散。
| 样本田块 Field No. | 二项分布 Binomial distribution | 泊松分布 Poisson distribution | 负二项分布 Negative binomial distribution | 奈曼分布 Neyman distribution |
| 1 | <0.01* * | <0.01* * | 0.2341NS | 0.2503NS |
| 2 | <0.01* * | <0.01* * | 0.0425* | 0.1022NS |
| 3 | <0.01* * | 0.0810NS | 0.1531NS | 0.1685NS |
| 4 | <0.01* * | 0.1326NS | 0.0396* | 0.0861NS |
| 5 | <0.01* * | 0.0251* | 0.2813NS | 0.2927NS |
| 6 | <0.01* * | 0.2136NS | 0.3277NS | 0.3514NS |
| * *表示极不适合,P<0.01;*表示不适合,P<0.05;NS表示适合,P>0.05 | ||||
大田病株在6块样本田地中的平均拥挤程度或个体邻居数介于1.3538—1.9446之间,扩散系数C>1,丛生指数I>0,聚集度指数m*/m>1,久野Ca指标CA>0,负二项分布K>0,因此,其在田间的空间图式符合聚集分布。此外,根据负二项分布K的大小可比较聚集程度,样本田地1、2、5的聚集程度显著高于其它样地(表 5)。
| 样本田块 Field No. | 方差 Variance S2 | 平均拥挤度 Mean crowing m* | 扩散系数 Diffusion coefficient C | 丛生指数 Moore I | 聚集度指数 Cluster index m*/m | 久野Ca指标 Cassie index CA | 负二项分布 Negative binomial distribution K |
| 1 | 1.5548 | 1.4941 | 1.2274 | 0.2274 | 1.1796 | 0.1796 | 5.5693bc |
| 2 | 1.8133 | 1.7058 | 1.2225 | 0.2225 | 1.1500 | 0.1500 | 6.6672bc |
| 3 | 1.5692 | 1.5243 | 1.1076 | 0.1076 | 1.0760 | 0.0760 | 13.1609ab |
| 4 | 1.7658 | 1.7089 | 1.0922 | 0.0922 | 1.0570 | 0.0570 | 17.5300a |
| 5 | 1.3729 | 1.3538 | 1.2871 | 0.2871 | 1.2691 | 0.2691 | 3.7160c |
| 6 | 2.0709 | 1.9446 | 1.1613 | 0.1613 | 1.0904 | 0.0904 | 11.0576b |
大田病株的S2-m回归关系为lgS2=0.0916+0.9743lgm(R=0.9490,显著相关)。其中,lgα>0,β<1,且C>1,m>1,超出Taylor判据系统范围,参考沈佐锐、马占山、兰星平等[64, 65, 66]完善的判据系统,相关参数值表明大田病株趋于聚集分布,且聚集程度随着种群密度的升高而升高。
2.4.2 m*-m回归分析大田病株的m*-m回归关系为m*=0.2560+0.9774m(R=0.9648,显著相关)。其中,a>0,b<1,表明大田病株个体间相互吸引,空间分布的基本成分是个体群,即小聚集团,病株个体群在田间趋于均匀分布。
大田病株的m*-m改进模型为m*=20.9145-30.0477m+11.0962m2+0.0876m3(R复=0.8997,显著相关)。其中,α′>0,β′<1,表明大田病株个体间相互吸引,空间分布的基本成分是个体群,即小聚集团,病株个体群在田间趋于均匀分布。病株个体群的相对聚集度随种群密度而变化的速率为(11.0962+0.1752m),因此,病株个体群密度越大,相对聚集度随密度而变化的速率越大,其加速率为0.1752。
2.5 聚集原因分析计算出6块样本田地的λ值分别为1.1919、1.4080、1.3807、1.5811、0.9677、1.9717,均小于2,表明香蕉枯萎病大田病株的聚集是由环境作用所致,而不是因为其本身的聚集习性。
2.6 抽样模型分析 2.6.1 理论抽样模型将95%置信区间的概率保证值t=1.96代入最适理论抽样公式n=t2(1.2560/m-0.0226)/D2,可得出不同平均病情等级大田病株的理论抽样数。在相同平均病情等级的田块,随着允许误差值的增大,所需抽样数显著减少;在同一允许误差值时,平均病情等级较高的田块所需的抽样数显著少于平均病情等级较低的田块所需的抽样数(表 3,表 6)。
| 允许误差 Permissible variation D | 不同样本田块的理论抽样数Sampling number at different field (n)/株 | ||||||
| 1 | 2 | 3 | 4 | 5 | 6 | ||
| 0.1 | 372aB | 317aBC | 332aB | 290aBC | 444aA | 262aC | |
| 0.2 | 93bcB | 79bcBC | 83bcB | 72bcBC | 111bcA | 65bcC | |
| 0.3 | 41cB | 35cBC | 37cB | 32cBC | 49cA | 29cC | |
| 0.4 | 23cB | 20cBC | 21cB | 18cBC | 28cA | 16cC | |
| 同列数据后不同小写字母表示经Duncan氏新复极差法检验差异显著(P<0.05),同行数据后不同大写字母表示经Duncan氏新复极差法检验差异显著(P<0.05) | |||||||
当允许误差D=0.2时,所需抽样数适中,且误差较小(表 6),通过计算此时不同病情指数所对应的最适理论抽样数可知,当病情指数为5%时,所需抽样数为604株;当病情指数为25%时,所需抽样数为119株;当病情指数为45%时,所需抽样数为65株。可见,随着病情指数的增加,大田病株所需抽样数也相应减少(图 1)。

|
| 图1 不同病情指数所对应的理论抽样数 Fig.1 The optimum sampling number for each disease index |
按m*-m改进模型所得的相应抽样公式为n=t2(1.2560/m-0.0226+11.0962m+0.0876m2)/D2,在本研究中,在允许误差范围内利用该公式计算得出所需的理论抽样数较多,以样地1为例,其理论抽样数分别为5826(D=0.1)、1456(D=0.2)、647(D=0.3)、364(D=0.4),因此,从便于田间调查和保证误差的角度考虑,本研究采用Iwao理论抽样公式计算理论抽样数。
2.6.2 序贯抽样模型参考刘波等[6]提出的西瓜枯萎病防治指标m0为病情指数25%时的发病级别均值,计算本研究中m0=0.9996,将a、b、t、m0代入模型公式,得出判据上下限计算公式为:T1(n),T0(n)= 0.9996n±2.1763√n。据此可建立香蕉枯萎病序贯抽样检索表(表 7)。若累计病情等级超过上限可定为防治对象田,若累计病情等级低于下限可定为不防治田,若累计病情等级在上下限之间,则需增加抽样量继续调查,同时,以理论抽样模型中的最大抽样数终止抽样。
| T1(n) T0(n) | 调查株数Plant number (n)/株 | |||||||
| 10 | 20 | 30 | 40 | 50 | 60 | 70 | 80 | |
| 上限Upper limit | 16.8781 | 29.7247 | 41.9081 | 53.7481 | 65.3688 | 76.8335 | 88.1802 | 99.4334 |
| 下限Lower limit | 3.1139 | 10.2593 | 18.0679 | 26.2199 | 34.5912 | 43.1185 | 51.7638 | 60.5026 |
| T1(n)T0(n) | 调查株数Plant number (n)/株 | |||||||
| 90 | 100 | 110 | 120 | 130 | 140 | 150 | ||
| 上限Upper limit | 110.6102 | 121.723 | 132.7812 | 143.7922 | 154.7616 | 165.6943 | 176.5941 | |
| 下限Lower limit | 69.3178 | 78.197 | 87.1308 | 96.1118 | 105.1344 | 114.1937 | 123.2859 | |
| T1(n) T0(n) | 调查株数Plant number (n)/株 | |||||||
| 160 | 170 | 180 | 190 | 200 | 210 | 220 | 230 | |
| 上限Upper limit | 187.4643 | 198.3075 | 209.1261 | 219.9222 | 230.6975 | 241.4536 | 252.1917 | 262.9132 |
| 下限Lower limit | 132.4077 | 141.5565 | 150.7299 | 159.9258 | 169.1425 | 178.3784 | 187.6322 | 196.9028 |
| T1(n) T0(n) | 调查株数Plant number (n)/株 | |||||||
| 240 | 250 | 260 | 270 | 280 | 290 | 300 | ||
| 上限Upper limit | 273.6191 | 284.3103 | 294.9878 | 305.6523 | 316.3045 | 326.9451 | 337.5746 | |
| 下限Lower limit | 206.1889 | 215.4897 | 224.8042 | 234.1317 | 243.4715 | 252.8229 | 262.1854 | |
经曼-惠特尼U检测可得,6种抽样方法的u值均小于1.96,故这些取样方法均可用于抽样研究,进一步比较u值大小可知,棋盘式、单对角线、双对角线取样方式的误差率显著低于其它方法,可结合实际需要优先选择(表 8)。
| 抽样方法 Sampling methods | u | p | 抽样方法 Sampling methods | u | p |
| 五点Five points type | 1.9239c | >0.05 | 单对角线Single diagonal type | 0.9553a | >0.05 |
| 双对角线Double diagonal type | 0.9124a | >0.05 | 平行线Paralleism type | 1.3046b | >0.05 |
| 棋盘式Chessboard type | 0.8651a | >0.05 | “Z”字型"Z"type | 1.6587bc | >0.05 |
基于Taylor幂法则的大田病株的代换公式为:X′=X0.5129。基于Iwao回归模型的大田病株的代换公式为:X′=sin-1(0.0180X)1/2。
2.8 枯萎病菌在大田病株体内的分布不同发病程度样地的健康株和病株的心叶均未检测到病原菌,发病较轻的样本田块5未在病株老叶和假茎顶端检测到病原菌,发病程度中等的样本田块3未在老叶检测到病原菌,发病程度较重的样本田块6在老叶、假茎顶端均检测到病原菌,表明不同发病程度样地病株体内病原菌的分布有显著差异(图 2)。此外,在植株不同部位检测到的尖孢镰刀菌平均含量也存在显著差异,健康株和病株球茎的平均含菌量显著高于其它部位,病株根部的病原菌含量显著高于假茎顶部及叶部的病原菌含量,老叶仅检测到少量病原菌。健康株根部、假茎的病原菌平均含量均显著低于病株相应部位的病原菌平均含量。

|
| 图2 不同发病程度香蕉植株体内尖孢镰刀菌的平均含菌量测定 Fig.2 The result for the average amount of Fusarium oxyporum f. sp. cubense inside banana plants with disease index 不同字母表示经Duncan氏新复极差法检验差异显著(P<0.05) |
针对植物病害开展空间分布格局研究,有助于优化抽样技术,对了解病害流行的主导因子、消长动态、危害损失及其防控措施等至关重要。本文采用历来定量研究田间虫害[60, 67, 68, 69, 70, 71]及病害[3, 5, 6, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42]空间分布的统计生态学方法首次对香蕉枯萎病大田病株的空间分布型和抽样技术进行了分析,研究结果表明,除2号样地以外,其余样地的频次分布均符合两个或两个以上理论模型,因此频次分布仅能检验样地中病株的空间分布聚集与否,结合聚集度指标则可判断出每块样地中的病株均为聚集分布且可比较样地间的聚集程度,但仍不能确切得到大田病株的具体理论分布型,也不能反映全部样地大田病株的整体聚集或离散情况。Taylor幂法则和m*-m等基础性生态学模型是频次分布和聚集度指标在分析空间分布型方面的推广应用,在种群生态学研究中占有重要地位。但在实际应用中,Taylor幂法则仍存在不完备性,需进一步完善[64, 65, 66];Iwao的m*-m回归模型虽然克服了频次分布的缺陷,计算相对简单,但在实际拟合时,常出现非线性关系,徐汝梅、于秀林等[55, 56]先后对其进行了改进,改进后的模型可提供大量空间分布的信息,有助于研究枯萎病等病虫害的田间发病规律。
香蕉枯萎病大田病株的理论分布及空间图式均趋向于聚集分布,据此我们可判断该病害在大田中必然存在明显的发病中心,这与王振中、卓国豪等[44, 72]所述的香蕉枯萎病田间发病特点一致,刘晓光[5]、刘波[6]、敬甫松[21]等[22, 23, 27, 28, 30, 35, 36, 37]分别研究杨树冰核细菌溃疡病、西瓜枯萎病、水稻纹枯病等细菌性、真菌性土传病害时也发现其大田病株具有聚集分布的特征。形成这种聚集格局主要由环境作用所引起,跟大田病株本身的聚集习性无关,这与王家和等[35]对蚕豆枯萎病病株的聚集原因分析结果一致,推测环境作用表现为枯萎病菌通过田间病株残体、耕作工具、动物活动、带菌种苗、土壤和流水等传播,会“优先”侵染病株周周的健株,同时在同一田块的相邻地方,发病率也会有很大差异,符合王振中、魏岳荣、卓国豪等[3, 43, 44, 72]总结的香蕉枯萎病田间传播方式。
香蕉枯萎病是典型的土传病害,枯萎病菌是兼性寄生菌,腐生能力较强,在发病株及土壤中存活周期较长,特别是形成休眠孢子后,可存活数年[73]。目前,单一的防治措施难以有效控制该病害的发生,采用多种措施综合防控才可能有效降低香蕉枯萎病的发生,发病率较高的蕉园只能丢荒或改种其它作物,因此,在田间发现病株后,就应及时采取相关措施(另文发表),避免枯萎病菌以病株残体为中心向周围扩散蔓延。在这个过程中,抽样技术研究可帮助我们估计病虫害的田间种群密度,准确地进行预测和执行防治经济阀值,在建立作物-病虫害系统的控制与预测的数学模型及采取综合管理措施等方面意义重大。本研究的理论抽样数模型是将m*-m关系方程引入正态分布的理论抽样方程得到的,可在确定适宜抽样数量的同时保证抽样质量和节约抽样成本,主要用于种群资料积累,田间试验处理比较,病害预测,组建数学模型等。而序贯抽样(风险决策抽样),可在不影响结论准确性的前提下,尽量减少抽样数量,只对防治指标等进行统计检验,主要用于病害田间管理,检验病害的发生程度及防治标准等方面。研究时选择不受分布型限制的m*-m关系序贯抽样模型,以假设检验的方法,在一定的概率水平保证下,考虑接受或拒绝子样的调查结果,即将累积病情等级计数与抽样结论判据进行比较,根据判据的上下限得出结论,当结论不能确定时,以理论抽样方程计算的的最大理论抽样数终止抽样过程。对不同发病程度植株不同部位的取样分析表明尖孢镰刀菌在田间侵染香蕉极有可能是从其根部侵入,进入球茎后经维管束向假茎上部及老叶蔓延,与王振中、李春雨等[44, 72]所述的香蕉枯萎病菌侵染特点一致,当累积到一定含菌量后,引起植株病症出现。因此,无症状表现的健康株也有可能是处于潜育期状态的植株。此外,在侵染过程中,植株球茎含菌量显著高于其它部位,在症状上可明显观察到假茎基部开裂,解剖假茎基部可进一步观察到维管束堵塞褐变等。
植物病害的流行是病害在时间和空间两个方面的扩展过程,因此,应用统计生态学对香蕉枯萎病大田病株在时间上的依赖性与连续性的研究有待进一步完成。另外,植物病害流行的空间动态研究不仅包括病株的田间分布型,尚需进一步分析传播距离、传播速度等关键侵染特点,以加深对病害流行规律的认识。
致谢: 海南大学儋州校区应用科技学院2011级本科生贺江江、耿通云参与调查工作,特此致谢。
| [1] | Van der Plank J E. Plant Disease: Epidemics and Control. New York: Academic Press, 1963: 349-349. |
| [2] | Jeger M J. Analysing epidemics in time and space. Plant Pathology, 1983, 32(1): 5-11. |
| [3] | 王振中, 林孔勋, 范怀忠. 小白菜花叶病病株空间分布类型分析. 华南农业大学学报, 1989, 10(2): 48-53. |
| [4] | Taylor L R. Assessing and interpreting the spatial distributions of insect populations. Annual Review of Entomology, 1984, 29: 321-357. |
| [5] | 刘晓光, 项存悌, 董爱荣, 邵忠文, 王传伟, 林海波, 徐柏荣, 张景华, 尹宁. 杨树冰核细菌溃疡病株空间分布格局及其应用. 东北林业大学学报, 1999, 27(3): 36-39. |
| [6] | 刘波, 朱育菁, 肖荣凤, 林抗美, 冒乃和, 李芳, 苏明星, 史怀. 西瓜枯萎病病株空间分布格局及其抽样技术. 生态学报, 2004, 24(9): 2043-2049. |
| [7] | Garwood F. Fiducial limits for the poisson distribution. Biometrika, 1936, 28(3/4): 437-442. |
| [8] | Fisher R A. The negative binomial distribution. Annals of Eugenics, 1941, 11(1): 182-187. |
| [9] | Bliss C I, Fisher R A. Fitting the negative binomial distribution to biological data. Biometrics, 1953, 9(2): 176-200. |
| [10] | Taylor L R, Woiwod I P, Perry J N. The Negative binomial as a dynamic ecological model for aggregation, and the density dependence of k. The Journal of Animal Ecology, 1979, 48(1): 289-304. |
| [11] | Neyman J. On a new class of "contagious" distributions, applicable in entomology and bacteriology. The Annals of Mathematical Statistics, 1939, 10(1): 35-57. |
| [12] | Douglas J B, Fitting the Neyman type A (two parameter) contagious distribution. Biometrics, 1955, 11(2): 149-173. |
| [13] | Massé J C, Theodorescu R. Neyman type A distribution revisited. Statistica Neerlandica, 2005, 59(2): 206-213. |
| [14] | Edwards A W F. The meaning of binomial distribution. Nature, 1960, 186(4730): 1074-1074. |
| [15] | Altham P M E. Two generalizations of the binomial distribution. Journal of the Royal Statistical Society: Series C (Applied Statistics), 1978, 27(2): 162-167. |
| [16] | 丁岩钦. 昆虫数学生态学. 北京: 科学出版社, 1994: 22-42, 45-56. |
| [17] | Lloyd M. Mean crowding. Journal of Animal Ecology, 1967, 36(1): 1-30. |
| [18] | 韩铭哲. 关于负二项分布参数K值的讨论. 生物数学学报, 1989, 4(2): 97-101. |
| [19] | David F N, Moore P G. Notes on contagious distributions in plant populations. Annals of Botany, 1954, 18(1): 47-53. |
| [20] | Kuno E. Studies on population dynamics of rice leaf hopper in a paddy field. Bulletin of the Kyushu National Agricultural Experiment Station, 1968, (14): 131-246. |
| [21] | 敬甫松, 李兴谦, 徐昌能. 纹枯病株在杂交稻田的空间分布型的研究及其应用. 植物病理学报, 1983, 13(4): 27-35. |
| [22] | 穆永顺, 李刚. 稻瘟病叶瘟和穗颈瘟空间分布型及理论抽样研究. 西北农林科技大学学报: 自然科学版, 1987, 15(2): 77-83. |
| [23] | 杨志华, 刘康成, 周淑兰, 曾水根. 水稻粒黑粉病空间分布及抽样技术研究. 植物病理学报, 1994, 24(2): 185-192. |
| [24] | 肖满开. 水稻紫鞘病株空间分布型及理论抽样数量. 植物病理学报, 1994, 24(3): 250-250. |
| [25] | 刘逸卿, 汤其豹. 小麦白粉病田间分布型的初步研究. 植物保护学报, 1984, 11(2): 91-94. |
| [26] | 刘逸卿, 汤其豹. 小麦纹枯病株的空间分布型及其应用. 江苏农业学报, 1985, 1(2): 45-47. |
| [27] | 杨焱杰, 王贵生, 王永祥, 任千祥. 小麦散黑穗病空间分布型及抽样技术. 植物保护学报, 1995, 22(3): 256-258. |
| [28] | 王昆, 胡家琪, 狄文枝, 金芝芳, 王其友. 棉花黄萎病株空间格局及抽样技术研究. 植物病理学报, 1992, 22(2): 169-174. |
| [29] | 张德雍, 王振中, 范怀忠. 番木瓜环斑病的空间分布及流行动态模糊聚类分析. 华南农业大学学报, 1995, 16(4): 27-31. |
| [30] | 甘国福, 甘国君, 王发有, 王英兰. 胡麻枯萎病株田间分布型及抽样技术. 甘肃农业大学学报, 1996, 31(4): 395-398. |
| [31] | 周仲驹, 黄志宏, 郑国璋, 林奇英, 谢联辉. 香蕉束顶病的研究V. 病株的空间分布型及其抽样. 福建农业大学学报, 1997, 26(2): 177-181. |
| [32] | 赵兰勇, 刘振宇, 高祥伟, 季延平. 草坪草条锈病空间格局及抽样技术. 中国草地, 2001, 23(6): 39-41. |
| [33] | 梁巧兰, 魏列新, 徐秉良. 白三叶草病毒病病株空间分布型及抽样技术研究. 中国草地学报, 2013, 35(2): 48-51. |
| [34] | 张卫民, 王振中. 番茄花叶病病株及传毒介体空间分布型研究. 华南农业大学学报, 2002, 23(4): 23-26. |
| [35] | 王家和, 王崇德. 蚕豆枯萎病病株田间分布型及调查取样技术研究. 云南农业大学学报, 2003, 18(4): 343-345, 381-381. |
| [36] | 王向阳. 芦笋茎枯病病株空间分布型及其抽样研究. 安徽农业大学学报, 2004, 31(3): 344-347. |
| [37] | 张路生, 刘俊展, 刘庆年, 巴秀成. 保护地西葫芦灰霉病病株空间分布格局及其抽样技术. 石河子大学学报: 自然科学版, 2009, 27(2): 202-205. |
| [38] | 段显德, 马海霞, 杨信东. 白菜霜霉病及软腐病空间分布型研究. 吉林农业大学学报, 2010, 32(2): 130-135. |
| [39] | 李刚, 吴永汉, 陈文华, 都林娜, 许方程. 番茄黄化曲叶病病株及传毒介体烟粉虱的空间分布型. 浙江农业学报, 2010, 22(6): 779-783. |
| [40] | 赵敏, 杨廉伟, 王华弟, 李荣, 吴传伟, 张国忠, 祝小祥. 玉米粗缩病危害动态及空间格局和抽样技术的研究. 中国农学通报, 2012, 28(12): 187-191. |
| [41] | 朱金良, 陈跃, 王光华, 王华弟, 祝增荣. 油菜菌核病轻发生条件下的空间分布格局与抽样技术. 浙江农业学报, 2012, 24(3): 464-468. |
| [42] | 徐森富, 王会福, 余山红, 汪恩国. 油菜菌核病病株空间分布格局及抽样技术研究. 科技通报, 2013, 29(3): 96-99. |
| [43] | 魏岳荣, 黄秉智, 杨护, 许林兵, 邱继水. 香蕉镰刀菌枯萎病研究进展. 果树学报, 2005, 22(2): 154-159. |
| [44] | 王振中. 香蕉枯萎病及其防治研究进展. 植物检疫, 2006, 20(3): 198-200. |
| [45] | Fourie G, Steenkamp E T, Gordon T R, Vilioen A. Evolutionary relationships among the Fusarium oxysporum f. sp. cubense vegetative compatibility groups. Applied and Environmental Microbiology, 2009, 75(14): 4770-4781. |
| [46] | 李敏慧, 习平根, 姜子德, 戚佩坤. 广东香蕉枯萎病菌生理小种的鉴定. 华南农业大学学报, 2007, 28(2): 38-41. |
| [47] | 莫贱友,郭堂勋,李焜华. 广西香蕉枯萎病的发生与病原鉴定. 广西农业科学, 2008, 39(4): 481-484. |
| [48] | 高乔婉. 香蕉枯萎病//中国农业百科全书编辑部. 植物病理学卷. 北京: 中国农业出版社, 1996: 482-482. |
| [49] | Dong Z Y, Wang Z Z. Isolation and characterization of an exopolygalacturonase from Fusarium oxysporum f.sp. cubense race 1 and race 4. BMC Biochemistry, 2011, 12: 51-51. |
| [50] | 徐汝梅. 昆虫种群生态学. 北京: 北京师范大学出版社, 1987: 8-14, 31-34. |
| [51] | 唐启义, 冯明光. 实用统计分析及其DPS数据处理系统. 北京: 科学出版社, 2002: 36-36, 114-150. |
| [52] | Taylor L R. Aggregation, variance and the mean. Nature, 1961, 189(4766): 732-735. |
| [53] | Iwao S. A new regression method for analyzing the aggregation pattern of animal populations. Researches on Population Ecology, 1968, 10(1):1-20. |
| [54] | Iwao S. Application of the M*-M method to the analysis of spatial patterns by changing the quadrat size method to the analysis of spatial patterns by changing the quadrat size. Researches on Population Ecology, 1972, 14(1): 97-128. |
| [55] | 徐汝梅, 刘来福, 丁岩钦. 改进的IWAO M-M模型. 生态学报, 1984, 4(2): 111-118. |
| [56] | 于秀林, 任朝佐. 再次改进的IWAO M-M模型. 生态学报, 1986, 6(3): 193-196. |
| [57] | Iwao S. An approach to the analysis of aggregation pattern in biological populations. Statistical Ecology, 1971, 1: 461-513. |
| [58] | Iwao S. A new method of sequential sampling to classify populations relative to a critical density. Researches on Population Ecology, 1975, 16(2): 281-288. |
| [59] | Iwao S. Relation of frequency index to population density and distribution pattern. Physiological Ecology Japan, 1976, 17: 457-464. |
| [60] | 峗薇, 杨茂发, 廖启荣, 杨洪, 陈文龙. 两种育秧方式下稻水象甲幼虫的空间分布型及其抽样技术. 植物保护学报, 2013, 40(2): 128-132. |
| [61] | 方中达. 植病研究方法. 北京: 中国农业出版社, 1998: 122-125. |
| [62] | 肖荣凤, 刘波, 林抗美, 郑雪芳, 蓝江林. 斑叶露兜树茎腐病病原鉴定及植株体内菌量测定. 园艺学报, 2009, 36(2): 251-256. |
| [63] | 苏明星, 肖荣凤, 朱育菁, 黄素芳, 刘波. 苦瓜枯萎病病原的鉴定及植株体内菌量测定. 中国蔬菜, 2010, (16): 62-66. |
| [64] | 沈佐锐. 关于Taylor幂法则的统计学讨论. 生态学杂志, 1990, 9(6): 64-67. |
| [65] | 马占山. Taylor幂法则的进一步解释与种群聚集临界密度. 青年生态学者论丛(一), 1991, 1: 284-288. |
| [66] | 兰星平. 关于Taylor幂法则的讨论. 林业科学, 1996, 32(1): 86-91. |
| [67] | 刘缠民, 廉振民. 棉田棉蝗卵块空间分布型及抽样技术研究. 西北农业学报, 2003, 12(1): 136-138. |
| [68] | 卢辉, 刘奎, 钟义海, 赵冬香, 尹炯. 海南香蕉根颈象甲卵的空间分布型及理论抽样数确定. 热带作物学报, 2009, 30(7): 995-999. |
| [69] | 张总泽, 刘双平, 张静, 罗礼智, 江幸福. 向日葵螟幼虫的空间分布型及抽样技术. 昆虫知识, 2010, 47(4): 720-724. |
| [70] | 沈平, 刘萍, 吴建华, 常承秀, 朱惠英, 邓刚. 甘肃狭啮卵在花椒园中的空间分布型及抽样技术. 中国生物防治学报, 2012, 28(3): 435-438. |
| [71] | 张聪, 葛星, 赵磊, 王振营, 何康来, 白树雄. 双斑长跗萤叶甲越冬卵在玉米田的空间分布型. 生态学报, 2013, 33(11): 3452-3459. |
| [72] | 卓国豪, 黄有宝, 吴运新, 潘礼增, 冯伟扬, 刘绍钦. 香蕉枯萎病的综合防治技术. 植物检疫, 2003, 17(5): 279-280. |
| [73] | Li C Y, Chen S, Zuo C W, Sun Q M, Ye Q, Yi G J, Huang B Z. The use of GFP-transformed isolates to study infection of banana with Fusarium oxysporum f. sp. cubense race 4. European Journal of Plant Pathology, 2011, 131(2): 327-340. |