 2015, Vol. 35
2015, Vol. 35文章信息
- 王兴昌, 王传宽
- WANG Xingchang, WANG Chuankuan
- 森林生态系统碳循环的基本概念和野外测定方法评述
- Fundamental concepts and field measurement methods of carbon cycling in forest ecosystems: a review
- 生态学报, 2015, 35(13): 4241-4256
- Acta Ecologica Sinica, 2015, 35(13): 4241-4256
- http://dx.doi.org/10.5846/stxb201407011359
-
文章历史
- 收稿日期:2014-07-01
- 网络出版日期:2015-01-27






森林在全球碳循环中扮演重要角色,在调节大气CO2浓度和维持生命系统等方面具有不可替代的作用。森林土壤碳库(742 Pg C)占陆地土壤碳库(1502 Pg C)[1]的49%,而森林植被碳库(363 Pg C)[2]占陆地植被碳库(497 Pg C)[3]的73%。全球陆地生态系统的总初级生产力大约一半由森林完成[4]。全球陆地生态系统固定(2.6 ± 0.8) Pg C/a[5],其中森林(未包括土地利用变化)固定(2.4 ± 0.4) Pg C/a[2],甚至大于海洋碳汇(2.2 Pg C/a)[6]。
由于全球陆地生态系统复杂多样,其碳汇很难直接定量评估,因此绝大多数研究将其作为全球大气碳平衡方程的残余项来处理[5]。森林是陆地生态系统各类型中最复杂的,其丰富的树种组成、高大的冠层和巨大的空间变异等特点决定了全球森林碳汇的直接评估面临更大的困难。例如,一项分析全球森林生态系统尺度CO2平衡数据集的整合研究发现,各森林生物群区的碳收支大约需要10%—60%的总初级生产力才能闭合[7]。
国内已有文献总结和评论过陆地或森林生态系统碳循环的概念[8]和测量方法[9, 10],但这些文献各有侧重,尚缺乏系统阐述。本文目的是在这些文献的基础上,系统评述国内外的最新进展,进一步澄清森林生态系统碳循环的相关概念,在此基础上重点讨论生态系统尺度森林碳循环主要过程的野外测定方法及其误差来源,为我国森林生态系统碳循环研究提供可靠的方法论。
1 森林碳循环相关概念和术语 1.1 碳浓度“浓度”一词首先出现在化学中,指溶液中溶质的分量。在大气科学中,气体标量强度常用3类方法表示[11, 12],以CO2为例:(1)质量密度(ρc,mg/m3)和摩尔浓度(ρc,m,mmol/m3);(2)摩尔分数(cc,μmol/mol1 或μL/L);(3)干质量分数(ρc,m,mg/kg)和干摩尔分数(ρc,μmol/mol),又分别称为质量混合比和摩尔混合比。干摩尔分数消除了水汽物质的稀释作用而具有守恒性,且方便用于理想气体状态方程,是CO2通量研究中最方便的表达方式[12, 13, 14, 15]。而在生态学中,植物[16]、植物残体和土壤[17]的碳“浓度”、含碳量或含碳率,是描述固体混合物中某种物质的相对量,实质上均为碳的质量分数(Mass fraction,% dry mass或mg/g)。植物非结构性碳水化合物(NSC)的浓度也为质量分数[18, 19]。
1.2 碳储量碳储量、碳贮量或碳蓄积量是描述碳库大小的术语,与研究的空间尺度密切相关。碳密度是面积标准化的碳储量或碳贮量(Mg C/hm2或kg C/m2)[20]。森林生态系统的碳库可分为植被、植物残体和土壤3部分。植被碳库即为生物量碳库,表示所有生物体组分的质量,一般指活体植物的质量[8, 21]。与生物量相区别的是植物残体,包括枯枝落叶、倒木、枯立木、树桩和死根,或称死生物量。将生物量和植物残体量转化为碳密度要用到碳质量分数[16]。土壤碳储量既与空间尺度有关,还受到取样深度的制约[17],所以常用单位体积原状土壤所含的碳总干重即SOC密度(mg C/cm3或kg C/m3)表示。国际上常用1 m作为深度标准[22],因此单位面积SOC储量也称SOC密度(kg C/m2)[23]。
1.3 碳通量相关概念碳通量表示生态系统中单位时间通过单位地表面积的某一特定组分碳的量[24]。森林生态系统碳循环通过植物光合作用固定CO2进入生态系统,因此与生态系统的生产力概念密不可分[8]。不同时空尺度生产力有不同的内涵,主要有如下概念和术语:
(1)总初级生产力(GPP)是指单位地表面积上单位时间内绿色植物通过光合作用途径所固定的碳量[25]。GPP一般是日尺度到年尺度上生态系统的总光合作用[25],并没有扣除光合器官叶片暗呼吸作用消耗的碳。
(2)自养呼吸(Ra)是指单位地表面积单位时间内初级生产者(绿色植物)活体部分的呼吸总量(CO2-C产量)[26],又称为群落呼吸[21]。Ra包括叶片呼吸(有时林下草本呼吸单独测定)、枝干呼吸和根系呼吸。
(3)净初级生产力(NPP)表示植被的净碳吸收,也译为净第一性生产力,即NPP等于总初级生产力减去自养呼吸碳消耗[25]:

(4)异养呼吸(Rh)指单位地表面积单位时间异养生物的呼吸量,包括土壤有机质、枯枝落叶层和粗木质残体呼吸。动物呼吸因量级很小而常被忽略。值得注意的是,根际微生物和共生菌根菌的呼吸作为Rh的一部分,但所利用的碳源主要是根系分泌物,因此属于Ra还是Rh尚存争议[27]。
(5)生态系统呼吸(Re)指单位地表面积单位时间生态系统所有有机体的呼吸总量,包括自养呼吸和异养呼吸,即:

(6)净生态系统生产力(NEP)指生态系统光合作用固定的碳与呼吸作用释放的碳之差[26]:

在稳定的自然生态系统中,NEP接近生态系统净碳累积速率,但在生态系统中发生明显的非CO2形式的碳通量或非呼吸CO2流失时(如火烧和采伐),则NEP与净碳累积速率会出现较大偏差。为此,Chapin等[26]提出了净生态系统碳平衡的概念。
(7)净生态系统碳平衡(NECB)为生态系统的净碳积累速率[26],可用如下公式表示:
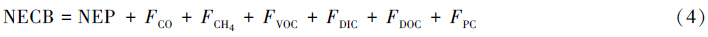
式中,FCO、FCH4、FVOC、FDIC、FDOC和FPC分别表示一氧化碳、甲烷、可挥发性有机碳、可溶性无机碳、可溶性有机碳和颗粒碳的净通量。为了得到生态系统的碳平衡,应该在公式(3)中增加气象、水文和干扰等途径产生的碳通量。在更大的时空尺度上,研究区域NECB的平均值等价于净生物群区生产力(NBP)[26]。林火和采伐干扰导致的瞬时碳通量在数值上往往很大,人们通常给予了较为充分的考虑,但绝大多数研究没有考虑可溶性无机碳(DIC)、可溶性有机碳(DOC)和颗粒碳(PC)通量[26, 28]。试验观测的净碳通量与长期碳平衡之间的差异主要是由非CO2形式的碳流失和非呼吸CO2流失造成的[7]。
(8)净生态系统交换(NEE)定义为大气—植被界面的净CO2通量[29],常用涡度协方差(EC)技术测定。NEE是气象学家的定义,负值表示生态系统从大气中吸收CO2,在符号上与NEP相反。如果忽略两种方法的误差以及无机过程CO2气体通量,二者在数值上相等[26, 30]。
(9)总生态系统交换(GEE)表示大气与生态系统的总CO2交换量。GEE在数值上等于总生态系统生产力(GEP),近似等同于GPP,但符号相反。因此有下式:
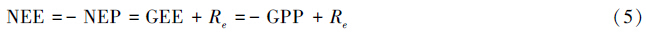
(10)其他术语:碳吸收表示为GPP,有时也指NEP;与其对应的是碳排放或释放。还有碳固定、碳固持或碳吸存,指生态系统从大气中吸收的CO2-C并封存在长期碳库中[31],包括生物和非生物过程[32],强调固碳的长效性。还有碳收支、碳平衡,二者含义有相似之处,但前者注重碳吸收和释放的过程。国内文献经常使用“净固碳量”,有时指NPP,有时指NEP,有时介于二者之间,这由研究对象是植被地上部分、整个植被还是生态系统决定。
根据以上基本概念和术语,为了更清楚地表明森林生态系统碳循环中各个过程之间的联系和区别,提出如下森林生态系统碳循环的概念模型(图 1)。

|
| 图 1 森林生态系统碳的生物地球化学循环概念模型 Fig.1 Biplot of the first two axes of the RDA (redundancy analysis) for water-salt indicators GPP: 总初级生产力gross primary productivity;NPP: 净初级生产力net primary productivity;NEP: 净生态系统生产力net ecosystem productivity;NEE: 净生态系统碳交换net ecosystem exchange;NECB: 生态系统净碳平衡net ecosystem carbon balance;NBP: 净生物群区生产力net biome production;Ra: 自养呼吸autotrophic respiration;Rh: 异养呼吸heterotrophic respiration;VOC: 植物源可挥发性有机碳biogenic volatile organic compound;DC: 可溶性碳dissolved carbon;PC: 颗粒碳particulate carbon;CO: 一氧化碳inorganic carbonaceous gas (carbon dioxide and carbon monoxide);SC: 烟尘碳soot carbon;BC: 啃食和收获的生物量碳损失biomass carbon flow to herbivores and harvesting;DIC: 可溶性无机碳dissolved inorganic carbon;DOC: 可溶性有机碳dissolved organic carbon; 箭头方向和粗细表示碳通量方向和大小 |
森林碳分配常常涉及三个方面[24]:(1)生物量分配,指各组分的当前生物量(现存量),是对碳库的静态描述;(2)碳通量,指各组分消耗的GPP绝对量;(3)碳分割,即各组分消耗的GPP分数。生物量分配指个体或植被不同组分生物量占总生物量的百分比[24],是生态系统各组分碳通量积累的结果。由于碳在树木不同器官的质量分数变化范围相对不大,生物现存量大体上体现了碳现存量的分布格局。植物个体的生物量分配情况反映了植物的异速生长关系对环境的响应,是极其重要的生理生态学参数[33]。如根冠比(RSR)指根与地上生物量的比值,它是大尺度上根据地上生物量估计地下生物量的有效参数[34]。生态系统碳分配是生态系统不同组分的碳库(如植被、土壤和植物残体库)占总碳库的百分比,与“碳分配”对应[20]。陆地生态系统模型需要碳分割的信息,而完整的碳通量测定是计算碳分割的前提。然而,生物量分配不能真实反映森林生态系统各组分的碳通量和碳分割[24]。光合产物(GPP)在生态系统中的分配格局决定了碳在生态系统中的驻留时间,对生物量和土壤碳储量至关重要,进而可对区域和全球碳循环产生深远的影响。以往绝大多数研究集中在个体尺度,生态系统尺度的研究还很薄弱,只有少量研究测定了完整的碳通量组分和分割系数[24, 35]。
1.5 碳周转生态系统的碳驻留时间指通过光合作用输入生态系统到通过呼吸作用或化学转换进入大气的时间[7],又叫做周转时间,即周转率的倒数。生态系统碳库各组分的周转时间差异很大,木质部为几年到几个世纪,而叶和细根的周转时间只有几个月到几年[7]。此外,细根周转率可以定义为细根年生产量与细根最大生物量[36]或平均生物量[37]的比值,是细根寿命的倒数。当细根生物量处于稳态时,细根周转率为细根NPP估算提供了极大的方便。SOC组成复杂,包括埋藏的植物残体、颗粒有机碳、腐殖质碳和惰性有机碳,不同组分的周转时间从月尺度变化至千年尺度[38],并随着土壤深度的增加逐渐延长[39]。
2 森林碳循环野外测定方法 2.1 碳储量测定方法 2.1.1 生物量碳库森林生物量主要有4种测定方法[40]:(1)收获法,(2)测树学法,(3)材积源法和(4)遥感反演法。由于林木高大,收获法尤其是根系挖掘难度很大,一般不常用[41];而材积源和遥感反演法一般用于大尺度生物量估算[42]。限于篇幅,本文只讨论生态系统尺度最常用的测树学方法。该方法通过破坏性取样获取标准木生物量,并与易测变量(比如胸径DBH和树高)建立生物量异速生长模型,又称为异速生长法或清单法。它的优势在于方便可靠,是森林生态系统碳循环过程应用最普遍的方法[43, 44]。尽管如此,该方法应用不当时也会产生较大误差。其误差主要包括以下4部分:(a)采样误差,即生物量的空间变异和样地的代表性问题[45, 46]。(b)生物量异速生长方程的误差。其误差包括建模与应用区域不同引起的误差[47]、模型形式和建模方式的误差[48]、超出建模DBH范围的误差。一元生物量方程比二元方程精度略低,但是树高难以测量且误差大,因此权衡精度与可操作性,一元方程通常已经足够[41, 49]。(c)变量测量误差。DBH测量一般采用围尺测定(二元模型还包括树高),但是该方法的人为因素也不容忽视[50]。(d)植物碳质量分数。假设树木碳质量分数为常数(50%)也是生物量碳估算的误差来源之一[51],例如在东北典型森林乔木层中碳密度估算误差可达±7%[52]。碳质量分数测定方法可以分为湿烧法和干烧法,后者比前者精确得多[51],是近年来的国际标准方法。
2.1.2 植物残体碳库植物残体碳库可分为粗木质残体碳库和细植物残体碳库。粗木质残体(CWD),包括枯立木、倒木、粗枝、木质碎片、树桩和粗根[53]。一般来讲以直径≥10 cm,长度≥1 m的木质残体定义为CWD,尺寸小于该标准的为细木质残体[54]。CWD最小直径定义过大将给细木质残体的测量带来较大的麻烦和误差,而定义过小将大大增加测量CWD的工作量。CWD空间分布[55]、腐烂等级和密度的高度空间异质性,导致其碳储量估算精度不高。枯立木碳库一般在样地植被调查时已经包括,但有时需要校正木材密度降低导致的误差。地面的CWD常用固定面积样方取样法、样带截面法和垂直距离取样法估计生态系统的CWD现存量[56]。直径小于起测直径的全部木质残体加上凋落叶和枯草称为细植物残体,通常采用更小的样方法调查,可以进一步分为小枝(小于CWD的最小直径)、未分解叶和半分解叶3部分。各组分现存量根据对应的碳质量分数和样方面积转化为碳密度。植物残体碳质量分数一般也采用干烧法测定。针叶树种C质量分数随CWD腐烂等级增大而升高,阔叶树种则没有变化[57]。
2.1.3 土壤有机碳库生态系统的土壤有机碳库的测量有两种方法:一种是土壤剖面法,按土壤剖面的自然发生层取样[17]或机械分层取样[1];另一种是土钻法,按机械分层取样。此外,土壤采样的同时应测定各层土壤容重。土壤剖面法可用环刀法采样,土钻法可直接根据土钻内径和高度确定的体积和土壤干重计算容重。第一种方法由于取样面积较大,土壤容重测定准确,实际估算的结果更可靠,缺点是费时费力。第二种方法较为简易,但在多石土壤中难以完成深层取样。需要注意的是,无论用哪种方法,土壤容重都应该剔除直径>2 mm石砾的质量[17, 58],因为土壤SOC测定的土壤样品通常已经剔除了石砾影响。各层土壤的SOC质量分数乘以各自容重得到各层SOC密度,各层求和就是剖面的SOC密度或储量[17]。SOC质量分数一般采用干烧法测定,越来越多的研究表明以往认为的土壤有机质含58%碳元素与实际不符,缺省时用理论值50%更合理[59]。
2.2 森林碳通量主要测定方法森林生态系统碳通量的测定方法主要包括测树学法或清单法[44]、箱法[60, 61, 62, 63, 64]、涡度协方差法(EC)[11, 29]、遥感法[65]、大气反演法[66]和生态模型法[67, 68]。按照研究的空间尺度,测树学方法和箱法属于“自下而上”的方法,而遥感法、大气反演和生态模型属于“自上而下”的方法[10]。EC法起到了将“自下而上”法与“自上而下”法联系起来的桥梁作用。测树学法、箱法和EC法是碳通量的实测方法,其余方法是大尺度碳通量的估计方法,必须以前面的实测结果为基础才能应用[9]。3种实测方法适宜的时间尺度也不同[9]:箱法的时间尺度是h—a;EC法可以精确到30 min,而时间跨度可达10 a以上;测树学法更适合碳库长期变化监测。因此本文只讨论前3种实测方法。
2.2.1 测树学方法测树学方法可以测量NPP和NECB,其优点是设备和技术要求低,可以实现长期监测。但往往只能得到年尺度至年代尺度的碳通量,时间分辨率低。由于NPP和NECB的概念和基本假设均不同,用测树学方法测量森林生态系统碳通量时应当注意二者的区别,否则容易让读者产生误解。下面分别阐述其测量方法。
森林生态系统NPP最简单也是最常见的测量方法是通过估计一定间隔期内(年到数年尺度)生物量碳的增加量实现的[43, 69],常用单位g biomass m-2 a-1或g C m-2 a-1表示[70]。但这种方法常因忽略一些组分(主要是测量期间的生产力转化)而低估NPP[43, 69]。为了澄清基本概念并提出一个完整的框架,Clark等[69]定义NPP*为实际NPP的野外测量估计值,它是以下两部分的总和:(1)在测量间隔末期活体植物保存下来的新有机质的量,(2)同期内植物生产但又损失的有机质的量。根据国际最新研究进展,在原框架的基础上对内容略作改动(图 2)。
(1)净初级生产力
①生物量增长量的测定 乔木直径变化可以通过连续每木检尺或者钻取生长芯测量年轮宽度两种方法获得。挂牌样地适宜根据每一样木的情况单独计算活立木的净增量和进级木(前一次测量小于起测径级,而该次测量大于起测径级的树木)的生长量。而没有挂牌样地适宜计算整个样地的活立木生物量之差,加上测量期间死亡的立木生物量,再减去所有进级木的起测径级生物量之和。如果出现单株负增长的情况,可能是测量误差所致,也可能是树木活力下降而逐渐失水导致的收缩,净增量可记为零[71]。为了减小误差,可用直径生长测量仪。起测径级应该根据样地树木的径级分布情况和工作量来确定。在计算时,进级木增量应减去起测径级的生物量碳。小于起测径级的灌木和乔木幼树,其NPP也需要隔年测定,或者利用生物量碳除以平均年龄粗略估算。

②测量间隔期的设定受诸多条件的限制,最重要的是避免严重低估NPP 挂牌样地推荐每年或隔年测量1次,而地处偏远的样地更适宜2—5a间隔期。枯死木的处理不当会引起更大的误差[43],测量期间死亡的样木实质上是从活体植物碳库转移到植物残体碳库,相当于负正相互抵消而没有变化(NPP不包括分解作用释放的碳)。如果两次测量间隔期较长,将枯死木生长量视为零可能低估NPP,而且两次测量期间产生的枯倒木将会带来更大的误差。
③地上凋落物是非常重要的NPP组分 相当一部分叶片(常绿针叶树种的一部分和落叶树种的全部)、小枝、繁殖器官和树皮在测量期间脱落,这是异速生长方程无法考虑的。落叶树种的叶NPP通常直接等同于凋落叶量,小枝、繁殖器官和树皮的凋落量一般也采用凋落物收集法估计。枯立木凋落的嫩枝不能计算在内,否则就会因为重复计算枯损量而高估NPP。凋落物收集应该尽量避免收集间隔期的分解导致的质量损失[69]。特别地,粗枝枯损量具有很大时空变异和偶发性,不宜用取样面积过小的凋落物收集器估计,而应该在更长的时间尺度上采用更大的取样面积估计[44]。这部分NPP并不是新近产生而是多年前的光合产物积累的,如果测量间隔很短(如1a)误差就会较大。
④地上取食量包括叶、花和果实取食以及树液吸食,是最常忽略的NPP组分之一 这部分NPP在健康森林中所占比例很小,但当虫害爆发或存在明显的植食脊椎动物种群时不能忽略[72]。一般叶取食量可以通过叶面积损失比例或粪便与取食量的关系估算[73]。
⑤生物挥发性有机化合物(BVOC) BVOC通量可用箱法或涡度协方差法测量[74]。例如大齿杨(Populus grandidentata)森林生态系统的BVOC通量在5.2—9.9 g C m-2 a-1,约占NPP的0.8%—1.6%[75]。
⑥淋溶有机质 NPP中另一个很少量化的组分是地上淋溶有机质,主要指穿透雨和树干茎流淋洗的植物分泌的有机物,包括可溶性有机碳(DOC)和颗粒有机碳(POC)。淋溶有机碳通常需要通过收集林内雨量和树干茎流量并测定其溶解的碳浓度估计。如果大气干湿沉降的碳输入明显[28],就要减去这部分额外的碳输入。据研究,美国Hubbard Brook站大气碳沉降仅为1.7 g C m-2 a-1,林冠层穿透雨净淋溶的碳为3.1 g C m-2 a-1[76]。
⑦粗根NPP* 粗根净增量与地上生物量净增量测定一致,缺少粗根生物量方程时可以通过根冠比估计。测定期间死亡的树木粗根相对容易估计,但活立木的粗根枯损量的确定比粗枝枯损量更加困难。Fahey等[76]用根冠比代表地下木质组织(粗根)与地上木质组织(枝凋落物)枯损量的比值,再根据地上木质枯损量粗略估计粗根枯损量为18 g C m-2 a-1。
⑧细根NPP* 它是测量最困难也是不确定性最大的NPP*组分,实践中通常将细根生物量增量和枯损量一起估算。根系向共生菌输出的NSC是枯损量中难以估计的组分。各种方法之间的详细比较可以参阅文献[77, 78, 79, 80, 81, 82]。根钻法[37]、内生长芯法[83]、碳平衡法[84]、氮平衡法[85]、微根管法[86]和放射性碳示踪法[79]各有利弊,笔者推荐采用多种途径交互验证,其中地下碳分配可以作为细根NPP*的上限约束其他方法[84]。
⑨林下草本NPP* 草本植物生物量最大值通常并不等于NPP,因为很大一部分生物量产生在测量期间凋落甚至分解[87]。Scurlock等[87]比较6种草本NPP的计算方法(生物量最大值、现存量最大值、生物量极差、生物量正增长量之和、现存量净增量之和与经过分解量校正的现存量变化量之和)认为,不同方法影响NPP*,考虑凋落物动态的估计是生物量最大值的2—5倍。而地上多年生草本植物来说,生物量最大值可能高估NPP。
(2)净生态系统碳平衡
采用测树学方法测量NECB,即相当于单位时间内生态系统各碳库变化量之和:
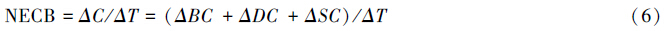
式中,ΔC表示生态系统碳库变化量,ΔBC、ΔDC和ΔSC分别表示植被、植物残体和土壤碳库的变化量,ΔT表示测量间隔期。据估计,我国森林表土SOC密度在20世纪最后20年间平均增长率为20 g C m-2 a-1[88]。由于高背景值和空间变异,在短期内难以检测到土壤碳库的显著变化,每10a测定1次较为合适[89]。有时候可采用年龄系列法粗略估计土壤碳库的变化速率,但绝大多数研究变化不显著[90]。因此迫切需要原位测定方法来检测土壤碳库随时间的变化[91]。非弹性中子散射技术就是一种革命性的土壤碳质量分数的原位检测方法[92]。
2.2.2 箱法箱法可以测量叶片、枝条、树干和根系以及土壤等生态系统某一组分的气体交换,涉及到光合作用(透明箱)与呼吸作用(暗箱)等众多生理生态学参数,因此又称为生理生态学方法。该方法采用不同类型的箱体将土壤、植被或植被的一部分密封,通过测定单位时间箱体内气体干摩尔分数的变化来计算研究对象的气体交换速率[64],可以测量GPP和Re及其组分(Ra和Rh)。需要指出的是,NEP和GPP有时候是由测树学法和箱法相结合测定的。由于凋落物和土壤碳库的变化难以在年尺度上准确测定,通常采用公式(3)估计NEP,即由测树学法的NPP与箱法的Rh之差计算得到。而公式(1)则表明了GPP可由NPP和Ra之和计算。由于成本相对低廉(但全自动箱系统价格昂贵)、构建简单、技术难度较低且便于操作实施,箱式气体交换系统广泛应用于陆地生态系统,在森林生态系统主要面临尺度上推问题。
根据气体的流动性和CO2测量方法,箱法的分类多种多样[64]。其中动态密闭箱-红外气体分析仪法的精度高且适用对象多,是目前最流行的箱式测量系统;叶片光合作用的控制测量则以开放箱-红外气体分析仪法(LI-6400,LI-COR,USA)为主。箱法固有的问题在于气体交换箱的密闭会影响测定对象的自然环境(辐射、温度、湿度和风速等)所产生的干扰作用,因此控制箱内的温度和湿度很重要。叶片尺度的气体交换还受到夹式叶室(Clamp-on leaf cuvette)的横向扩散泄漏效应的影响,在叶片大于叶室面积时容易低估光合作用速率而高估暗呼吸速率[93, 94]。箱体效应可通过优化箱体和采样设计(如增加通气孔平衡箱体内外气压)和缩短测量时间来降低[62]。
从小尺度的局部测量推算整个森林生态系统的碳通量是箱法的另一大难题[62]。例如利用箱法估算光合作用至少需要光在冠层内的传输模拟、叶面积指数(LAI)和空气温湿度的动态数据[95],而估算冠层叶片呼吸至少需要叶片(或空气)温度和LAI动态数据。直接测定的枝干呼吸为单位树干表面积的瞬时呼吸速率,存在巨大的时空变异[96],上推到生态系统尺度需要枝干表面积指数或林分的边材体积[97]或氮含量[98],如果是间歇性测量还要通过呼吸速率与树干或空气温度的动态监测数据才能实现时间积分估算年呼吸量。CWD呼吸可以基于体积或现存量实现空间尺度上推。土壤呼吸(Rs)尺度上推只需考虑空间代表性[62]和时间积分(呼吸模型的选择)产生的误差[99]。近年来,多通道Rs自动连续监测系统(如LI-8150,LI-COR,USA)大大减小了箱体效应,而且充分考虑空间变异,最大限度地获取连续数据集,因此极大地提高了Rs估算的可靠性[100]。
由于Ra和Rh的影响因素不同,区分组分是Rs研究的重要课题[101]。Rs包括植物根系、根际微生物、自由微生物和土壤动物呼吸几大组分。土壤Ra和Rh的区分方法可参阅文献[27, 102, 103],其中森林生态系统应用最普遍的方法是挖壕法[103]。该方法采用挖壕沟切断植物根系,并用塑料布等材料隔离新根侵入。笔者推荐塑料布作为隔断新根侵入的材料,尽管这种材料不透水,但可以有效避免对照区域的高浓度CO2扩散到挖壕样方高估异养呼吸组分。该方法的缺点是人为切断根系导致额外增加的死根分解产生的CO2通量而高估Rh[27, 103],挖壕处理对土壤环境因子的改变(诸如温度和水分)也产生间接影响[104]。挖壕处理实施后的1—2a内测定的CO2通量明显高于实际Rh[105],这可能是我国Rs组分区分最大的问题。但随着测定时间延长,该效应会由于底物减少而逐渐减弱。另一方面,挖壕切断了细根周转过程中产生的碳出入,因此挖壕样方的CO2通量缺乏新鲜细根凋落物分解的贡献,是造成Rh低估的潜在因素。
2.2.3 涡度协方差方法EC法是微气象学方法的代表,它可以直接估算陆地生态系统NEE,还可以通过通量拆分估计GPP和Re。该方法的优势是研究尺度较大,直接测定碳通量,可以自动长期连续精细观测。缺点在于仪器昂贵,对研究人员技术要求高,数据处理复杂,在复杂地形容易受到泄流影响。该方法的详细原理可参阅文献[14, 29]和专著[11, 12, 106, 107]。
首先EC通量观测地点的选取应该遵循地形尽量平坦、通量风浪区(Fetch)足够大、植被异质性小等原则。EC系统安装高度一般以1.5—3倍冠层高度为宜。地形复杂和风浪区较小时安装高度可适当降低,但过低时将导致仪器受植被粗糙单元影响过大而产生严重的系统偏差。EC通量观测的基本原理是单点的物质守恒方程[11],包括湍流通量、储存通量和平流通量3项。通量观测的标准配置为单套EC系统(测量CO2湍流通量)和CO2混合比廓线系统测量(测量CO2储存通量),CO2平流通量则忽略[11, 12, 29]。由于森林多处于坡面和山谷地形,由中尺度环流(比如山谷风)引发的气流辐合(辐散)常常导致明显的平流通量是EC技术面临的最重要的挑战之一[108]。
由高频(通常10HZ)时间序列计算原始通量必需经过一系列的校正[12]。这些校正主要包括:野点去除、坐标旋转、超声虚温校正、仪器频率响应、管路延时和密度校正(通常忽略气压脉动,又称WPL校正)。在温度较低时,旧式开路红外气体分析仪LI-7500(LI-COR,USA)的加热效应也不容忽略,对历史数据的经验校正方法参阅文献[109],但当LI-7500非垂直安装时的误差还难以估计。数据质量控制也是必不可少的,野点去除、稳态测试、湍流积分测试和通量贡献区控制是最重要的内容[11, 110]。
恶劣天气条件、设备维护、仪器损坏和停电以及数据质量控制等原因,会导致缺失一部分通量半小时数据。在晴朗无风(天气尺度)的夜晚,常常因为平流贡献明显而低估Re。常用的方法是摩擦风速阈值过滤这些无效数据[111, 112]。若无法检测到摩擦风速阈值,可采用夜间最大呼吸法确定有效数据[113],该方法的基本原理是夜间初期平流作用尚未充分发展,利用湍流通量和储存通量可充分代表NEE[113]。为了得到日、月、年和年代尺度的NEE,必须进行数据插补[114, 115]。数据插补最基本的原则是误差最小化。对于小于2 h的缺失数据,可用线性内插法估计。而长时间缺失数据常用的插补方法有:平均日变化法、查表法、非线性回归法、边缘分布抽样和人工神经网络法等[114],其中最可靠的方法通常是人工神经网络、非线性回归和边缘分布抽样法[114]。
EC技术直接观测的是NEE,要得到GPP和Re就必须进行通量拆分[11, 116, 117]。夜间和非生长季的GPP为零,因此NEE等于Re;根据公式(5),NEE是-GPP与Re之和。光合作用只发生在生长季白天,因而此时Re的准确估计是通量拆分的关键。通量拆分有两种方法:一种是夜间呼吸外推法,将夜间测定的有效NEE(即Re)通过温度响应模型外推得到白天Re[116];另一种是光响应曲线法,利用白天NEE的冠层光合作用光响应模型估计白天Re[117]。夜间呼吸外推法中有效数据筛选、呼吸模型的类型和温度变量的选择均会影响数据插补误差[118],也会影响通量拆分。光响应曲线法是建立NEE与光合有效辐射的关系方程,当光合有效辐射为零时的NEE即为Re。叶片尺度的模型[119]在冠层尺度均可应用。由于叶片光合作用还受到水汽压饱和亏缺的影响,而暗呼吸速率受温度的影响,因此在模型中考虑水汽压饱和亏缺和空气温度将更好地估计白天的Re[117]。
EC法的不确定性主要包括测量随机误差、通量校正误差、通量数据异常值剔除、数据插补策略和通量拆分[11],其中通量数据筛选和数据插补策略是最重要的误差源[110, 120, 121, 122]。除了不确定性分析之外,该技术最关键的问题是避免NEE的系统误差,比如夜间数据普遍低估Re和不恰当的原始通量数据校正方法。此外,能量平衡不闭合可能反映能量通量(感热和潜热通量)系统低估[123],由于CO2通量与能量的传输过程相似,因此有研究[110, 124]推荐在计算NEE时应该校正这一系统误差。
3 森林碳循环研究的主要问题和建议(1) 生物量方程建模不规范,应用尚缺乏验证。生物量方程选择不当不仅给植被碳储量估计带来误差,还会影响到NPP和NEP的估计,因此必须予以重视。一方面,部分异速生长方程建模时并没有给出DBH区间,选择幂函数模型时也没有考虑异方差的影响。20世纪末冯宗炜等[125]系统整理了我国主要树种的生物量异速生长方程,但当时的数据库存在一些不足(比如树种和地区覆盖不全面,模型形式和建模方法可能存在误差,DBH范围缺乏或过小),限制了模型的准确性和可用性。近10年来我国森林生物量方程取得了长足进展,有必要按地区系统整理、比较和验证现有生物量异速生长方程,为生物量方程的选择提供客观依据。另一方面,由于时间和工作量的限制,很多研究在引用现有生物量模型时缺乏比较和验证,这也可能造成较大的误差。
(2) 森林碳循环参数测量不完整,限制了不同研究之间的比较和数据整合分析。例如,Luyssaert等[7]收集了全球森林碳通量数据集,报道了林下植被NPP的文献不足30%,而估计了果实和繁殖器官NPP的研究不足4%,几乎没有研究报道VOC和渗流的碳输出。虽然因实际困难而很难保证测量NPP的所有组分,但笔者建议报道NPP时需要明确指出忽略的组分。在区域尺度上验证模型结果时,应该注意所收集的验证数据是否完整,因为这直接影响估计结果的可靠性。此外,将未受明显干扰的森林生态系统的NEP等同于NECB,可能会高估陆地生态系统的碳汇[26, 126]。在典型森林生态系统的长期监测样地开展碳通量组分的全面精细测量,进而为周边地区相似森林生态系统提供校正参数,可以大大提高区域尺度上NPP估算的可靠性。
(3) 森林碳循环参数的实测数值和不确定性影响区域和全球森林碳汇的估算精度。大尺度的森林碳收支估测精度在很大程度上取决于生态系统尺度的测量以及这些观测点的空间分布[108],但在生态系统尺度上各种途径的碳循环数据的不确定性评估还不够完善。例如,测树学方法的4个误差源(空间代表性、异速生长方程选择和应用、DBH测量和碳质量分数)、箱法的测量误差和尺度上推的不确定性以及EC技术的不确定性和系统误差研究尚不多见。因此,森林生态系统尺度碳循环实测数据的误差和不确定性的评估是森林碳循环研究必不可少的内容。
(4) 绝大多数森林碳循环研究只采用一种方法,碳通量的多途径估计还十分匮乏[9]。国外一些站点系统比较了EC法与测树学法测定的NEP[127, 128]。Piao等[129]用3种途径估计我国陆地生态系统的碳汇在0.19—0.26 Pg/a之间,不同方法之间仍然存在一定差距。我国基于EC方法的森林碳通量观测正在蓬勃发展[130],但这些观测大多还没有与测树学方法或箱式法的NEP相互比较验证,仅长白山站[131]、西双版纳站[132]和哀牢山站[133]比较了不同途径的实测NEP。全球范围森林生态系统的测树学和和EC法的比较发现,后者NEP总体上大于前者,表明EC法倾向于高估森林碳汇[134]。在有条件的典型森林生态系统开展碳循环多途径交互验证,有助于降低碳通量测量的误差,进而为碳循环模型和区域乃至全球尺度碳平衡估算提供可靠的数据。
| [1] | Jobbágy E G, Jackson R B. The vertical distribution of soil organic carbon and its relation to climate and vegetation. Ecological Applications, 2000, 10(2): 423-436. |
| [2] | Pan Y D, Birdsey R A, Fang J, Houghton R, Kauppi P E, Kurz W A, Phillips O L, Shvidenko A, Lewis S L, Canadell J G, Ciais P, Jackson R B, Pacala S W, McGuire A D, Piao S L, Rautiainen A, Sitch S, Hayes D. A large and persistent carbon sink in the world's forests. Science, 2011, 333(6045): 988-993. |
| [3] | Ruesch A, Gibbs H K. New IPCC Tier-1 Global Biomass Carbon Map for the Year 2000. Oak Ridge National Laboratory, Oak Ridge, Tennessee: Carbon Dioxide Information Analysis Center, 2008. |
| [4] | Beer C, Reichstein M, Tomelleri E, Ciais P, Jung M, Carvalhais N, Rödenbeck C, Arain M A, Baldocchi D, Bonan G B, Bondeau A, Cescatti A, Lasslop G, Lindroth A, Lomas M, Luyssaert S, Margolis H, Oleson K W, Roupsard O, Veenendaal E, Viovy N, Williams C, Woodward F I, Papale D. Terrestrial gross carbon dioxide uptake: global distribution and covariation with climate. Science, 2010, 329(5993): 834-838. |
| [5] | Le Quéré C, Andres R J, Boden T, Conway T, Houghton R A, House J I, Marland G, Peters G P, van der Werf G R, Ahlström A, Andrew R M, Bopp L, Canadell J G, Ciais P, Doney S C, Enright C, Friedlingstein P, Huntingford C, Jain A K, Jourdain C, Kato E, Keeling R F, Klein Goldewijk K, Levis S, Levy P, Lomas M, Poulter B, Raupach M R, Schwinger J, Sitch S, Stocker B D, Viovy N, Zaehle S, Zeng N. The global carbon budget 1959-2011. Earth System Science Data, 2013, 5: 165-185. |
| [6] | Canadell J G, Le Quéré C, Raupach M R, Field C B, Buitenhuis E T, Ciais P, Conway T J, Gillett N P, Houghton R, Marland G. Contributions to accelerating atmospheric CO2 growth from economic activity, carbon intensity, and efficiency of natural sinks. Proceedings of the National Academy of Sciences of the United States of America, 2007, 104(47): 18866-18870. |
| [7] | Luyssaert S, Inglima I, Jung M, Richardson A D, Reichstein M, Papale D, Piao S L, Schulze E D, Wingate L, Matteucci G, Aragao L, Aubinet M, Beer C, Bernhofer C, Black K G, Bonal D, Bonnefond J M, Chambers J, Ciais P, Cook B, Davis K J, Dolman A J, Gielen B, Goulden M, Grace J, Granier A, Grelle A, Griffis T, Grünwald T, Guidolotti G, Hanson P J, Harding R, Hollinger D Y, Hutyra L R, Kolari P, Kruijt B, Kutsch W, Lagergren F, Laurila T, Law B E, Le Maire G, Lindroth A, Loustau D, Malhi Y, Mateus J, Migliavacca M, Misson L, Montagnani L, Moncrieff J, Moors E, Munger J W, Nikinmaa E, Ollinger SV, Pita G, Rebmann C, Roupsard O, Saigusa N, Sanz M J, Seufert G, Sierra C, Smith M L, Tang J, Valentini R, Vesala T, Janssens I A. CO2 balance of boreal, temperate, and tropical forests derived from a global database. Global Change Biology, 2007, 13(12): 2509-2537. |
| [8] | 方精云, 柯金虎, 唐志尧, 陈安平. 生物生产力的"4P"概念、估算及其相互关系. 植物生态学报, 2001, 25(4): 414-419. |
| [9] | 于贵瑞, 王秋凤, 朱先进. 区域尺度陆地生态系统碳收支评估方法及其不确定性. 地理科学进展, 2011, 30(1): 103-113. |
| [10] | 赵德华, 李建龙, 齐家国, 范亚民. 陆地生态系统碳平衡主要研究方法评述. 生态学报, 2006, 26(8): 2655-2662. |
| [11] | Aubinet M, Vesala T, Papale D. Eddy Covariance: A Practical Guide to Measurement and Data Analysis. 2nd ed. Berlin: Springer, 2012. |
| [12] | Burba G. Eddy Covariance Method for Scientific, Industrial, Agricultural and Regulatory Applications: A Field Book on Measuring Ecosystem Gas Exchange and Areal Emission Rates. Lincoln, Nebraska: LI-COR Biosciences, 2013. |
| [13] | Kowalski A, Serrano-Ortiz P. On the relationship between the eddy covariance, the turbulent flux, and surface exchange for a trace gas such as CO2. Boundary-Layer Meteorology, 2007, 124(2): 129-141. |
| [14] | Gu L H, Massman W J, Leuning R, Pallardy S G, Meyers T, Hanson P J, Riggs J S, Hosman K P, Yang B. The fundamental equation of eddy covariance and its application in flux measurements. Agricultural and Forest Meteorology, 2012, 152: 135-148. |
| [15] | 王静, 王兴昌, 王传宽. 基于不同浓度变量的温带落叶阔叶林CO2储存通量的误差分析. 应用生态学报, 2013, 24(4): 975-982. |
| [16] | Thomas S C, Martin A R. Carbon content of tree tissues: a synthesis. Forests, 2012, 3(2): 332-352. |
| [17] | 杨金艳, 王传宽. 东北东部森林生态系统土壤碳贮量和碳通量. 生态学报, 2005, 25(11): 2875-2882. |
| [18] | Hoch G, Richter A, Körner C. Non-structural carbon compounds in temperate forest trees. Plant, Cell & Environment, 2003, 26(7): 1067-1081. |
| [19] | 张海燕, 王传宽, 王兴昌. 温带12个树种新老树枝非结构性碳水化合物浓度比较. 生态学报, 2013, 33(18): 5675-5685. |
| [20] | 张全智, 王传宽. 6种温带森林碳密度与碳分配. 中国科学: 生命科学, 2010, 40(7): 621-631. |
| [21] | 周玉荣, 于振良, 赵士洞. 我国主要森林生态系统碳贮量和碳平衡. 植物生态学报, 2000, 24(5): 518-522. |
| [22] | 王绍强, 刘纪远, 于贵瑞. 中国陆地土壤有机碳蓄积量估算误差分析. 应用生态学报, 2003, 14(5): 797-802. |
| [23] | 王绍强, 周成虎, 李克让, 朱松丽, 黄方红. 中国土壤有机碳库及空间分布特征分析. 地理学报, 2000, 55(5): 533-544. |
| [24] | Litton C M, Raich J W, Ryan M G. Carbon allocation in forest ecosystems. Global Change Biology, 2007, 13(10): 2089-2109. |
| [25] | Chapin F S, Matson P A, Mooney H A. Principles of Terrestrial Ecosystem Ecology. New York: Springer, 2002. |
| [26] | Chapin F S, Woodwell G, Randerson J T, Rastetter E B, Lovett G, Baldocchi D D, Clark D, Harmon M E, Schimel D S, Valentini R, Wirth C, Aber J D, Cole J J, Goulden M L, Harden J W, Heimann M, Howarth R W, Matson P A, McGuire A D, Melillo J M, Mooney H A, Neff J C, Houghton R A, Pace M L, Ryan M G, Running S W, Sala O E, Schlesinger W H, Schulze E D. Reconciling carbon-cycle concepts, terminology, and methods. Ecosystems, 2006, 9(7): 1041-1050. |
| [27] | Kuzyakov Y. Sources of CO2 efflux from soil and review of partitioning methods. Soil Biology and Biochemistry, 2006, 38(3): 425-448. |
| [28] | 孙忠林, 王传宽. 森林生态系统可溶性碳和颗粒碳通量. 生态学报, 2014, 34(15): 4133-4141. |
| [29] | Baldocchi D D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: past, present and future. Global Change Biology, 2003, 9(4): 479-492. |
| [30] | 王兴昌, 王传宽, 于贵瑞. 基于全球涡度相关的森林碳交换的时空格局. 中国科学 D辑: 地球科学, 2008, 38(9): 1092-1108. |
| [31] | Lorenz K, Lal R. Carbon Sequestration in Forest Ecosystems. New York: Springer, 2010. |
| [32] | Lal R. Carbon sequestration. Philosophical Transactions of the Royal Society B: Biological Sciences, 2008, 363(1492): 815-830. |
| [33] | Poorter H, Niklas K J, Reich P B, Oleksyn J, Poot P, Mommer L. Biomass allocation to leaves, stems and roots: meta-analyses of interspecific variation and environmental control. New Phytologist, 2012, 193(1): 30-50. |
| [34] | Mokany K, Raison R, Prokushkin A S. Critical analysis of root: shoot ratios in terrestrial biomes. Global Change Biology, 2006, 12(1): 84-96. |
| [35] | Chen G S, Yang Y S, Robinson D. Allocation of gross primary production in forest ecosystems: allometric constraints and environmental responses. New Phytologist, 2013, 200(4): 1176-1186. |
| [36] | Gill R A, Jackson R B. Global patterns of root turnover for terrestrial ecosystems. New Phytologist, 2000, 147(1): 13-31. |
| [37] | Brunner I, Bakker M R, Björk R G, Hirano Y, Lukac M, Aranda X, Børja I, Eldhuset T D, Helmisaari H S, Jourdan C, Konôpka B, López B C, Miguel Pérez C, Persson H, Ostonen I. Fine-root turnover rates of European forests revisited: an analysis of data from sequential coring and ingrowth cores. Plant and Soil, 2013, 362(1/2): 357-372. |
| [38] | Stockmann U, Adams M A, Crawford J W, Field D J, Henakaarchchi N, Jenkins M, Minasny B, McBratney A B, de Remy de Courcelles V, Singh K, Wheelera I, Abbottb L, Angersc D A, Baldockd J, Birde M, Brookesf P C, Chenug C, Jastrowh J D, Lali R, Lehmannj J, O'Donnellk A G, Partonl W J, Whiteheadm D, Zimmermannn M. The knowns, known unknowns and unknowns of sequestration of soil organic carbon. Agriculture, Ecosystems & Environment, 2013, 164: 80-99. |
| [39] | Gaudinski J B, Trumbore S E, Davidson E A, Zheng S H. Soil carbon cycling in a temperate forest: radiocarbon-based estimates of residence times, sequestration rates and partitioning of fluxes. Biogeochemistry, 2000, 51(1): 33-69. |
| [40] | 罗云建, 张小全, 王效科, 朱建华, 侯振宏, 张治军. 森林生物量的估算方法及其研究进展. 林业科学, 2009, 45(8): 129-134. |
| [41] | 谢亭亭, 李根, 周光益, 吴仲民, 赵厚本, 邱治军, 梁瑞友. 南岭小坑小红栲-荷木群落的地上生物量. 应用生态学报, 2013, 24(9): 2399-2407. |
| [42] | Saatchi S S, Harris N L, Brown S, Lefsky M, Mitchard E T, Salas W, Zutta B R, Buermann W, Lewis S L, Hagen S, Petrova S, White L, Silman M, Morel A. Benchmark map of forest carbon stocks in tropical regions across three continents. Proceedings of the National Academy of Sciences of the United States of America, 2011, 108(24): 9899-9904. |
| [43] | Gower S T, Kucharik C J, Norman J M. Direct and indirect estimation of leaf area index, fAPAR, and net primary production of terrestrial ecosystems. Remote Sensing of Environment, 1999, 70(1): 29-51. |
| [44] | Hoover C M. Field Measurements for Forest Carbon Monitoring. Netherlands: Springer, 2008. |
| [45] | Keller M, Palace M, Hurtt G. Biomass estimation in the Tapajos National Forest, Brazil: examination of sampling and allometric uncertainties. Forest Ecology and Management, 2001, 154(3): 371-382. |
| [46] | Chave J, Condit R, Aguilar S, Hernandez A, Lao S, Perez R. Error propagation and scaling for tropical forest biomass estimates. Philosophical Transactions of the Royal Society of London. Series B: Biological Sciences, 2004, 359(1443): 409-420. |
| [47] | Ketterings Q M, Coe R, van Noordwijk M, Ambagau Y, Palm C A. Reducing uncertainty in the use of allometric biomass equations for predicting above-ground tree biomass in mixed secondary forests. Forest Ecology and Management, 2001, 146(1/3): 199-209. |
| [48] | 曾伟生, 唐守正. 国外立木生物量模型研究现状与展望. 世界林业研究, 2010, 23(4): 30-35. |
| [49] | Wang C K. Biomass allometric equations for 10 co-occurring tree species in Chinese temperate forests. Forest Ecology and Management, 2006, 222(1/3): 9-16. |
| [50] | Butt N, Slade E, Thompson J, Malhi Y, Riutta T. Quantifying the sampling error in tree census measurements by volunteers and its effect on carbon stock estimates. Ecological Applications, 2013, 23(4): 936-943. |
| [51] | 马钦彦, 陈遐林, 王娟, 蔺琛, 康峰峰, 曹文强, 马志波, 李文宇. 华北主要森林类型建群种的含碳率分析. 北京林业大学学报, 2002, 24(5/6): 96-100. |
| [52] | Zhang Q Z, Wang C K, Wang X C, Quan X K. Carbon concentration variability of 10 Chinese temperate tree species. Forest Ecology and Management, 2009, 258(5): 722-727. |
| [53] | Harmon M E, Franklin J F, Swanson F J, Sollins P, Gregory S, Lattin J, Anderson N, Cline S, Aumen N, Sedell J, Lienkaemper G, Kermit Cromack J, Cummins K. Ecology of coarse woody debris in temperate ecosystems. Advances in Ecological Research, 1986, 15: 133-302. |
| [54] | 闫恩荣, 王希华, 黄建军. 森林粗死木质残体的概念及其分类. 生态学报, 2005, 25(1): 158-167. |
| [55] | Woldendorp G, Keenan R J, Barry S, Spencer R D. Analysis of sampling methods for coarse woody debris. Forest Ecology and Management, 2004, 198(1/3): 133-148. |
| [56] | Williams M S, Gove J H. Perpendicular distance sampling: an alternative method for sampling downed coarse woody debris. Canadian Journal of Forest Research, 2003, 33(8): 1564-1579. |
| [57] | Harmon M E, Fasth B, Woodall C W, Sexton J. Carbon concentration of standing and downed woody detritus: Effects of tree taxa, decay class, position, and tissue type. Forest Ecology and Management, 2013, 291: 259-267. |
| [58] | Yang Y H, Mohammat A, Feng J M, Zhou R, Fang Y. Storage, patterns and environmental controls of soil organic carbon in China. Biogeochemistry, 2007, 84(2): 131-141. |
| [59] | Pribyl D W. A critical review of the conventional SOC to SOM conversion factor. Geoderma, 2010, 156(3/4): 75-83. |
| [60] | Law B E, Ryan M G, Anthoni P M. Seasonal and annual respiration of a ponderosa pine ecosystem. Global Change Biology, 1999, 5(2): 169-182. |
| [61] | Lavigne M B, Ryan M G, Anderson D E, Baldocchi D D, Crill P M, Fitzjarrald D R, Goulden M L, Gower S T, Massheder J M, McCaughey J H, Rayment M, Striegl R G. Comparing nocturnal eddy covariance measurements to estimates of ecosystem respiration made by scaling chamber measurements at six coniferous boreal sites. Journal of Geophysical Research, 1997, 102(D24): 28977-28985. |
| [62] | Davidson E A, Savage K, Verchot L V, Navarro R. Minimizing artifacts and biases in chamber-based measurements of soil respiration. Agricultural and Forest Meteorology, 2002, 113(1/4): 21-37. |
| [63] | Xu M, DeBiase TA, Qi Y, Goldstein A, Liu Z G. Ecosystem respiration in a young ponderosa pine plantation in the Sierra Nevada Mountains, California. Tree Physiology, 2001, 21(5): 309-318. |
| [64] | 袁凤辉, 关德新, 吴家兵, 王安志, 金昌杰. 箱式气体交换观测系统及其在植物生态系统气体交换研究中的应用. 应用生态学报, 2009, 20(6): 1495-1504. |
| [65] | Running S W, Nemani R R, Heinsch F A, Zhao M S, Reeves M, Hashimoto H. A continuous satellite-derived measure of global terrestrial primary production. BioScience, 2004, 54(6): 547-560. |
| [66] | Ciais P, Rayner P, Chevallier F, Bousquet P, Logan M, Peylin P, Ramonet M. Atmospheric inversions for estimating CO2 fluxes: methods and perspectives. Climatic Change, 2010, 103(1/2): 69-92. |
| [67] | Cramer W, Bondeau A, Woodward F I, Prentice I C, Betts R A, Brovkin V, Cox P M, Fisher V, Foley J A, Friend A D, Kucharik C, Lomas M R, Ramankutty N, Sitch S, Smith B, White A, Young-Molling C. Global response of terrestrial ecosystem structure and function to CO2 and climate change: results from six dynamic global vegetation models. Global Change Biology, 2001, 7(4): 357-373. |
| [68] | Piao S, Sitch S, Ciais P, Friedlingstein P, Peylin P, Wang X H, Ahlström A, Anav A, Canadell J G, Cong N, Huntingford C, Jung M, Levis S, Levy P E, Li J S, Lin X, Lomas M R, Lu M, Luo Y Q, Ma Y C, Myneni R B, Poulter B, Sun Z Z, Wang T, Viovy N, Zaehle S, Zeng N. Evaluation of terrestrial carbon cycle models for their response to climate variability and to CO2 trends. Global Change Biology, 2013, 19(7): 2117-2132. |
| [69] | Clark D A, Brown S, Kicklighter D W, Chambers J Q, Thomlinson J R, Ni J. Measuring net primary production in forests: concepts and field methods. Ecological Applications, 2001, 11(2): 356-370. |
| [70] | Gower S T, Krankina O, Olson R J, Apps M, Linder S, Wang C. Net primary production and carbon allocation patterns of boreal forest ecosystems. Ecological Applications, 2001, 11(5): 1395-1411. |
| [71] | Ohtsuka T, Akiyama T, Hashimoto Y, Inatomi M, Sakai T, Jia S G, Mo W H, Tsuda S, Koizumi H. Biometric based estimates of net primary production (NPP) in a cool-temperate deciduous forest stand beneath a flux tower. Agricultural and Forest Meteorology, 2005, 134(1/4): 27-38. |
| [72] | Hicke J A, Allen C D, Desai A R, Dietze M C, Hall R J, Hogg E H, Kashian D M, Moore D, Raffa K F, Sturrock R N, Vogelmann J. Effects of biotic disturbances on forest carbon cycling in the United States and Canada. Global Change Biology, 2012, 18(1): 7-34. |
| [73] | 王晓伟, 姬兰柱, 王绪高, 安琳莉, 燕丽波. 森林冠层食叶昆虫取食水平的定量方法与研究进展. 生态学杂志, 2011, 30(7): 1403-1410. |
| [74] | Spirig C, Neftel A, Ammann C, Dommen J, Grabmer W, Thielmann A, Schaub A, Beauchamp J, Wisthaler A, Hansel A. Eddy covariance flux measurements of biogenic VOCs during ECHO 2003 using proton transfer reaction mass spectrometry. Atmospheric Chemistry and Physics, 2005, 5(2): 465-481. |
| [75] | Karl T, Guenther A, Spirig C, Hansel A, Fall R. Seasonal variation of biogenic VOC emissions above a mixed hardwood forest in northern Michigan. Geophysical Research Letters, 2003, 30(23), doi: 10.1029/2003GL018432. |
| [76] | Fahey T, Siccama T, Driscoll C, Likens G E, Campbell J, Johnson C E, Battles J J, Aber J D, Cole J J, Fisk M C, Groffman P M, Hamburg S P, Holmes R T, Schwarz P A, Yanai R D. The biogeochemistry of carbon at Hubbard Brook. Biogeochemistry, 2005, 75(1): 109-176. |
| [77] | Osawa A, Aizawa R. A new approach to estimate fine root production, mortality, and decomposition using litter bag experiments and soil core techniques. Plant and Soil, 2012, 355(1/2): 167-181. |
| [78] | Vogt K A, Vogt D J, Bloomfield J. Analysis of some direct and indirect methods for estimating root biomass and production of forests at an ecosystem level. Plant and Soil, 1998, 200(1): 71-89. |
| [79] | Strand A E, Pritchard S G, McCormack M L, Davis M A, Oren R. Irreconcilable differences: fine-root life spans and soil carbon persistence. Science, 2008, 319(5862): 456-458. |
| [80] | Guo D L, Li H B, Mitchell R J, Han W X, Hendricks J J, Fahey T J, Hendrick R L. Fine root heterogeneity by branch order: exploring the discrepancy in root turnover estimates between minirhizotron and carbon isotopic methods. New Phytologist, 2008, 177(2): 443-456. |
| [81] | Hendricks J J, Hendrick R L, Wilson C A, Mitchell R J, Pecot S D, Guo D L. Assessing the patterns and controls of fine root dynamics: an empirical test and methodological review. Journal of Ecology, 2006, 94(1): 40-57. |
| [82] | 全先奎, 于水强, 史建伟, 于立忠, 王政权. 微根管法和同位素法在细根寿命研究中的应用及比较. 生态学杂志, 2007, 26(3): 428-434. |
| [83] | Li X F, Zhu J, Lange H, Han S J. A modified ingrowth core method for measuring fine root production, mortality and decomposition in forests. Tree Physiology, 2013, 33(1): 18-25. |
| [84] | Nadelhoffer K J, Raich J W. Fine root production estimates and belowground carbon allocation in forest ecosystems. Ecology, 1992, 73(4): 1139-1147. |
| [85] | Aber J D, Melillo J M, Nadelhoffer K J, McClaugherty C A, Pastor J. Fine root turnover in forest ecosystems in relation to quantity and form of nitrogen availability: a comparison of two methods. Oecologia, 1985, 66(3): 317-321. |
| [86] | Johnson M G, Tingey D T, Phillips D L, Storm M J. Advancing fine root research with minirhizotrons. Environmental and Experimental Botany, 2001, 45(3): 263-289. |
| [87] | Scurlock J, Johnson K, Olson R. Estimating net primary productivity from grassland biomass dynamics measurements. Global Change Biology, 2002, 8(8): 736-753. |
| [88] | Yang Y H, Li P, Ding J Z, Zhao X, Ma W H, Ji C J, Fang J Y. Increased topsoil carbon stock across China's forests. Global Change Biology, 2014, 20(8): 2687-2696. |
| [89] | Schrumpf M, Schulze E D, Kaiser K, Schumacher J. How accurately can soil organic carbon stocks and stock changes be quantified by soil inventories?. Biogeosciences, 2011, 8(5): 1193-1212. |
| [90] | Yang Y H, Luo Y Q, Finzi A C. Carbon and nitrogen dynamics during forest stand development: a global synthesis. New Phytologist, 2011, 190(4): 977-989. |
| [91] | Chatterjee A, Lal R, Wielopolski L, Martin M Z, Ebinger M. Evaluation of different soil carbon determination methods. Critical Reviews in Plant Sciences, 2009, 28(3): 164-178. |
| [92] | Wielopolski L, Chatterjee A, Mitra S, Lal R. In situ determination of soil carbon pool by inelastic neutron scattering: Comparison with dry combustion. Geoderma, 2011, 160(3/4): 394-399. |
| [93] | Flexas J, Díaz-Espejo A, Berry J, Cifre J, Galmes J, Kaldenhoff R, Medrano H, Ribas-Carbó M. Analysis of leakage in IRGA's leaf chambers of open gas exchange systems: quantification and its effects in photosynthesis parameterization. Journal of Experimental Botany, 2007, 58(6): 1533-1543. |
| [94] | Rodeghiero M, Niinemets V, Cescatti A. Major diffusion leaks of clamp-on leaf cuvettes still unaccounted: how erroneous are the estimates of Farquhar et al. model parameters?. Plant, Cell & Environment, 2007, 30(8): 1006-1022. |
| [95] | Wang Y P, Leuning R. A two-leaf model for canopy conductance, photosynthesis and partitioning of available energy I: Model description and comparison with a multi-layered model. Agricultural and Forest Meteorology, 1998, 91(1/2): 89-111. |
| [96] | Ceschia É, Damesin C, Lebaube S, Pontailler J Y, Dufrêne É. Spatial and seasonal variations in stem respiration of beech trees (Fagus sylvatica). Annals of Forest Science, 2002, 59(8): 801-812. |
| [97] | Damesin C, Ceschia E, Le Goff N, Ottorini J M, Dufrêne E. Stem and branch respiration of beech: from tree measurements to estimations at the stand level. New Phytologist, 2002, 153(1): 159-172. |
| [98] | Lavigne M B, Franklin S E, Hunt E R Jr. Estimating stem maintenance respiration rates of dissimilar balsam fir stands. Tree Physiology, 1996, 16(8): 687-695. |
| [99] | Davidson E, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology, 1998, 4(2): 217-227. |
| [100] | Wang C K, Han Y, Chen J Q, Wang X C, Zhang Q Z, Bond-Lamberty B. Seasonality of soil CO2 efflux in a temperate forest: Biophysical effects of snowpack and spring freeze-thaw cycles. Agricultural and Forest Meteorology, 2013, 177: 83-92. |
| [101] | Trumbore S. Carbon respired by terrestrial ecosystems-recent progress and challenges. Global Change Biology, 2006, 12(2): 141-153. |
| [102] | Hanson P J, Edwards N T, Garten C T, Andrews J A. Separating root and soil microbial contributions to soil respiration: a review of methods and observations. Biogeochemistry, 2000, 48(1): 115-146. |
| [103] | Subke J A, Inglima I, Cotrufo M F. Trends and methodological impacts in soil CO2 efflux partitioning: a metaanalytical review. Global Change Biology, 2006, 12(6): 921-943. |
| [104] | Ngao J, Longdoz B, Granier A, Epron D. Estimation of autotrophic and heterotrophic components of soil respiration by trenching is sensitive to corrections for root decomposition and changes in soil water content. Plant and Soil, 2007, 301(1/2): 99-110. |
| [105] | 常建国, 刘世荣, 史作民, 陈宝玉, 朱学凌. 北亚热带-南暖温带过渡区典型森林生态系统土壤呼吸及其组分分离. 生态学报, 2007, 27(5): 1791-1802. |
| [106] | Lee X H, Massman W, Law B E. Handbook of Micrometeorology: A Guide for Surface Flux Measurement and Analysis. New York: Springer, 2004. |
| [107] | 于贵瑞, 孙晓敏. 陆地生态系统通量观测的原理与方法. 北京: 高等教育出版社, 2006. |
| [108] | Rotach M W, Wohlfahrt G, Hansel A, Reif M, Wagner J, Gohm A. The world is not flat: Implications for the global carbon balance. Bulletin of the American Meteorological Society, 2014, 95(7): 1021-1028. |
| [109] | Burba G G, McDermitt D K, Grelle A, Anderson D J, Xu L K. Addressing the influence of instrument surface heat exchange on the measurements of CO2 flux from open-path gas analyzers. Global Change Biology, 2008, 14(8): 1854-1876. |
| [110] | Mauder M, Cuntz M, Drüe C, Graf A, Rebmann C, Schmid H P, Schmidt M, Steinbrecher R. A strategy for quality and uncertainty assessment of long-term eddy-covariance measurements. Agricultural and Forest Meteorology, 2013, 169: 122-135. |
| [111] | Barr A G, Richardson A D, Hollinger D Y, Papale D, Arain M A, Black T A, Bohrer G, Dragoni D, Fischer M L, Gu L, Law B E, Margolis H A, McCaughey J H, Munger J W, Oechel W, Schaeffer K. Use of change-point detection for friction-velocity threshold evaluation in eddy-covariance studies. Agricultural and Forest Meteorology, 2013, 171-172: 31-45. |
| [112] | Goulden M L, Munger J W, Fan S M, Daube B C, Wofsy S C. Measurements of carbon sequestration by long-term eddy covariance: methods and a critical evaluation of accuracy. Global Change Biology, 1996, 2(3): 169-182. |
| [113] | van Gorsel E, Delpierre N, Leuning R, Black A, Munger J W, Wofsy S, Aubinet M, Feigenwinter C, Beringer J, Bonal D, Chen B Z, Chen J Q, Clement R, Davis K J, Desai A R, Dragoni D, Etzold S, Grünwald T, Gu L H, Heinesch B, Hutyra L R, Jans W W P, Kutsch W, Law B E, Leclerc M Y, Mammarella I, Montagnani L, Noormets A, Rebmann C, Wharton S. Estimating nocturnal ecosystem respiration from the vertical turbulent flux and change in storage of CO2. Agricultural and Forest Meteorology, 2009, 149(11): 1919-1930. |
| [114] | Moffat A M, Papale D, Reichstein M, Hollinger D Y, Richardson A D, Barr A G, Beckstein C, Braswell B H, Churkina G, Desai A R, Falge E, Gove J H, Heimann M, Hui D F, Jarvis A J, Kattge J, Noormets A, Stauch V J. Comprehensive comparison of gap-filling techniques for eddy covariance net carbon fluxes. Agricultural and Forest Meteorology, 2007, 147(3): 209-232. |
| [115] | Falge E, Baldocchi D, Olson R, Anthoni P, Aubinet M, Bernhofer C, Burba G, Ceulemans R, Clement R, Dolman H, Granier A, Gross P, Grünwald T, Hollinger D, Jensen N O, Katul G, Keronen P, Kowalski A, Lai C T, Law B E, Meyers T, Moncrieff J, Moors E, Munger J W, Pilegaard K, RannikV, Rebmann C, Suyker A, Tenhunen J, Tu K, Verma S, Vesala T. Gap filling strategies for defensible annual sums of net ecosystem exchange. Agricultural and Forest Meteorology, 2001, 107(1): 43-69. |
| [116] | Reichstein M, Falge E, Baldocchi D, Papale D, Aubinet M, Berbigier P, Bernhofer C, Buchmann N, Gilmanov T, Granier A, Grünwald T, Havránková K, Ilvesniemi H, Janous D, Knohl A, Laurila T, Lohila A, Loustau D, Matteucci G, Meyers T, Miglietta F, Ourcival J M, Pumpanen J, Rambal S, Rotenberg E, Sanz M, Tenhunen J, Seufert G, Vaccari F, Vesala T, Yakir D, Valentini R. On the separation of net ecosystem exchange into assimilation and ecosystem respiration: review and improved algorithm. Global Change Biology, 2005, 11(9): 1424-1439. |
| [117] | Lasslop G, Reichstein M, Papale D, Richardson A D, Arneth A, Barr A, Stoy P, Wohlfahrt G. Separation of net ecosystem exchange into assimilation and respiration using a light response curve approach: critical issues and global evaluation. Global Change Biology, 2010, 16(1): 187-208. |
| [118] | Richardson A D, Braswell B H, Hollinger D Y, Burman P, Davidson E A, Evans R S, Flanagan L B, Munger J W, Savage K, Urbanski S P, Wofsy S C. Comparing simple respiration models for eddy flux and dynamic chamber data. Agricultural and Forest Meteorology, 2006, 141(2/4): 219-234. |
| [119] | 叶子飘. 光合作用对光和 CO2响应模型的研究进展. 植物生态学报, 2010, 34(6): 727-740. |
| [120] | Papale D, Reichstein M, Aubinet M, Canfora E, Bernhofer C, Kutsch W, Longdoz B, Rambal S, Valentini R, Vesala T, Yakir D. Towards a standardized processing of Net Ecosystem Exchange measured with eddy covariance technique: algorithms and uncertainty estimation. Biogeosciences, 2006, 3: 571-583. |
| [121] | Elbers J A, Jacobs C M J, Kruijt B, Jans W W P, Moors E J. Assessing the uncertainty of estimated annual totals of net ecosystem productivity: A practical approach applied to a mid latitude temperate pine forest. Agricultural and Forest Meteorology, 2011, 151(12): 1823-1830. |
| [122] | 刘敏, 何洪林, 于贵瑞, 孙晓敏, 朱旭东, 张黎, 赵新全, 王辉民, 石培礼, 韩士杰. 数据处理方法不确定性对CO2通量组分估算的影响. 应用生态学报, 2010, 21(9): 2389-2396. |
| [123] | Twine T E, Kustas W P, Norman J M, Cook D R, Houser P R, Meyers T P, Prueger J H, Starks P J, Wesely M L. Correcting eddy-covariance flux underestimates over a grassland. Agricultural and Forest Meteorology, 2000, 103(3): 279-300. |
| [124] | Barr A G, Morgenstern K, Black T A, McCaughey J H, Nesic Z. Surface energy balance closure by the eddy-covariance method above three boreal forest stands and implications for the measurement of the CO2 flux. Agricultural and Forest Meteorology, 2006, 140(1/4): 322-337. |
| [125] | 冯宗炜, 王效科, 吴刚. 中国森林生态系统的生物量和生产力. 北京: 科学出版社, 1999. |
| [126] | 于贵瑞, 王秋凤, 刘迎春, 刘颖慧. 区域尺度陆地生态系统固碳速率和增汇潜力概念框架及其定量认证科学基础. 地理科学进展, 2011, 30(7): 771-787. |
| [127] | Goulden M L, McMillan A M S, Winston G C, Rocha A V, Manies K L, Harden J W, Bond-Lamberty B P. Patterns of NPP, GPP, respiration, and NEP during boreal forest succession. Global Change Biology, 2011, 17(2): 855-871. |
| [128] | Curtis P S, Hanson P J, Bolstad P, Barford C, Randolph J C, Schmid H P, Wilson K B. Biometric and eddy-covariance based estimates of annual carbon storage in five eastern North American deciduous forests. Agricultural and Forest Meteorology, 2002, 113(1/4): 3-19. |
| [129] | Piao S L, Fang J Y, Ciais P, Peylin P, Huang Y, Sitch S, Wang T. The carbon balance of terrestrial ecosystems in China. Nature, 2009, 458(7241): 1009-1013. |
| [130] | Yu G R, Zhu X J, Fu Y L, He H L, Wang Q F, Wen X F, Li X R, Zhang L M, Zhang L, Su W, Li S G, Sun X M, Zhang X P, Zhang J H, Yan J H, Wang H M, Zhou G S, Jia B R, Xiang W H, Li Y N, Zhao L, Wang Y F, Shi P L, Chen S P, Xin X P, Zhao F H, WangY Y, Tong C L. Spatial patterns and climate drivers of carbon fluxes in terrestrial ecosystems of China. Global Change Biology, 2013, 19(3): 798-810. |
| [131] | Wang M, Guan D X, Han S J, Wu J L. Comparison of eddy covariance and chamber-based methods for measuring CO2 flux in a temperate mixed forest. Tree Physiology, 2010, 30(1): 149-163. |
| [132] | Tan Z H, Zhang Y P, Yu G R, Sha L Q, Tang J W, Deng X B, Song Q H. Carbon balance of a primary tropical seasonal rain forest. Journal of Geophysical Research, 2010, 115(D4): D00H26, doi: 10.1029/2009JD012913. |
| [133] | Tan Z H, Zhang Y P, Schaefer D, Yu G R, Liang N S, Song Q H. An old-growth subtropical Asian evergreen forest as a large carbon sink. Atmospheric Environment, 2011, 45(8): 1548-1554. |
| [134] | Xu B, Yang Y, Li P, Shen H, Fang J. Global patterns of ecosystem carbon flux in forests: A biometric data-based synthesis. Global Biogeochemical Cycles, 2014, 28(9): 962-973. |