 2015, Vol. 35
2015, Vol. 35文章信息
- 李忠炉, 单秀娟, 金显仕, 戴芳群, 卢伙胜
- LI Zhonglu, SHAN Xiujuan, JIN Xianshi, DAI Fangqun, LU Huosheng
- 黄海中南部黄鮟鱇生物学特征及其资源密度的年际变化
- Interannual variations in the biological characteristics, distribution and stock density of anglerfish Lophius litulon in the central and southern Yellow Sea
- 生态学报, 2015, 35(12): 4007-4015
- Acta Ecologica Sinica, 2015, 35(12): 4007-4015
- http://dx.doi.org/10.5846/stxb201310262585
-
文章历史
- 收稿日期:2013-10-26
- 修订日期:2014-11-28






2. 山东省渔业资源与生态环境重点实验室 中国水产科学研究院黄海水产研究所, 青岛 266071
2. Key Laboratory for Fishery Resources and Eco-environment, Shandong Province, Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Qingdao 266071, China
黄鮟鱇(Lophius litulon)为暖温性底层鱼类,主要分布于西北太平洋,在我国分布于黄海和东海。近年来,我国近海传统经济鱼类衰退严重,但黄鮟鱇渔获量上升明显,成为张网渔获物中仅次于带鱼、小黄鱼和银鲳的底层鱼类[1, 2],是捕捞和出口的主要水产品之一[2]。因此,其种群生物学特征和资源密度的变化受到国内外学者关注。目前,对黄东海黄鮟鱇的生长[3, 4]、繁殖和分布[5, 6]、摄食[7, 8]以及资源状况[1, 2, 9]等的报道较多,但对其种群特征的长期变化研究较少。Jin等[10]发现2000—2002年黄海黄鮟鱇的食性较1985—1987年发生了较大的转变,并认为主要由于黄海生态系统食物网结构发生了变化。对种群生物学特征长期变化的研究,是了解渔业资源变化趋势和种群动态影响因子的重要方法,能为渔业科学管理和海洋生态系统研究提供基础资料。本文根据1985年、2000年、2005年和2009年秋季共4航次黄海中南部底拖网渔业调查中黄鮟鱇的渔获量和在实验室测定的生物学数据,结合环境资料和世界粮农组织(FAO)的黄鮟鱇年产量数据,对黄海中南部黄鮟鱇生物学特征和相对资源密度的年际变化进行研究,以期为该种资源的可持续利用提供参考,为研究人类影响和环境变化双重压力下黄海生态系统的变化提供基础资料。
1 数据和方法 1.1 数据来源黄鮟鱇的生物学数据根据中国水产科学研究院黄海水产研究所“北斗”号渔业调查船在黄海进行底拖网调查时捕获的样品测定,调查时间分别为1985年、2000年、2005年和2009年秋季(9—10月),4个航次共测定样品1501尾。黄鮟鱇的渔获量统计于拖网卡片。1985年使用的底拖网网口周长为450目×17 cm,网囊网目2.0 cm,网口高度5—7 m;2000年后各年网口周长836目×20 cm,网囊网目2.4 cm,网口高度5—7 m。拖网站点如图 1所示,调查逐站拖网,每站0.5—1 h,昼夜连续进行,平均拖速3.0 kn。

|
| 图 1 黄海中南部拖网取样站点 Fig.1 Survey and sampling sites in the central and southern Yellow Sea |
温度和盐度数据采集于2009年秋季航次的海上同步测定,测定工具为Seabird-19型温盐深仪(CTD),以离海底5 m水层的海水温度和盐度代表黄鮟鱇的栖息地环境。
FAO的黄鮟鱇年产量数据主要为1985年至2009年韩国的年捕捞量[11],相应时间的黄海中南部海表温度数据取自哥伦比亚大学海洋环境数据库[12] [Jul. 8th 2010 from IRI Data Library: http:// iridl.ldeo.columbia. edu/]。
1.2 分析方法 1.2.1 种群特征体长和体重分别测量到0.1 cm和1.0 g,体长以2.0 cm为间隔计算优势体长范围和体长频率分布。性腺成熟度分为Ⅰ—Ⅵ期[13],本研究设定Ⅳ期以上(Ⅳ—Ⅵ期)为性成熟。
黄鮟鱇体长-体重关系用幂指数方程表示[3]:

式中,W为体重(g),L为体长(cm);使用协方差分析(AVCOVA)检验雌、雄体长-体重关系的差异显著性[14]。
1.2.2 相对资源密度各站位的拖网时间均标准化为1 h,并根据网口面积比对1985年调查数据进行标准化[15]。各站点黄鮟鱇相对资源密度以单位时间渔获量(kg/h)表示;各航次的相对资源密度以平均渔获量(单位时间渔获量的平均值,kg/h)和个体出现率表示;以黄鮟鱇占总渔获量的重量百分比表示其对总渔获量的贡献率。使用方差分析(One-way ANOVA)检验各年平均渔获量的差异显著性[14]。
2 结果 2.1 体长组成调查表明,黄海中南部黄鮟鱇雌、雄群体的体长优势范围均由1985年的双峰型转变为2000年及其后的单峰型,种群的体长结构简单化趋势明显,平均体长减小(表 1)。雌、雄群体间比较表明,雌性黄鮟鱇的体长范围较雄性的广,雌性平均体长较雄性平均体长大,且1985年和2005年雌、雄群体平均体长均差异显著(P<0.05)。
| 年份 Year |
性别 Sex |
体长Body length/ cm | ||
| 范围 Range |
优势范围 Dominant range |
平均值±标准差 Mean±SD |
||
| * 表示该年黄鮟鱇雌、雄平均体长差异显著(P<0.05) | ||||
| 1985* | 雌 | 17.5—62.4 | 20.0—24.0 (26.7%) 3 6.0—40.0 (43.3%) |
35.6±11.3 |
| 雄 | 17.5—51.0 | 20.0—26.0 (42.3%) 32.0—36.0 (34.6%) |
30.1±8.5 | |
| 2000 | 雌 雄 |
12.7—42.2 1 7.3—41.0 |
20.0—22.0 (68.4%) 18.0—22.0 (80.0%) |
22.0±5.5 2 1.9±5.0 |
| 2005* | 雌 雄 |
8.3—55.0 1 2.2—37.0 |
18.0—20.0 (52.4%) 16.0—20.0 (69.7%) |
21.1±7.6 1 9.0±4.2 |
| 2009 | 雌 雄 |
12.0—50.7 1 0.0—37.8 |
18.0—20.0 (48.4%) 16.0—20.0 (59.8%) |
19.7±5.1 1 9.6±5.1 |
黄鮟鱇雌、雄群体的体长-体重关系参数见表 2。协方差分析表明,同一年内,雌、雄群体体长-体重关系幂指数b无显著差异(P>0.05),但参数a差异较大,其中1985年、2005年均差异显著(P<0.05)。 t检验显示,1985年和2000年雌、雄群体的幂指数b均显著小于3(P<0.05),个体均呈负异速生长;2005和2009年b与3无显著差异(P>0.05),个体都呈等速生长。
| 年份 Year |
性别 Sex |
a (×10-2) |
b | R2 | 样品量/尾 Sample size/ indiv |
| 1985 | ♀ | 10.76 | 2.644 | 0.952 | 28 |
| ♂ | 5.181 | 2.827 | 0.974 | 51 | |
| 2000 | ♀ | 65.24 | 2.075 | 0.799 | 19 |
| ♂ | 10.56 | 2.643 | 0.938 | 20 | |
| 2005 | ♀ | 4.330 | 2.920 | 0.917 | 206 |
| ♂ | 3.776 | 2.950 | 0.892 | 313 | |
| 2009 | ♀ | 2.509 | 3.037 | 0.932 | 157 |
| ♂ | 2.194 | 3.084 | 0.947 | 249 |
除2000年样品的性比与1无显著差异外,各年份样品的性比均与1差异显著(P<0.05),雄性的数量显著多于雌性;各体长组中,小于20 cm体长组和40—50 cm体长组的性比年间变化明显,分别由1985年、2000年的性比与1无显著差异转变为2005年和2009年的雄性占优势(<20 cm体长组)和全部样品为雌性(40—50 cm体长组,表 3)。
| 年份 Year |
综合 Overall sex ratio |
体长组Body length group/ cm | ||||
| <20 | 20—30 | 30—40 | 40—50 | >50 | ||
| * 表示性比与1∶1显著差异(P<0.05),— 表示无数据,♀表示样品全部为雌性 | ||||||
| 1985 | 0.58∶1* | 0.67∶1 | 0.36∶1* | 0.50∶1 | 2.25∶1 | 2.00∶1ns |
| 2000 | 0.95∶1 | 0.56∶1 | 1.3∶1 | — | 1∶1 | — |
| 2005 | 0.66∶1* | 0.59∶1* | 0.65∶1* | 0.93∶1 | ♀ | — |
| 2009 | 0.63∶1* | 0.66∶1* | 0.64∶1* | 0.21∶1* | ♀ | — |
黄鮟鱇样品的性腺成熟度以Ⅱ期为主(图 2),1985年和2000年的样品中无性成熟个体,2005年和2009年样品中出现少数性成熟个体,部分个体处于产卵期。

|
| 图 2 秋季黄海中南部黄鮟鱇雌性个体性腺成熟度各期百分比组成 Fig.2 Proportion of sexually mature stages of female Lophius litulon in the central and southern Yellow Sea during autumn sampling |
各年黄鮟鱇相对资源密度和贡献率如表 4。2000年黄鮟鱇平均渔获量和贡献率最低,但出现频率最高,表明该年黄鮟鱇分布较广,但个体较小。2005年和2009年,黄鮟鱇相对资源密度和贡献率较1985年和2000年明显升高,其中平均渔获量较2000年显著升高(P<0.05)。
| 调查年份 Survey year |
平均渔获量 Average catch±S.D./(kg/h) |
出现频率 Occurrence frequency/% |
贡献率 Contribution rate/% |
| 1985 | 2.94±7.04 | 41.2 | 3.0 |
| 2000 | 1.33±2.03 | 61.8 | 1.2 |
| 2005 | 5.86±9.30 | 50.0 | 10.5 |
| 2009 | 5.62±8.75 | 59.6 | 9.8 |
黄鮟鱇的分布与底层海水温度、盐度关系密切(图 3),分布区水温范围7.1—13.6 ℃,密集区水温范围9.3—10.8 ℃,密集区渔获量占黄鮟鱇总产量的73.3%;分布区盐度范围31.5—33.3,密集区盐度范围32.4—33.2,密集区渔获量占黄鮟鱇总产量的81.7%。

|
| 图 3 2009年秋季黄海中南部黄鮟鱇资源密度指数与底层水温和盐度的关系 Fig.3 Relationship between relative stock density of Lophius litulon and bottom water temperature and salinity in the central and southern Yellow Sea during autumn 2009 |
同时,黄鮟鱇的分布受到水深的影响(图 4)。黄海西部,群体主要分布在50 m等深线东侧较深水域,50 m以浅水域分布较少。鱼群密集区34°—35° N,122°—123° E之间。

|
| 图 4 黄海中南部黄鮟鱇相对资源密度 (实心点代表黄鮟鱇资源量零值站点,空心圆表示非零值站点,半径越大代表相对资源密度越高) Fig.4 Relative stock density of Lophius litulon in the central and southern Yellow Sea (solid dots represent stations with no-capture of Lophius litulon; hollow circles indicate Lophius litulon capture; the radius of the hollow circles indicate relative stock density) |
黄鮟鱇种群呈向黄海北部扩展分布的趋势。从36° N以北各站位的资源密度变化情况看,1985年该海域无黄鮟鱇渔获量,2000年该海域黄鮟鱇出现率为57.9%,相对资源密度为(1.0±1.4) kg/h,2009年该海域黄鮟鱇的出现率增加到83.3%,相对资源密度上升到(6.9±9.8) kg/h,出现率和相对资源密度均明显升高。
2.7 产量与温度的关系黄海黄鮟鱇的产量情况可分为两个较明显的阶段,1999年之前表现为较剧烈的波动,1999年之后呈逐年升高趋势,1997—2001年黄鮟鱇年产量处于历史较低水平。与此相对应,黄海中南部海表温度也可分为两阶段,1994—2009年平均海表温度较1985—1994年平均海表温度升高约0.7 ℃,其中1996—2001年的海表温度处于历史较高水平(图 5)。回归分析表明,剔除1997—2001年产量和对应的海表温度,黄鮟鱇年产量(Yn)与提前2a的海表温度(SSTn-2)呈极显著正相关关系:
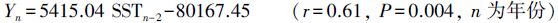

|
| 图 5 黄海中南部年均海表温度(1983—2007)与鮟鱇类年产量(1985—2009)的变化趋势(1997年—2001年鮟鱇类年产量与对应的1995年—1999年海表温度未参与拟合) Fig.5 Interannual changes in yearly mean sea surface temperature(1983—2007)and total catch of Lophiidae per year(1985—2009)in the central and southern Yellow Sea(excluding the yearly total catch of Lophiidae between 1997 and 2001 and the corresponding sea surface temperature between 1995 and 1999) |
黄鮟鱇雌性平均体长明显大于雄性,这是因为黄鮟鱇雌性的生长速度和渐近体长均大于雄性所致[3]。这种生长差异普遍存在于鮟鱇属各种中,如美洲鮟鱇(L. americanus)、欧洲鮟鱇(L.budegassa)和鮟鱇(L. piscatorius),且在高龄阶段差异更明显[16]。因此当黄鮟鱇种群年龄组成较多时,雌雄群体间平均体长差异就更显著,而当种群组成趋向低龄时,雌雄间平均体长的差异就减小。
根据Yoneda等[3]估算的生长参数,1985年秋季黄鮟鱇的优势年龄包括Ⅲ龄(2+ 龄)和Ⅴ龄(4+ 龄);到2000年仅为Ⅲ龄(2+ 龄),且2005和2009年雄性优势年龄趋向Ⅱ龄(1+ 龄),这种变化趋势和徐开达等[4]的研究结果相同,但如果按徐开达等[4]估算的生长参数,黄鮟鱇种群2000年后仅以Ⅰ龄(0+)占优势。种群结构简单化、低龄化趋势明显。目前,黄海存在着强大的捕捞压力,特别在渔业资源较为丰富的黄海中南部[17],底拖网对大个体的选择是黄鮟鱇种群结构简单化和低龄化的重要原因。
需要注意的是,Yoneda等[3]和徐开达等[4]估算的生长参数存在差异,这可能源于鉴定黄鮟鱇年龄的材料差异。Yoneda等[3]使用黄鮟鱇第9节脊柱骨作为年龄鉴定主要材料,徐开达等[4]采用的是黄鮟鱇的耳石。因此,需对黄鮟鱇脊柱骨和耳石2种硬质材料鉴定年龄的精度进一步验证。
体长-体重关系参数b反应了种群生长发育的不均匀性,与群体组成有关[18]。2000年样品量相对较少,但Froese[19]研究表明样品体长范围是影响体长-体重关系参数的主要因子。本研究中所有样品均参与拟合,因此结果具有代表性。近年来黄海中南部黄鮟鱇群体趋向低龄化,b值趋向表现低龄群体的生长。黄鮟鱇在低龄时摄食强度较高[8],摄食的饵料大小与高龄鱼无显著差异[7],因此低龄鱼体重增加更快,群体由负异速生长转变为等速生长。
张学健等[6]发现黄海南部黄鮟鱇的性比存在较明显的月变化,从11月到翌年1月以雄性为主,且全长组间的性比差异明显。本研究表明,秋季黄海中南部黄鮟鱇亦以雄性占优势,且存在体长组间差异,较小体长组以雄性为主,而大体长组以雌性为主。相同的现象也存在于美洲鮟鱇[20]、欧洲鮟鱇和鮟鱇[21]等鮟鱇属的其它种中。目前的研究认为这和鮟鱇属鱼类生长速度的性别间差异[21]、索饵或产卵洄游[20]以及繁殖策略[6]等有关。黄鮟鱇雌性个体生长快,雄性生长慢,且寿命相对短[3],因此在较小的体长组积累了较多的雄性从而占据优势,这也是欧洲鮟鱇和鮟鱇性比随体长波动的原因[21]。此外,Yoneda等[5]发现黄鮟鱇的卵母细胞从11月份开始发育,且雌性群体也从11月份开始向东海北部和外海扩张,翌年2—5月在东海至日本九州岛附近的广大海域产卵,而黄鮟鱇雄性的性成熟个体则全年均可捕获,这表明雌性性腺发育对海水温度更敏感。在海水温度的影响下,秋季黄海中南部黄鮟鱇雌性群体开始产卵洄游,雄性群体则相对滞后,这也将引起该海域黄鮟鱇种群以雄性为主。
黄鮟鱇的繁殖主要在春季[5, 6],尽管存在细微的地理差异,但由于黄、东海黄鮟鱇无明显的遗传分化[22],这种差异主要受环境因子的影响。然而本研究中,2005年和2009年样品中雌性黄鮟鱇亦有部分性成熟个体(图 2),表明秋季黄海黄鮟鱇少数个体也可产卵。Yoneda等[5]曾观察到2—7月少数黄鮟鱇个体多次产卵,美洲鮟鱇[20, 23]和黄东海的黑鮟鱇(Lophiomus setigrus)[24]也可能存在多次产卵现象,相同海域小黄鱼产卵亲体的分布范围与产卵时间均发生了变化[25, 26],因此这种现象可能是黄鮟鱇对外界压力的一种适应性反应。
3.2 相对资源密度和群体分布的变化薛莹等[27]估算2000—2002年秋季黄海中南部黄鮟鱇对饵料生物的摄食量高达到12153 t,食物重量比最高的2种饵料生物分别是鳀(Engraulis japonicus,13.0%)和脊腹褐虾(Crangon affinis,12.8%)。通过这2种饵料与黄鮟鱇形成强烈食物竞争关系的主要有带鱼(Trichiurus lepturus)、高眼鲽(Cleisthenes herzensteini)和细纹狮子鱼(Liparis tanakae)等[28]。近年来,黄东海的带鱼和高眼鲽低龄化趋势明显,资源量显著降低[29, 30],减少了饵料生物的消耗,为黄鮟鱇提供了稳定的饵料基础,是近年来黄鮟鱇相对密度显著升高的重要保证。
本研究发现鮟鱇鱼类的产量与提前2a的海表温度之间有极显著相关关系(P<0.01),温度解释了约61%的产量波动,这可能是因为黄鮟鱇的产量主要来自2龄及以上的个体,而海表温度主要影响黄鮟鱇生活史的初期,因此产量与提前2a的海表温度显著相关。FAO的资料表明,1997—2001年黄鮟鱇的产量较低,与本研究中2000年黄鮟鱇相对资源密度较低相对应(表 4),这与1996—2001年的海表温度处于历史较高水平可能有直接关系,已有研究显示1997—1999年海表温度升高造成韩国主要渔业的明显波动[31],且会降低长江口以北海区中上层鱼类资源的平均网产量[32]。
黄鮟鱇分布区海水温度较低、盐度较高且范围较窄(图 3),秋季时主要分布区的水深超过50 m,这和林龙山等[1]的研究结果相同。东海北部和日本沿岸只有当秋末水温下降后黄鮟鱇才逐渐出现[1, 5]。近年来,黄鮟鱇分布区向黄海北部扩散可能是种群对秋季黄海中南部水温升高的一种适应,模拟研究表明温度升高能促使生物向高纬度地区移动[33, 34],而从1983年到2009年,黄海中南部年均海表温度约升高了0.7 ℃(图 5)。
致谢: 感谢中国水产科学研究院黄海水产研究所资源室科研人员及“北斗”号全体船员在室内实验和野外调查取样过程中给予的帮助。
| [1] | 林龙山, 郑元甲. 东海区黄鮟鱇资源状况的初步探讨. 海洋渔业, 2004, 26(3): 179-183. |
| [2] | 程家骅, 张学健. 鮟鱇属鱼类的渔业生物学与渔业的研究概况. 中国水产科学, 2010, 17(1): 161-167. |
| [3] | Yoneda M, Tokimura M, Fujita H, Takeshita N, Takeshita K, Matsuyama M, Matsuura S. Age and growth of Anglerfish Lophius litulon in the East China Sea and the Yellow Sea. Fisheries Science, 1997, 63(6): 887-892. |
| [4] | 徐开达, 贺舟挺, 李鹏飞, 薛利建, 朱文斌. 东海北部、黄海南部黄鮟鱇的年龄和生长. 渔业科学进展, 2010, 31(6): 9-14. |
| [5] | Yoneda M, Tokimura M, Fujita H, Takeshita N, Takeshita K, Matsuyama M, Matsuura S. Reproductive cycle, fecundity, and seasonal distribution of the anglerfish Lophius litulon in the East China and Yellow Seas. Fishery Bulletin Publisher, 2001, 99(2): 356-370. |
| [6] | 张学健, 程家骅, 沈伟, 刘尊雷, 袁兴伟. 黄鮟鱇繁殖生物学研究. 中国水产科学, 2011, 18(2): 290-298. |
| [7] | 薛莹, 徐宾铎, 高天翔, 徐浩, 林龙山. 北黄海秋季黄鮟鱇摄食习性的初步研究. 中国海洋大学学报, 2010, 40(9): 39-44. |
| [8] | 张学健, 程家骅, 沈伟, 刘尊雷. 黄海南部黄鮟鱇摄食生态. 生态学报, 2010, 30(12): 3117-3125. |
| [9] | 徐开达, 贺舟挺, 朱文斌, 李鹏飞, 章守军. 东海北部和黄海南部黄鮟鱇的数量分布及其群体结构特征. 大连水产学院学报, 2010, 25(5): 465-470. |
| [10] | Jin X, Zhang B, Xue Y. The response of the diets of four carnivorous fishes to variations in the Yellow Sea ecosystem. Deep Sea Research Part II: Topical Studies in Oceanography, 2010, 57(11-12): 996-1000. |
| [11] | FAO, Fisheries Global Information System (FIGIS), in Updated 13 July 2010. [Cite 11 August 2010]. http://www.fao.org/fishery/figis/en, FAO Fisheries and Aquaculture Department, 2010: Rome. |
| [12] | Smith T M, Reynolds R W, Peterson T C, Lawrimore J. Improvements to NOAA's historical merged land-ocean surface temperature analysis (1880-2006). Journal of Climate, 2008, 21(10): 2283-2296. |
| [13] | 黄海水产研究所. 海洋水产资源调查手册 (第二版). 上海: 上海科学技术出版社, 1981: 39-39. |
| [14] | Zar J H. Biostatistical Analysis. 4th ed. Englewood Cliffs, New Jersey: Prentice-Hall, 1999: 929-929. |
| [15] | Xu B D, Jin X S. Variations in fish community structure during winter in the southern Yellow Sea over the period 1985-2002. Fisheries Research, 2005, 71(1): 79-91. |
| [16] | Fariña A C, Azevedo M, Landa J, Duarte R, Sampedro P, Costas G, Torres M A, Cañás L. Lophius in the world: a synthesis on the common features and life strategies. ICES Journal of Marine Science, 2008, 65(7): 1272-1280. |
| [17] | 徐宾铎, 金显仕, 梁振林. 秋季黄海底层鱼类群落结构的变化. 中国水产科学, 2003, 10(2): 148-154. |
| [18] | 李忠炉, 金显仕, 单秀娟, 戴芳群. 小黄鱼体长-体质量关系和肥满度的年际变化. 中国水产科学, 2011, 18(3): 602-610. |
| [19] | Froese R. Cube law, condition factor and weight-length relationship: history, meta-analysis and recommendations. Journal of Applied Ichthyology, 2006, 22(4): 241-253. |
| [20] | Richards R A, Nitschke P C, Sosebee K A. Population biology of monkfish Lophius americanus. ICES Journal of Marine Science, 2008, 65(7): 1291-1305. |
| [21] | Duarte R, Azevedo M, Landa J, Pereda P. Reproduction of anglerfish (Lophius budegassa Spainola and Lophius piscatorius linnaeus) from the Atlantic Iberian coast. Fisheries Research, 2001, 51(2/3): 349-361. |
| [22] | 姬广磊, 高天翔, 柳本卓. 黄海和日本海黄鮟鱇的形态和同工酶差异. 海洋水产研究, 2007, 28(3): 73-79. |
| [23] | Johnson A K, Richards R A, Cullen D W, Sutherland S J. Growth, reproduction, and feeding of large monksh, Lophius americanus. ICES Journal Marine Science, 2008, 65(7): 1306-1315. |
| [24] | Yoneda M, Tokimura M, Fujita H, Takeshita N, Takeshita K, Matsuyama M, Matsuura S. Ovarian structure and batch fecundity in Lophiomus setigerus. Journal of Fish Biology, 1998, 52(1): 94-106. |
| [25] | 林龙山, 姜亚洲, 严利平, 高天翔,王金辉. 黄海南部和东海小黄鱼产卵亲体分布特征与繁殖力的研究. 上海海洋大学学报, 2009, 18(4): 453-459. |
| [26] | 林龙山, 程家骅, 姜亚洲, 袁兴伟, 李建生, 高天翔. 黄海南部和东海小黄鱼(Larimichthys polyactis)产卵场分布及其环境特征. 生态学报, 2008, 28(8): 3485-3494. |
| [27] | 薛莹, 金显仕, 赵宪勇, 梁振林, 李显森. 秋季黄海中南部鱼类群落对饵料生物的摄食量. 中国海洋大学学报, 2007, 37(1): 75-82. |
| [28] | 张波, 唐启升, 金显仕, 薛莹. 东海和黄海主要鱼类的食物竞争. 动物学报, 2005, 51(4): 616-623. |
| [29] | 凌建忠, 李圣法, 严利平, 程家骅. 基于Beverton-Holt模型的东海带鱼资源利用与管理. 应用生态学报, 2008, 19(1): 178-182. |
| [30] | 张波. 黄海中部高眼鲽的摄食及随体长的变化. 应用生态学报, 2007, 18(8): 1849-1854. |
| [31] | Kim S, Zhang C I, Kim J Y, Oh J H, Kang S, Lee J B. Climate variability and its effects on major fisheries in Korea. Ocean Science Journal, 2007, 42(3): 179-192. |
| [32] | 张甲珅, 苏奋振, 杜云艳. 东海区中上层鱼类资源与海表温度关系. 资源科学, 2004, 26(5): 147-152. |
| [33] | Pörtner H O, Berdal B, Blust R, Brix O, Colosimo A, Wachter B D, Giuliani A, Johansen T, Fischer T, Knust R, Lannig G, Naevdal G, Nedenes A, Nyhammer G, Sartoris F J, Serendero I, Sirabella P, Thorkildsen S, Zakhartsev M. Climate induced temperature effects on growth performance, fecundity and recruitment in marine fish: developing a hypothesis for cause and effect relationships in Atlantic cod (Gadus morhua) and common eelpout (Zoarces viviparus). Continental Shelf Research, 2001, 21(18-19): 1975-1997. |
| [34] | Cheung W W L, Lam V W Y, Sarmiento J L, Kearney K, Waston R, Pauly D. Projecting global marine biodiversity impacts under climate change scenarios. Fish and Fisheries, 2009, 10(3): 235-251. |