 2015, Vol. 35
2015, Vol. 35文章信息
- 赵瑞一, 吕现福, 蒋建建, 段逸凡
- ZHAO Ruiyi, LÜ Xianfu, JIANG Jianjian, DUAN Yifan
- 土壤CO2及岩溶碳循环影响因素综述
- Factors affecting soil CO2 and karst carbon cycle
- 生态学报, 2015, 35(13): 4257-4264
- Acta Ecologica Sinica, 2015, 35(13): 4257-4264
- http://dx.doi.org/10.5846/stxb201412112460
-
文章历史
- 收稿日期:2014-12-12
- 修订日期:2015-01-27






随着全球气候变暖和极端气候的频频发生,CO2的排放和运移日益受到重视。碳酸盐岩作为全球最大的碳库,深入研究岩溶作用对揭示自然界碳循环规律和全球碳减排具有重要意义[1]。尽管碳酸盐岩溶蚀消耗的CO2最终会因碳酸盐在海洋中沉积而被释放到大气中,岩溶作用也被认为只是CO2的转移而不是大气CO2的汇[2]。但由于快速的岩溶作用以及碳酸盐岩较高的可溶蚀性,其对大气CO2浓度的调节作用不容忽视[3-5]。另外,岩溶作用消耗的CO2以溶解无机碳(DIC)的形式被河流运移到海洋中,而在运移过程中,DIC被河流中水生植物消耗并使其转化为有机碳而储存在植物体中,从而增强了岩溶的碳汇效应[6-7]。
土壤与大气之间的碳通量对调节大气CO2浓度具有重要意义。植物通过光合作用固定大气中的碳,并且植物残体会以有机碳的形式进入到土壤中。同时,植物的呼吸作用以及土壤有机质的分解也会向大气中释放CO2,从而保持大气与土壤之间的碳收支平衡。但是岩溶作用通过消耗土壤CO2,导致土壤向大气释放的CO2量减少,虽然这并没有直接消耗大气中的CO2,但对大气CO2起到了减源的作用,因此岩溶作用对土壤CO2的消耗也可认为是大气CO2一个重要的汇[8, 9]。对土下溶蚀速率而言,无论是不同土地利用类型还是不同土壤深度之间的对比,土下溶蚀速率都与土壤CO2浓度变化趋势一致[10, 11]。如在地质、气候相似的条件下,与基岩裸露流域相比,土壤和植被覆盖较好流域由于有更多的土壤CO2参与到岩溶作用中,其土下溶蚀速率加快,岩溶碳汇效应也更强[12, 13],这说明土壤CO2与岩溶碳循环之间有着密切联系。因此本文通过对土壤CO2与岩溶碳循环的影响因素进行综述分析,为准确评估岩溶碳汇效应提供科学依据。
1 温度与降水 1.1 土壤温湿度调控土壤CO2浓度土壤温度与土壤CO2浓度有着密切的关系。土壤温度一方面控制植物的新陈代谢活动,温度升高可能导致植物呼吸作用加强,根部向土壤中释放的CO2增加。另一方面,温度升高增强了微生物活性,微生物对土壤中有机质及凋落物的分解速率加强,从而增强土壤呼吸。由于土壤CO2通量与土壤CO2浓度之间具有显著的正相关关系[14],说明土壤呼吸能够反映土壤CO2浓度的变化。许多研究已表明土壤呼吸与温度具有显著的相关关系[15, 16, 17, 18, 19]。然而土壤呼吸对温度的敏感性(通常用Q10值表示)会随温度的增加而降低。如在中国北方,Wang等[20]对河北省7个生态系统进行研究,认为土壤呼吸速率会随温度升高而升高,在年尺度上,土温单独对土壤呼吸的贡献率为86%—94%,生长季节贡献率为52%—90%;中国亚热带地区不同土地利用类型下,土壤温度仍是控制土壤呼吸的重要因素,可解释26%—59%的土壤CO2通量变化[21];而在坦桑尼亚干旱的热带农田中,土壤CO2通量则主要受到土壤湿度影响,与温度没有相关性[22]。这说明在不同的气候条件下,温度对土壤CO2产生的贡献会存在差异,并且其贡献率可能会随温度的增加而减小。
土壤湿度对土壤CO2产生的正或负作用都有记载。土壤湿度增加会刺激微生物活性,从而促进了土壤CO2的产生[23, 24, 25]。Jassal等[26]发现降水使土壤湿度突然增高,刺激了土壤中微生物活性,从而导致土壤CO2浓度明显升高。并且在干燥的土壤中,湿度成为限制土壤微生物活性的主要因子时,土壤湿度与土壤呼吸之间的相关性更为明显[27]。Sugihara等[22]也发现,土壤CO2通量与土壤湿度具有明显相关性,在雨季土壤CO2通量通常很高,而在旱季则较低。但土壤湿度并不总是会促进土壤CO2的产生,许多研究都发现土壤湿度对土壤呼吸的影响存在阀值。当浅层土壤湿度>0.6时,CO2通量会因水分阻碍气体传输而迅速下降[28];深层土壤同样也存在上下阀值(0.2—0.8)。Schindlbacher等[29]则认为土壤体积含水量为25%—45%时最有利于微生物活动。尽管不同的研究中所提出的限制土壤呼吸的阀值并不完全一致,但都认为在到达阀值之前土壤呼吸会随土壤湿度增加而增加,而到达阀值后,土壤呼吸则随土壤湿度的增加而减弱。
在岩溶区,土壤温湿度会通过土壤CO2而间接影响岩溶作用强度,即土壤CO2浓度升高,岩溶作用也随之增强,反之亦反[30]。然而与土壤CO2一样,岩溶作用通常受到土壤温度和湿度的共同控制,当其中任何一种因子成为限制条件时都会制约岩溶作用强度。较低的温度会限制土壤活性,土壤水在下渗过程中由于未能携带充分的土壤CO2补给,从而限制了岩溶作用的进行[30]。同样,在土壤湿度较低时,即使温度保持较高水平,微生物活性也会减弱,导致土壤呼吸排放的CO2速率处于较低水平,进而限制了其参与岩溶作用的程度;当土壤湿度到达中间最佳范围时,温度敏感性增加,土壤呼吸随土壤温度的升高而增强;而随着土壤湿度增加并成为土壤CO2产生的限制条件时,温度敏感性又会下降[31, 32, 33, 34, 35]。
1.2 水动力条件对土壤CO2运移的影响降水不仅可以通过土壤湿度改变土壤CO2浓度,其水动力条件更能直接导致土下溶蚀速率发生变化。在西南亚热带气候背景下,降雨对土下溶蚀速率的贡献甚至要比气温更为重要,尤其是在旱季,土壤持水能力与土壤的供水强度是决定溶蚀能力的关键因素[36]。因为在岩溶区,土壤中一部分CO2会通过地-气界面并以土壤呼吸的方式返回大气中,还有一部分会溶解到下渗水中,形成碳酸并与碳酸盐岩发生反应,最后以DIC的形式渗入地下水中[37, 38, 39]。Yang等[40]认为水动力条件与土壤CO2是主导表层岩溶带岩溶作用的两个主导因素,并且二者对岩溶作用的影响会因降水的强弱而发生改变:在强降雨条件下,由于降水造成大量土壤CO2溶于土壤水和地下水,土壤CO2浓度迅速降低,此时稀释效应对岩溶作用的影响要大于CO2效应;在降水较弱的条件下,土壤湿度引起的土壤CO2浓度变化则又成为了控制岩溶作用的主导因素。
尽管雨后岩溶泉水或地下水的水化学性质可能会受到降雨稀释效应的影响,但在水流量增加的情况下,碳通量也会随之增加[41],这说明降水溶解的CO2量增加,从而导致岩溶作用增强。另外考虑到地形因素的影响,Yan等[42]认为如果忽略水动力条件对岩溶作用的影响,在评估溶蚀速率和强度时可能会造成偏差。综上所述,降水不仅可以通过土壤湿度影响土壤CO2的产生,其带来的水动力条件变化也会通过土壤中CO2的运移而成为岩溶作用的重要影响因素。
2 土壤理化性质 2.1 土壤pH土壤pH可通过土壤中微生物的数量和种类影响土壤有机质分解以及土下溶蚀速率。土壤微生物分解凋落物和土壤有机质产生的CO2是土壤CO2的重要组成部分,而且微生物可以通过化学降解、生物物理作用以及酶作用直接对碳酸盐岩进行风化[43]。如真菌由于菌丝的穿插包裹带来的机械作用使其对碳酸盐岩的风化能力明显高于细菌[44, 45, 46, 47];碳酸酐酶高效快速催化CO2和HCO-3之间相互转化反应,可使石灰岩的溶蚀速率提高10倍[48]。因此微生物在岩溶动力系统中扮演重要角色。然而土壤中细菌、真菌、放线菌都有各自的生存环境,其对土壤pH的要求也不相同。细菌生存适宜的pH为中性和微碱性,放线菌在酸性强的土壤中生长不良,很多真菌对pH不敏感,在酸性土壤中也能良好生长[49]。如针叶林地中由于针叶树种凋落物灰分少,含树脂、单宁等酸性物质,引起土壤酸性增加,其细菌数量要少于阔叶林。但真菌数量并没有受到土壤pH值影响,针叶林地由于针叶林凋落物难分解成分多,使得能分解纤维素半纤维素、木质素等难分解物质的真菌数量大于阔叶林[50]。
此外,土壤pH对岩溶碳汇的影响还体现在土壤pH决定着下渗水中DIC的赋存形式。H2CO3、HCO-3和CO2是DIC的3种形态,它们在DIC中所占的比例则与pH有关。通常岩溶区碳酸盐岩的溶蚀会使地下水呈中性或偏碱性,从而导致地下水中的DIC以HCO-3的形式存在[51]。但是由于土壤中植物残体产生的有机质酸化土壤或者农田中施用氮肥等活动导致土壤酸化,土壤中游离的H+会与HCO-3结合生成H2CO3。由于H2CO3的不稳定性并且土壤水中pCO2通常要高于土壤或大气,水中溶解的CO2会以气体的形式从水中释放到土壤或空气中(公式1)[52]:
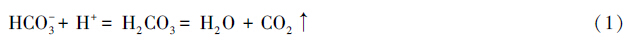
因此在具有较高pH的土壤中,参与岩溶作用中的H+主要是由CO2溶于水后形成的H2CO3提供的,碳酸盐岩的溶蚀可认为是大气或土壤CO2的汇;而在较低pH的土壤中,HNO3和H2SO4等强酸或植物分泌的有机酸向下渗水中提供H+,导致水体中DIC含量的减少并释放CO2,此时碳酸盐岩溶蚀则成为大气或土壤CO2的源[53]。
2.2 土壤颗粒与孔隙度相对于土壤中的砂粒,粘粒往往对土壤有机质的物理保护作用更强,其原因可能是由于粘粒具有较大的表面积和吸引力,其吸附土壤有机质的能力也较强[54]。在非岩溶区往往表现出砂土具有较高的CO2通量,而粘土的CO2通量则相对较低。Sotta等[55]发现砂土CO2通量比粘土高21%,并且土壤质地对土壤呼吸的贡献甚至要大于土壤湿度。Iqbal等[21]在对比不同土地利用类型下的土壤CO2通量时,也指出砂土的CO2通量要高于粘土。但在岩溶区,从农田到次生林,由于农田持续受到侵蚀和退化,粘粒成分在减少[56],土下溶蚀速率却往往表现相反的趋势。如蓝家程等[57]对重庆南川区3个不同土地利用类型下的土下溶蚀速率进行对比,发现林地下的全年溶蚀速率要高于草地和耕地。这说明在缺少土壤粘粒提供物理保护的情况下,尽管土壤有机质的矿化速率加快,但土壤孔隙度增大可能导致土壤CO2以土壤呼吸的形式返回到大气中,而并没有参与到岩溶作用中。
土壤孔隙度一方面通过下渗水影响土壤的蓄水和供水能力,如表层孔隙度大有利于水分下渗,而下层孔隙度小则有利于土壤水分的保存。深部土壤有机质以低质量的底物为主,使其需要更多的能量进行分解,并且与有机质连结在一起,使得它们难以被微生物利用[28]。但是降水下渗过程中,把表层活性碳输入到深部则会促进深部土壤有机质的分解[58]。另外表层丰富的营养物质也有可能通过土壤水下渗到深部,与水分一起为微生物生命活动提供能源,从而促进了下层土壤有机碳的矿化。另一方面土壤孔隙度也制约着土壤CO2的运移方向[59]。Jassal等[26]计算出超过75%的土壤CO2通量来自于顶部20cm处,而50cm以下的土壤有机质对土壤CO2通量的贡献很小,并且降雨后表层土壤CO2梯度增加了50%,但向大气中的通量却下降了约40%,这说明土壤CO2受土壤透气性的制约,其运移方向也有所不同。而土壤CO2的运移方向同样对岩溶作用至关重要,由于下部土壤更加紧实,气孔减小并且气体交换削弱[60],土壤中的CO2可能更容易溶于土壤水并参与到岩溶作用中,所以岩溶区土壤CO2浓度会随深度表现出双向梯度,即土壤CO2浓度最高处出现在土壤中部而不是底部[31]。
3 土地利用类型对土壤CO2的影响上覆植被的演变会明显改变土壤中碳平衡,并且会带来凋落物输入量、根部的分布及微环境的变化,这些又是影响凋落物分解、微生物活性的重要因子。
土地利用类型的转变首先会使NPP发生较大的变化。Archer等[61]和Hibbard[62]等研究表明,草原净初级生产量(NPP)为87—340g cm-2 a-1,而被木本植物侵入后,NPP增加到510—600 g cm-2 a-1。NPP增加会使植物储存的碳增多,而通过凋落物进入到土壤中的碳也会随之增加。Sugihara等[17]发现加入植物残渣的地块土壤的碳储存量和土壤CO2的通量都明显增加。Creamer等[63]在对比木本植物与草本植物下的土壤呼吸与土壤有机碳时发现,不但木本植物下土壤呼吸高于草地,而且由于其向土壤中输入的有机碳仍大于土壤呼吸消耗的有机碳,木本植物下土壤有机碳的积累速率同样大于草本植物,这也是林地土下溶蚀速率要大于草地土下溶蚀速率的一个重要原因。土壤CO2与岩溶碳循环不仅受上覆植被控制,不同土地利用类型下土壤性质的差异也是其重要影响因素。土壤为CO2提供了储存空间,使其能够溶于下渗水并与底部碳酸盐岩发生反应。反之,在缺少土壤覆盖的情况下,植物根部释放的CO2更容易返回到大气中。因此在相同植被覆盖条件下,与土壤较厚区域相比,其土下岩溶作用也相对较弱。然而,岩溶生态系统土壤的形成、演化与植被的生长相互促进[64]。土壤为植被提供了良好的生存环境,而植被带来的生态效益则有利于土壤保持[65],二者密切联系并共同影响土壤CO2与岩溶碳循环过程。
凋落物作为土壤有机质的主要来源,凋落物质量与凋落物分解速率之间有密切关系。李志安等[66]在总结前人的研究时,提出能反映凋落物质量的指标有各种含碳化合物量(如木质素、纤维素、单宁等)与养分元素含量(N、P、K等)的比,也可以直接以养分含量表示。Ge等[67]在总结森林凋落物分解的影响因素时,指出N、P、Mn、Ca、木质素等对凋落物分解的影响可能会因分解阶段以及研究地区的不同而存在差异。例如在凋落物的早期分解阶段,充足的N会刺激凋落物早期分解,而在后期,大量的N则可能会抑制凋落物的分解[68],其原因可能与微生物种类及凋落物质量有关。真菌对木质素的降解往往会受到高N含量的限制,所以在凋落物的早期分解过程中,水溶性有机碳等易分解物质占主导,N含量往往对凋落物的分解起促进作用,而在后期,分解物质主要为木质素等难分解物质,N含量则又会对凋落物分解和土壤CO2排放起到阻碍作用[69]。
植物根系也是影响土壤CO2的一个重要因素。植物根部不仅仅直接通过呼吸作用向土壤中释放CO2,而且根部产生的分泌物使根际环境非常有利于微生物生存。另外植物根部分泌物还可以对土壤有机碳的矿化起到促进或抑制作用。许多证据表明提高植物活性和地下碳输入会直接或间接的导致土壤有机质分解加强[70],但由于植物与根际微生物之间的竞争或土壤微生物首先利用来自根系的有机物,导致其对土壤有机碳的分解起到抑制作用[71]。Kuzyakov[72]则认为根际环境产生的激发效应与植物分泌物的成分有密切关系:在土壤缺少易分解C和N的情况下,根与土壤微生物竞争会阻碍土壤有机质分解;在C含量及C利用率较好的情况下,根际低C/N的输入会促使微生物分解土壤有机质以获得额外的N;在高N低C的情况下,易被利用的根际分泌物的输入使微生物分解从土壤有机质转向根际分泌物;如果二者都不是限制因素,根际环境的激发效应也不会存在。
4 HNO3与H2SO4对岩溶碳循环的影响随着人类活动的干预(如氮肥的使用、大气酸沉降以及工业废渣的堆放),HNO3与H2SO4也参与到了岩溶碳循环中。从地质时间尺度上讲,HNO3与H2SO4溶蚀碳酸盐岩不但不会消耗CO2,其产生的HCO-3会随碳酸盐矿物沉淀而向大气释放CO2(公式2,公式3),从而减少了岩溶作用对大气或土壤CO2的净消耗量,并导致岩溶碳汇效应减弱:
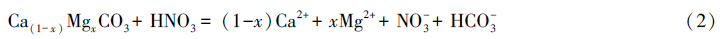

氮肥硝化作用产生的HNO3溶蚀碳酸盐岩释放的CO2占硅酸盐风化碳汇的2%—5%(包括页岩)或6%—15%(不包括页岩)[73]。我国大陆H2SO4风化碳酸盐矿物则向大气释放的CO2达28×1012g/a,这相当于全球硅酸盐风化消耗CO2量的26%[74]。因此在研究岩溶碳循环时,如不考虑HNO3与H2SO4的影响,往往会造成对岩溶碳汇的过高估算。
水化学成分分析法以及碳同位素的应用有利于准确评估岩溶碳汇作用。在没有HNO3或H2SO4等其它酸的干扰下,CO2溶于水后形成的H2CO3是溶蚀碳酸盐岩唯一的酸。溶蚀后产生的HCO-3一半来自大气或土壤中的CO2,另一半来自碳酸盐岩,Ca+Mg与HCO-3的摩尔比为0.5。然而在HNO3或H2SO4参与的情况下,其溶蚀作用产生的HCO-3全部来自于碳酸盐岩,因此Ca+Mg与HCO-3的摩尔比要>0.5。运用这一原理,对河流水化学成分进行测量就可分析HNO3或H2SO4对产生HCO-3的贡献量以及岩溶作用消耗的CO2量。如Etchanchu等[75]和Semhi等[76]通过对水化学成分进行分析,估算了氮肥产生的HNO3对碳酸盐岩溶解的贡献为30%和12%—26%。碳同位素分析也是研究HNO3或H2SO4参与岩溶作用的一种重要方法。农田和住宅区由于HNO3与H2SO4参与到岩溶作用中,其δ13CDIC明显偏重于林地和草地[77]。李军等[78]对长江及其主要支流河水水化学和δ13CDIC值进行研究发现,H2SO4溶解碳酸盐岩使DIC含量与δ13CDIC值都升高,并且二者之间呈现明显的正相关关系,即H2SO4风化碳酸盐岩对河流DIC的贡献每增加10%就将导致δ13CDIC值升高0.85‰。
5 结论土壤CO2作为岩溶作用的主要驱动条件,受到气候条件、土壤性质以及植被类型等因素控制,进而影响到岩溶碳循环。在植被覆盖条件较好的土地利用类型下,植被调节土壤温湿度;植物残体以及根部分泌物向土壤中输入有机物为土壤微生物提供了物质和能量来源;以及植被促进土壤的形成与演化。这些因素都将促进土壤CO2的产生以及岩溶碳循环过程。因此加强对石漠化地区的治理,不仅会提高生物和土壤的固碳能力,也会增强岩溶碳汇效应。
各个影响因子之间相互联系并相互制约,在不同的条件下,其对土壤CO2及岩溶碳循环的影响也会出现差异。另外,HNO3与H2SO4溶蚀碳酸盐岩并释放CO2,导致岩溶作用对CO2的净消耗量减少,从而减弱岩溶碳汇效应。加强环境保护将会减少HNO3与H2SO4的参与,进而间接对岩溶碳循环起到增汇减源的作用。
| [1] | Larson C. An unsung carbon sink. Science, 2011, 334(6058): 886-887. |
| [2] | Curl R L. Carbon shifted but not sequestered. Science, 2012, 335(6069): 655. |
| [3] | Yuan D X. The carbon cycle in karst. Zeitschrift für Geomorphologie Neue Folge, 1997,108: 91-102. |
| [4] | Yuan D X. Sensitivity of karst process to environmental change along the PEP II transect. Quaternary International, 1997, 37: 105-113. |
| [5] | Cao J H, Yuan D X, Groves C, Fen H, Hui Y, Qian L. Carbon fluxes and sinks: the consumption of atmospheric and soil CO2 by carbonate rock dissolution. Acta Geologica Sinica-English Edition, 2012, 86(4): 963-972. |
| [6] | 刘再华, 李强, 孙海龙, 汪进良, 吴孔运. 云南白水台钙华水池中水化学日变化及其生物控制的发现. 水文地质工程地质, 2005, 32(6): 10-15. |
| [7] | Liu Z H, Dreybrodt W, Wang H J. A new direction in effective accounting for the atmospheric CO2 budget: Considering the combined action of carbonate dissolution, the global water cycle and photosynthetic uptake of DIC by aquatic organisms. Earth-Science Reviews, 2010, 99(3): 162-172. |
| [8] | White W B. Carbon fluxes in Karst aquifers: Sources, sinks, and the effect of storm flow. Acta Carsologica, 2013, 42(2/3):177-186. |
| [9] | Martin J B, Brown A, Ezell J. Do carbonate karst terrains affect the global carbon cycle. Acta Carsologica, 2013, 42(2/3):187-196. |
| [10] | 章程. 不同土地利用下的岩溶作用强度及其碳汇效应. 科学通报, 2011, 56(26): 2174-2180. |
| [11] | Xu S, He S. The CO2 regime in soil profile and its drive to dissolution in carbonate rock area. Carsological Sinica, 1996, 15(1/2): 50-57. |
| [12] | 赵敏, 曾成, 刘再华. 土地利用变化对岩溶地下水溶解无机碳及其稳定同位素组成的影响. 地球化学, 2009, 38(6): 565-572. |
| [13] | Zhao M, Zeng C, Liu Z H, Wang S J. Effect of different land use/land cover on karst hydrogeochemistry: a paired catchment study of Chenqi and Dengzhanhe, Puding, Guizhou, SW China. Journal of Hydrology, 2010, 388(1): 121-130. |
| [14] | 程建中, 李心清, 周志红, 王兵, 程红光, 邢英, 唐源. 土壤CO2浓度与地表CO2通量的季节变化及其相互关系. 地球与环境, 2011, 39(2): 196-202. |
| [15] | Rochette P, Desjardins R L, Pattey E. Spatial and temporal variability of soil respiration in agricultural fields. Canadian Journal of Soil Science, 1991, 71(2): 189-196. |
| [16] | Lloyd J, Taylor J A. On the temperature dependence of soil respiration. Functional Ecology, 1994, 8(3): 315-323. |
| [17] | Liu X Z, Wan S Q, Su B, Hui D F, Luo Y Q. Response of soil CO2 efflux to water manipulation in a tallgrass prairie ecosystem. Plant and Soil, 2002, 240(2): 213-223. |
| [18] | Fang C, Moncrieff J B. The dependence of soil CO2 efflux on temperature. Soil Biology and Biochemistry, 2001, 33(2): 155-165. |
| [19] | Mielnick P C, Dugas W A. Soil CO2 flux in a tallgrass prairie. Soil Biology and Biochemistry, 2000, 32(2): 221-228. |
| [20] | Wang W, Peng S S, Wang T, Fang J Y. Winter soil CO2 efflux and its contribution to annual soil respiration in different ecosystems of a forest-steppe ecotone, north China. Soil Biology and Biochemistry, 2010, 42(3): 451-458. |
| [21] | Iqbal J, Ronggui H, Lijun D, Lan L, Shan L I N, Tao C, Leilei R. Differences in soil CO2 flux between different land use types in mid-subtropical China. Soil Biology and Biochemistry, 2008, 40(9): 2324-2333. |
| [22] | Sugihara S, Funakawa S, Kilasara M, Kosaki T. Effects of land management on CO2 flux and soil C stock in two Tanzanian croplands with contrasting soil texture. Soil Biology and Biochemistry, 2012, 46: 1-9. |
| [23] | Luo Y, Zhou X H. Soil Respiration and the Environment. London: Academic Press, 2010: 328. |
| [24] | Manzoni S, Schimel J P, Porporato A. Responses of soil microbial communities to water stress: results from a meta-analysis. Ecology, 2012, 93(4): 930-938. |
| [25] | Nagy Z, Pintér K, Czóbel S, Balogh J, Horváth L, Fóti S, Barcza Z, Weidinger T, Csintalan Z, Dinh N Q, Grosz B, Tuba Z. The carbon budget of semi-arid grassland in a wet and a dry year in Hungary. Agriculture, Ecosystems & Environment, 2007, 121(1): 21-29. |
| [26] | Jassal R, Black A, Novak M, Morgenstern K, Nesic Z, Gaumont-Guay D. Relationship between soil CO2 concentrations and forest-floor CO2 effluxes. Agricultural and Forest Meteorology, 2005, 130(3): 176-192. |
| [27] | Fóti S, Balogh J, Nagy Z, Herbst M, Pintér K, Péli E, Koncz P, Bartha S. Soil moisture induced changes on fine-scale spatial pattern of soil respiration in a semi-arid sandy grassland. Geoderma, 2014, 213: 245-254. |
| [28] | Gabriel C E, Kellman L. Investigating the role of moisture as an environmental constraint in the decomposition of shallow and deep mineral soil organic matter of a temperate coniferous soil. Soil Biology and Biochemistry, 2014, 68: 373-384. |
| [29] | Schindlbacher A, Zechmeister-Boltenstern S, Kitzler B, Jandl R. Experimental forest soil warming: response of autotrophic and heterotrophic soil respiration to a short-term 10℃ temperature rise. Plant and Soil, 2008, 303(1/2): 323-330. |
| [30] | 章程, 蒋忠诚, 何师意, 蒋勇军, 李林立, 王建力. 垂直气候带岩溶动力系统特征研究——以重庆金佛山国家级自然保护区为例. 地球学报, 2006, 27(5): 510-514. |
| [31] | 曹建华, 周莉, 杨慧, 卢茜, 康志强. 桂林毛村岩溶区与碎屑岩区林下土壤碳迁移对比及岩溶碳汇效应研究. 第四纪研究, 2011, 31(3): 431-437. |
| [32] | Curiel Yuste J, Baldocchi D D, Gershenson A, Goldstein A, Misson L, Wong S. Microbial soil respiration and its dependency on carbon inputs, soil temperature and moisture. Global Change Biology, 2007, 13(9): 2018-2035. |
| [33] | Jassal R S, Black T A, Novak M D, Gaumont-Guay D, Nesic Z. Effect of soil water stress on soil respiration and its temperature sensitivity in an 18-year-old temperate Douglas-fir stand. Global Change Biology, 2008, 14(6): 1305-1318. |
| [34] | Suseela V, Conant R T, Wallenstein M D, Dukes J S. Effects of soil moisture on the temperature sensitivity of heterotrophic respiration vary seasonally in an old-field climate change experiment. Global Change Biology, 2012, 18(1): 336-348. |
| [35] | Craine J M, Gelderman T M. Soil moisture controls on temperature sensitivity of soil organic carbon decomposition for a mesic grassland. Soil Biology and Biochemistry, 2011, 43(2): 455-457. |
| [36] | 章程. 不同土地利用土下溶蚀速率季节差异及其影响因素. 地质论评, 2010, 56(1): 136-140. |
| [37] | Brook G A, Folkoff M E, Box E O. A world model of soil carbon dioxide. Earth Surface Processes and Landforms, 1983, 8(1): 79-88. |
| [38] | Jassal R S, Black T A, Drewitt G B, Novak M D, Gaumont-Guay D, Nesic Z. A model of the production and transport of CO2 in soil: predicting soil CO2 concentrations and CO2 efflux from a forest floor. Agricultural and Forest Meteorology, 2004, 124(3): 219-236. |
| [39] | Liu Z H, Li Q, Sun H L, Wang J L. Seasonal, diurnal and storm-scale hydrochemical variations of typical epikarst springs in subtropical karst areas of SW China: Soil CO2 and dilution effects. Journal of Hydrology, 2007, 337(1): 207-223. |
| [40] | Yang R, Liu Z H, Zeng C, Zhao M. Response of epikarst hydrochemical changes to soil CO2 and weather conditions at Chenqi, Puding, SW China. Journal of Hydrology, 2012, 468: 151-158. |
| [41] | Jiang Z C, Lian Y Q, Qin X Q. Carbon cycle in the epikarst systems and its ecological effects in South China. Environmental Earth Sciences, 2013, 68(1): 151-158. |
| [42] | Yan J H, Li J M, Ye Q, Li K. Concentrations and exports of solutes from surface runoff in Houzhai Karst Basin, southwest China. Chemical Geology, 2012, 304-305: 1-9. |
| [43] | 连宾, 袁道先, 刘再华. 岩溶生态系统中微生物对岩溶作用影响的认识. 科学通报, 2011, 56(26): 2158-2161. |
| [44] | Lian B, Chen Y, Zhu L J, Yang R D. Effect of microbial weathering on carbonate rocks. Earth Science Frontiers, 2008, 15(6): 90-99. |
| [45] | Jongmans A G, Van-Breemen N, Lundström U, Van-Hees P A W, Finlay R D, Srinivasan M, Unestam T, Giesler R, Melkerud P A, Olsson M. Rock-eating fungi. Nature, 1997, 389(6652): 682-683. |
| [46] | Sterflinger K. Fungi as geologic agents. Geomicrobiology Journal, 2000, 17(2): 97-124. |
| [47] | Gorbushina A A. Life on the rocks. Environmental Microbiology, 2007, 9(7): 1613-1631. |
| [48] | 刘再华. 碳酸酐酶对碳酸盐岩溶解的催化作用及其在大气CO2沉降中的意义. 地球学报, 2001, 22(5): 477-480. |
| [49] | 亚历山大. 土壤微生物学导论. 北京: 科学出版社, 1983: 34. |
| [50] | 薛立, 邝立刚, 陈红跃, 谭绍满. 不同林分土壤养分、微生物与酶活性的研究. 土壤学报, 2003, 40(2): 280-285. |
| [51] | Wallin M, Buffam I, quist M, Laudon H, Bishop K. Temporal and spatial variability of dissolved inorganic carbon in a boreal stream network: Concentrations and downstream fluxes. Journal of Geophysical Research: Biogeosciences (2005-2012), 2010, 115(G2), doi: 10.1029/2009JG001100. |
| [52] | West T O, McBride A C. The contribution of agricultural lime to carbon dioxide emissions in the United States: dissolution, transport, and net emissions. Agriculture, Ecosystems & Environment, 2005, 108(2): 145-154. |
| [53] | Hamilton S K, Kurzman A L, Robertson G P. Liming of agricultural soils: a source or sink for CO2[2002-11-19]. http://chamaecyparis.sgcp.ncsu.edu/carbon2002/2002pres/Hamilton_Stephe- n.pdf, html. |
| [54] | Sposito G, Skipper N T, Sutton R, Park S H, Soper A K, Greathouse J A. Surface geochemistry of the clay minerals. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(7): 3358-3364. |
| [55] | Sotta E D, Veldkamp E, Guimaraes B R, Paixao R K, Ruivo M L P, Almeida S S. Landscape and climatic controls on spatial and temporal variation in soil CO2 efflux in an Eastern Amazonian Rainforest, Caxiuanã, Brazil. Forest Ecology and Management, 2006, 237(1): 57-64. |
| [56] | 张治伟, 朱章雄, 文志林. 岩溶山地土壤微团聚体组成及其与土壤性质的关系. 水土保持学报, 2014, 28(1): 123-128. |
| [57] | 蓝家程, 傅瓦利, 彭景涛, 周小萍, 肖时珍, 袁波. 不同土地利用方式土下岩溶溶蚀速率及影响因素. 生态学报, 2013, 33(10): 3205-3212. |
| [58] | Bissonnais Y. Aggregate stability and assessment of soil crustability and erodibility: Ⅰ. Theory and methodology. European Journal of Soil Science, 1996, 47(4): 425-437. |
| [59] | 刘文, 张强, 贾亚男. 气象要素及土壤理化性质对不同土地利用方式下冬夏岩溶作用的影响. 生态学报, 2014, 34(6): 1418-1428. |
| [60] | Weisskopf P, Reiser R, Rek J, Oberholzer H R. Effect of different compaction impacts and varying subsequent management practices on soil structure, air regime and microbiological parameters. Soil and Tillage Research, 2010, 111(1): 65-74. |
| [61] | Archer S, Boutton T W, Hibbard K A. Trees in grasslands: biogeochemical consequences of woody plant expansion. Global Biogeochemical Cycles in the Climate System, 2001: 115-138. |
| [62] | Hibbard K A, Schimel D S, Archer S, Ojima D S, Parton W. Grassland to woodland transitions: integrating changes in landscape structure and biogeochemistry. Ecological Applications, 2003, 13(4): 911-926. |
| [63] | Creamer C A, Filley T R, Boutton T W, Oleynik S, Kantola I B. Controls on soil carbon accumulation during woody plant encroachment: Evidence from physical fractionation, soil respiration, and δ13C of respired CO2. Soil Biology and Biochemistry, 2011, 43(8): 1678-1687. |
| [64] | 李阳兵, 王世杰, 李瑞玲. 岩溶生态系统的土壤. 生态环境, 2004, 13(3): 434-438. |
| [65] | 李阳兵, 王世杰, 周德全. 茂兰岩溶森林的生态服务研究. 地球与环境, 2005, 33(2): 39-44. |
| [66] | 李志安, 邹碧, 丁永祯, 曹裕松. 森林凋落物分解重要影响因子及其研究进展. 生态学杂志, 2004, 23(6): 77-83. |
| [67] | Ge X G, Zeng L X, Xiao W F, Huang Z L, Geng X S, Tan B W. Effect of litter substrate quality and soil nutrients on forest litter decomposition: A review. Acta Ecologica Sinica, 2013, 33(2): 102-108. |
| [68] | Berg B, Johansson M B, Ekbohm G, McClaugherty C, Rutigliano F, Santo A V D. Maximum decomposition limits of forest litter types: a synthesis. Canadian Journal of Botany, 1996, 74(5): 659-672. |
| [69] | Wilson H M, Al-Kaisi M M. Crop rotation and nitrogen fertilization effect on soil CO2 emissions in central Iowa. Applied Soil Ecology, 2008, 39(3): 264-270. |
| [70] | Taneva L, Gonzalez-Meler M A. Decomposition kinetics of soil carbon of different age from a forest exposed to 8 years of elevated atmospheric CO2 concentration. Soil Biology and Biochemistry, 2008, 40(10): 2670-2677. |
| [71] | Kuzyakov Y, Cheng W. Photosynthesis controls of CO2 efflux from maize rhizosphere. Plant and Soil, 2004, 263(1): 85-99. |
| [72] | Kuzyakov Y. Review: factors affecting rhizosphere priming effects. Journal of Plant Nutrition and Soil Science, 2002, 165(4): 382-396. |
| [73] | Perrin A S, Probst A, Probst J L. Impact of nitrogenous fertilizers on carbonate dissolution in small agricultural catchments: Implications for weathering CO2 uptake at regional and global scales. Geochimica et Cosmochimica Acta, 2008, 72(13): 3105-3123. |
| [74] | 刘丛强, 蒋颖魁, 陶发祥, 郎赟超, 李思亮. 西南喀斯特流域碳酸盐岩的硫酸侵蚀与碳循环. 地球化学, 2008, 37(4): 404-414. |
| [75] | Etchanchu D, Probst J L. Evolution of the chemical composition of the Garonne River water during the period 1971-1984. Hydrological Sciences Journal, 1988, 33(3): 243-256. |
| [76] | Semhi K, Amiotte Suchet P, Clauer N, Probst J L. Impact of nitrogen fertilizers on the natural weathering-erosion processes and fluvial transport in the Garonne basin. Applied Geochemistry, 2000, 15(6): 865-878. |
| [77] | Jiang Y J. The contribution of human activities to dissolved inorganic carbon fluxes in a karst underground river system: Evidence from major elements and δ13CDIC in Nandong, Southwest China. Journal of Contaminant Hydrology, 2013, 152: 1-11. |
| [78] | 李军, 刘丛强, 李龙波, 李思亮, 王宝利, Chetelat B. 硫酸侵蚀碳酸盐岩对长江河水DIC循环的影响. 地球化学, 2010, 39(4): 305-313. |