 2015, Vol. 35
2015, Vol. 35文章信息
- 王静, 王克林, 张明阳, 章春华
- WANG Jing, WANG Kelin, ZHANG Mingyang, ZHANG Chunhua
- 南方丘陵山地带植被净第一性生产力时空动态特征
- Tempo-spatial variations of net primary productivity in hilly terrain of southern China
- 生态学报, 2015, 35(11): 3722-3732
- Acta Ecologica Sinica, 2015, 35(11): 3722-3732
- http://dx.doi.org/10.5846/stxb201308162091
-
文章历史
- 收稿日期:2013-08-16
- 修订日期:2014-02-25






2. 中国科学院环江喀斯特生态系统观测研究站, 环江 547100;
3. 中国科学院大学, 北京 100049
2. Huanjiang Observation and Research Station for Karst Ecosystem, Chinese Academy of Science, Huanjiang 547100, China;
3. University of Chinese Academy of Sciences, Beijng 100049, China
植被作为陆地生物圈的重要组成部分,在调节全球碳平衡具有不可替代的作用。植被净第一性生产力(NPP)指绿色植物在单位面积、单位时间内所积累的有机物数量,由光合作用所产生的有机质总量扣除自养呼吸后的剩余成分,是真正用于植物生长和生殖的光合产物量或有机碳量。NPP不仅反映陆地生态系统的光合作用和呼吸作用的能力,表征陆地生态系统的生产能力状况,而且与大气圈-生物圈的碳循环具有密切关系,是判定生态系统碳源/汇和调节生态过程的主要因子,在全球变化及碳平衡中扮演着重要的角色[1, 2, 3, 4, 5]。区域尺度上估算陆地生态系统NPP已成为全球环境变化研究热点之一。
通常估算陆地NPP的模型有3种,即统计模型、参数模型和过程模型[6, 7, 8, 9, 10]。统计模型亦称气候相关模型,最为经典的有Miami模型和Thornthwaite Memorial模型,模型考虑因子简单,计算精度不高,缺乏广泛的适用性[11]。参数模型是植物生态生理学和统计相关方法相结合的产物(代表有Chikugo模型),模型所估算的NPP实际是潜在或最大NPP,与实际差别较大。过程模型有着完整的理论框架,从机理上对植物的生物物理过程以及影响因子进行分析和模拟[12],以TEM模型、BIOME-BGC模型、CERTURY模型为代表。但过程模型涉及参数多,精度难控制。遥感数据具有时间序列长和覆盖面广的特点,利用遥感数据、GIS技术和模型模拟NPP已成为一种全新手段[13, 14, 15, 16, 17]。Carnegie-Ames-Stan-ford Approach(CASA)[18]是基于光能利用率(LUE)的过程模型,是从植被的生理过程出发而建立的植被NPP模型,在全球以及区域生产力的估算中有着广泛的应用[18, 19, 20, 21, 22, 23, 24, 25]。植被吸收的光合有效辐射(APAR)和光能利用率(ε)的准确估算是利用CASA模型模拟NPP的基础。以往模型在估算光合有效辐射吸收比例(FAPAR)过程中忽略了植被覆盖分类精度对NPP估算结果的影响。朱文泉等[16]将植被覆盖分类引进模型,考虑了植被覆盖分类精度对NPP估算的影响,从而获得不同植被类型的比值植被指数最大值并实现FPAR估算。早期Potter等和Field等认为在理想状态下植被存在最大光能利用率(ε*),即全球植被的月最高光利用率ε*应为0.389 gC/MJ[2, 4, 18]。事实上,因光能利用率ε受气温、水分、土壤、个体发育、基因型差异和植物维持与生长的不同能量分配等因素的影响,不同植被的光能利用率ε存在很大的差异[15]。因此,要基于修正过的APAR准确模拟植被NPP,光能利用率ε的有效估算也尤为重要[26, 27]。
南方丘陵山地带作为我国 “两屏三带” 生态安全屏障骨架的重要组成部分,是长江流域与珠江流域的分水岭及源头区,对长江流域与珠江流域的主体功能的发挥有至关重要的作用。优越的气候环境,使其林地面积占研究区面积的74%,在我国碳平衡中扮演重要角色。但很少有人对南方丘陵山地带的植被第一性生产力进行估算研究,尤其是自1999年以来退耕还林(草)、人工造林等植被生态工程的实施,喀斯特石漠化综合治理施行的生态移民等工程,土地覆盖发生了明显变化,随之区域植被NPP空间分布及其年际变化状况急需通过一定的方法进行明确。因此本研究基于CASA模型,通过修正模型参数,对该区域2000—2010年的植被NPP进行模拟,并进一步分析植被生产力状况的时空变化及驱动机制,以期为该区域的生态建设工程效益评估和生态保护提供科学参考和基础数据。
1 研究区域概况南方丘陵山地带涉及江西、湖南、广东、广西、贵州和云南6省市自治区,含114个县(市),面积为28.8万平方公里。地域辽阔,东西跨15个经度(102°45′—117°08′E),南北跨6个纬度(22°45′—27°14′N)。地势大致西高东低,北高南低,高程在-25—3040 m之间。气候类型为亚热带季风气候,植被类型主要有亚热带常绿阔叶林、亚热带针叶林、混交林、温带阔叶/针叶林、亚热带灌丛、亚热带草地等(图 1)。

|
| 图1 研究区植被分类及高程图 Fig.1 Vegetation types and elevation in the study area |
本研究的遥感数据来自美国国家航空航天局(NASA)的数据信息服务中心(DISC)提供的2000—2010年每月MODIS遥感影像中的NDVI数据(http://modis.gsfc.nasa.gov/),其空间分辨率为250 m×250 m,时间分辨率为16d,共253幅图像。图像预处理包括大气校正、辐射校正、格式和投影转换,以及利用最大合成法(MVC)获得逐月数据集。
2.1.2 气象数据2000年1月—2010年12月的气象数据来源于中国气象科学数据共享服务网(http://cdc.cma.gov.cn),包括71个站台的月均气温和月总降水量,27个站台的月总太阳辐射。以分辨率为90 m的数字高程模型(DEM)(图 1)为基础,采用薄盘光滑样条法[28],将气象数据插值为250 m的经纬网投影的栅格数据。
2.1.3 植被数据该数据来源于Landsat TM全国30 m分辨率的十年生态系统分类影像数据,经过地面矫正和辐射校正后,采用面向对象分类方法将土地利用分为9类:亚热带常绿阔叶林、亚热带针叶林、混交林、温带阔叶/针叶林、亚热带灌丛、亚热带草地、耕地、水域及其他。分类精度为85%。在此基础上进行了重采样,提取了250 m分辨率的植被数据。
2.2 研究方法 2.2.1 模型介绍(1) 模型模拟
CASA模型是基于遥感、气象、植被及土壤类型数据共同驱动的光能利用率模型[2, 18, 25],植被NPP可以由植物吸收的光合辐射(APAR)和实际光能利用率(ε)两个因子确定。APAR取决于总辐射(SOL)和植被本身对入射光合有效辐射的吸收比例(FPAR)。通过引入植被覆盖分类,计算不同植被覆盖类型的比值植被指数最大值,实现对FPAR的估算;现实条件下的光能利用率受温度(Tε1和Tε2)及水分(Wε)胁迫系数影响,利用气象数据,结合区域蒸散模型估算温度胁迫因子和水分胁迫因子,其计算如下:
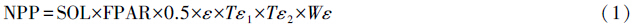
式中,FPAR在一定范围内与归一化植被指数(NDVI) 之间存在着线性关系[29, 30, 31],CASA模型即用NDVI表示FPAR对NPP进行估算(公式2—5),常数0.5表示植被所能利用的太阳有效辐射占太阳总辐射的比例,Tε1表示在低温和高温时植物体内在的生化作用对光合的限制而降低净第一性生产力(公式6),Tε2表示环境温度从最适宜温度向高温和低温变化时植物光能利用率逐渐变小的趋势(公式7),Wε反映了植物所能利用的有效水分条件对光能利用率的影响,随着环境有效水分的增加,Wε逐渐增大,它的取值范围为0.5(在极端干旱条件下)到1(非常湿润条件)(公式8):

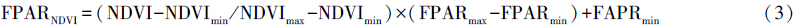
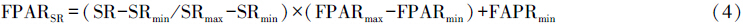
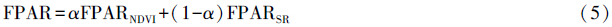



式中,SR代表比值植被指数,FPARSR和FPARNDVI分别代表根据SR和NDVI估算得到的FPAR,FPARmin和FPARmax的取值与植被类型无关,FPARmin=0.001,FPARmax=0.95,NDVImin和NDVImax分别对应某植被类型NDVI的5%和95%处的百分位数。α为两种方法间的调整系数在本研究中统一定为0.5。Topt为植物生长的最适温度,定义为某一区域一年内NDVI值达到最高时的当月平均气温。E为区域实际蒸散量(mm),可据周广胜和张新时[32, 33]建立的区域实际蒸散模型求取;Ep为区域潜在蒸散量(mm),可据Boucher提出的互补关系求取,具体参照Thornth-waite植被-气候关系模型的计算方法[34]。
(2) 最大光能利用率的确定
由于全球最大光能利用率ε*的取值对NPP的估算结果影响很大,不同学者在不同的模型中取值均不同,其范围从0.09到2.16 gC/MJ[35, 36, 37]。ε*的确定不仅受植被类型的影响,而且受空间分辨率和植被覆盖均匀程度的影响。
本研究区地域辽阔并存在典型的喀斯特地貌,特殊的碳酸盐岩基质造就了区别于亚热带典型顶极群落的常绿阔叶林的常绿落叶阔叶混交林。为使研究结果更为精确,本研究结合近年来学者对模型最大光能利用率的模拟改进,采用不同的最大光能利用率[15, 26, 27](表 1)。
| 植被类型 Vegetation type | 最大光能利用率/(gC/MJ) Maximum Light Use Efficiency (LUE)ε* | 适用省份 Zone of application | 文献 References |
| 亚热带常绿阔叶林 | 0.635 | 云南、贵州、广西、湖南 | [27] |
| Subtropical broadleaf evergreen forest | 1.250 | 广东、江西 | [15] |
| 亚热带针叶林 | 0.476 | 云南、贵州、广西、湖南 | [27] |
| Subtropical needleleaf forst | 1.250 | 广东 | [15] |
| 0.389 | 江西 | [26] | |
| 混交林mixed forest | 0.565 | 云南、贵州、广西、湖南 | [27] |
| 1.250 | 广东 | [15] | |
| 0.768 | 江西 | [26] | |
| 落叶阔叶林/针叶林 | 0.590 | 云南、贵州、广西、湖南 | [27] |
| Broadleaved/needleleaved | 1.250 | 广东 | [15] |
| deciduous forest | 0.692 / 0.485 | 江西 | [26] |
| 亚热带灌丛Subtropical shrub | 0.500 | 云南、贵州、广西、湖南 | [27] |
| 1.250 | 广东 | [15] | |
| 0.429 | 江西 | [26] | |
| 草地Grassland | 0.150 | 云南、贵州、广西、湖南 | [27] |
| 1.250 | 广东 | [15] | |
| 0.542 | 江西 | [26] | |
| 农业植被 | 0.500 | 云南、贵州、广西、湖南 | [27] |
| Agriculture vegetation | 0.542 | 广东、江西 | [26] |
| 无植被Non-vegetation | 0.001 | 云南、贵州、广西、湖南、广东、江西 | [27] |
为研究近十年来南方丘陵山地带植被生产力状况以及这种变化速率在空间上的差异,对CASA模型模拟出来的2000—2010年每个象元的NPP与年份进行线性回归分析,最小二乘法拟合得到的一元线性方程斜率即为趋势倾向率,并用相关系数检验法来确定变化是否显著,并设置0.05显著性水平对相关系数进行分类。
3 结果与分析 3.1 NPP空间分布由2000—2010年间年平均NPP分布图(图 2)分析可知,NPP平均为445.7 gC m-2 a-1,NPP高值区占研究区总面积的69.85%,主要为研究区东部(广东、江西)、中部(广西)及西南小部分地区(云南部分地区),而中低值区主要为研究区的北部(贵州、湖南部分地区)和西南地区(云南)。对比NPP空间分布与植被类型空间分布(图 2和图 1)可知,NPP空间分布与植被类型空间分布密切相关。NPP高值区主要分布在研究区亚热带常绿阔叶林、常绿针叶林及混交林覆盖地区(>410 gC m-2 a-1),而低值主要分布在草地及耕地覆盖区。

|
| 图2 2000—2010年均净初级生产力 (NPP) 空间分布 Fig.2 Spatial distribution of mean net primary productivity (NPP) during the period of 2000—2010 |
研究区2000—2010年NPP总量平均值为117.0 TgC/a,其中2009年NPP总量最大(126.4 TgC/a),2006年最小(仅为108 TgC/a)。10年来研究区年NPP上升趋势不显著(P=0.39),平均每年仅增加0.255 TgC(图 3)。

|
| 图3 2000—2010净初级生产力总量 (NPP) 及降水量、温度时间动态 Fig.3 Temporal variations in NPP,temperature,and precipitation in 2000—2010 |
研究区74.4%的区域年平均NPP呈增加趋势,其中8.1%的地区增加趋势显著(P<0.05),主要分布在研究地区西部云南省的丘北县、硯山县及贵州省境内的黔西南州和黔南州,研究区中部,北部湖南省的永州市和郴州市和东部广东省的大埔县和丰顺县(图 4)。这些地区均是由耕地及裸岩裸地构成的其他土地利用类型转为林地的主要地区,其中耕地转为林地面积有311.7 km2。NPP显著降低的区域占2.1%,主要分布在广西与云南交界处的广南县和西林县,贵州的黔东南州,广西的都安,江西的寻乌县及广东与江西接壤的连平、和平县等。

|
| 图4 2000—2010年净初级生产力总量 (NPP) 变化趋势的空间分布 Fig.4 Spatial distribution of trends in annual total net primary productivity (NPP) from 2000 to 2010 1: 显著减小 Decreased severely; 2: 减小,但不显著Decreased slightly; 3: 增加,但不显著Increased slightly; 4: 显著增加Increased greatly |
植被NPP变化不仅受气候变化影响,还受人类活动制约。某种程度上自然环境条件对土地覆盖变化起主导作用,但社会、经济、政策人为因素对土地覆盖的时空变化有重要影响。
3.3.1 温度、降水量对NPP的影响研究区2000—2010年温度上升趋势不显著(P=0.129),出现先上升后下降而后又上升的变化趋势(图 3b),除2004和2006年之外,温度与NPP的年总量呈显著正相关(R2=0.524,P=0.01)。研究区年降水量充沛(>1000 mm),温度对植物生长有着直接的效应:升温可以延长生长季,提高光合作用和水分利用效率,促进植物生长发育[38, 39],从而促进了植被净初级生产力的增加,温度对植被生长的影响在本研究区表现为正效应。
有研究表明NPP对水分敏感[40],方精云等[19, 22]在估算1982—1999年中国净初级生产力的研究结果表明NPP值与降水量有显著的正相关性。而本文图 3c显示NPP值与降水量相关性并不显著,且在NPP高值的2003、2004及2009年份,其年降水量(<1300 mm)均低于其他年份,原因可能是:(1)以往所得植被生产力与降水之间存在显著正相关的研究[19, 41, 42],其年降水量均在1000 mm以下,而本研究各年的年降水量均在1000 mm以上;可能因为本研究区降水量相对充沛,从而水分条件不是植被生长的限制因子,所以降水量对植被生产力的重要性并未体现出来[43, 44]。(2)本研究结果显示年降水量在1000—1300 mm范围内,NPP出现高值;而年降水量超过1300 mm,NPP总量均有不同程度的降低,可能是降水量过多造成雨天的增加,减弱了植被的光合作用,从而影响了NPP值,2002、2006、2008、2010年的降水量大而生产力不高一定程度能表征。
3.3.2 土地覆被变化对NPP的影响人类活动是土地覆被变化不可忽视的驱动因素之一,土地覆被变化直接影响着植被的生产力。常年以来的植树造林、封山育林及自1999年启动的退耕还林(草)植被建设工程,对土地覆被变化有着重要的影响。植被生态工程建设、劳动力的转移及经济水平的发展等人类活动是影响研究区植被覆盖变化的主要因素。图 5显示了2000—2010年研究区带不同植被类型的面积变化。结果显示,11年间研究区主要植被类型发生了明显的变化,尤以耕地变化剧烈。城镇用地面积有显著增加,随着城市化进程的加速,城镇建设急剧扩张,大量侵占其他生态类型用地,研究时段内城镇用地增加了818.0 km2。通过对NPP变化趋势图与土地利用类型转化图叠加分析得出,东部NPP显著下降区域土地覆被主要是由耕地和林地转为城镇用地。森林植被中除混交林有稍微下降,其余森林植被都有增加,其中以常绿阔叶林的面积增加最明显。研究区常绿阔叶林增加区主要分布在云南、广西、湖南及广东境内,尤以广西、广东分布集中。有研究表明广东、广西、海南是我国人工林桉树的主要基地,人工林桉树种植的迅速发展对研究区常绿阔叶林面积的增加有重大的贡献。森林和草地面积的增加及耕地面积的减少说明了植树造林、退耕还林(草)效果显著。

|
| 图5 2000—2010年不同植被类型面积变化 Fig.5 Area variation of different vegetation types during the period of 2000—2010 |
喀斯特作为研究区的典型地貌,面积有1.02×105 km2,占研究区总面积35.8%。喀斯特地区典型的生态脆弱出现了以石漠化为特征的土地退化。20世纪90年底以来,随着国家八七扶贫计划及西部大开发战略的实施,进行了退耕还林(草)、封山育林、生态移民等重大建设工程,其对石漠化区的植被恢复起到了重要作用。图 6显示了研究区喀斯特地貌的空间分布区及其所在区域11年来NPP的变化速率。喀斯特地貌集中分布在研究区广西、云南、贵州和湖南境内,广东西部也有少量分布。在研究时段内喀斯特区域NPP有显著增加趋势,表明西南石漠化综合治理工程的生态效益正在呈现。

|
| 图6 2000—2010年NPP变化速率及典型喀斯特地貌空间分布 Fig.6 The variation trend of annual NPP in 2000—2010 and the spatial distribution of typical karst areas |
不同植被类型因自身生理条件对光能的吸收和转化率不同,加上生态环境因素的差异,从而NPP存在一定的差异[24]。研究区平均NPP最高为混交林(501.0 gC m-2 a-1,集中分布于云贵高原和桂西北喀斯特地区,其中常绿落叶阔叶混交林为喀斯特区的顶极植被类型),其次为常绿阔叶林(486.7 gC m-2 a-1)和常绿针叶林(470.7 gC m-2 a-1),最低为耕地(396.7 gC m-2 a-1)和草地(390.7 gC m-2 a-1)(表 2)。从不同植被类型的面积比例来看,常绿阔叶林和常绿针叶林的分布范围广,是研究区面积最大的两类植被类型,二者占区域植被面积的比例分别为26.3%和28.7%;落叶针叶林的面积最小,集中分布于广东西北部。
| 植被类型 Vegetation type | 面积 Area/ ×104 km2 | 平均NPP Average NPP/ (gC m-2 a-1) | 标准差SD/ (gC m-2 a-1) | 总NPP Total NPP / (TgC) |
| NPP: 净第一性生产力Net Primary Productivity; SD: 标准差Standard Deviation | ||||
| 常绿阔叶林Broadleaved evergreen vegetation | 7.50 | 486.7 | 29.5 | 36.53 |
| 落叶阔叶林Broadleaved deciduous vegetation | 0.45 | 411.2 | 16.5 | 1.87 |
| 混交林 Mixed forest | 0.90 | 501.0 | 35.6 | 4.55 |
| 常绿针叶林 Needle-leaved evergreen vegetation | 8.26 | 470.7 | 28.2 | 38.88 |
| 落叶针叶林 Needle-leaved deciduous vegetation | 0.0008 | 463.4 | 30.8 | 0.039 |
| 亚热带灌丛 Subtropical shrub | 3.91 | 412.4 | 17.0 | 16.11 |
| 草地 Grassland | 0.50 | 390.7 | 15.5 | 1.87 |
| 农业植被Agriculture vegetation | 6.37 | 396.7 | 19.8 | 25.29 |
| 合计Total | 27.89 | 125.14 | ||
年际变化中不同植被类型的NPP变化有一定的相似性(图 7),2000—2004年大幅度增长,2005年、2006年急剧下降并分别达最低值,而后3年呈增长趋势,2010年又出现下降。除落叶针叶林呈下降趋势外,常绿阔叶林、混交林、常绿针叶林、落叶阔叶林、亚热带灌丛、草地、农业植被整体均呈增长趋势,其中草地和耕地的增长趋势最为明显(分别达2.65 gC m-2 a-1和2.61 gC m-2 a-1),混交林增加幅度最小(1.07 gC m-2 a-1)。有研究表明落叶针叶林对全球变暖的响应可能更为敏感[45],全球变暖使得落叶针叶林北移,且向常绿落叶林转变,本研究落叶针叶林的下降在一定程度上是对全球变暖的响应。

|
| 图7 2000—2010年各植被类型NPP逐年变化 Fig.7 Yearly NPP changes of various vegetation types from 2000 to 2010 |
对比前期相关学者对其它地区的NPP模型模拟结果,本研究结果比较可靠准确。董丹等[26]利用CASA模型对1999—2003年西南喀斯特植被的NPP总量估算得到年均NPP为402.34 gC m-2 a-1,王冰等[46]基于MODIS数据利用光能利用率模型测得贵州省植被NPP值范围在305—560 gC m-2 a-1 之间,与本文的NPP年均值整体较接近。蔡睿等[47]利用MODIS数据对广东省2000—2005年植被NPP年际变化趋势分析结果与本研究相同。模型和驱动数据的不同使得各估算值之间存在差距,这是当前NPP模拟研究中的存在的一个主要问题[42]。
近期研究结果表明自20世纪80年代以来在北半球中高纬度地区的植物生长或NPP都存在增长趋势[19]。本研究区11年的NPP呈波动增加的趋势,平均每年增加2.28 gC/m2。这种变化与气象因子有密切的关系。温度的变化趋势与年际NPP有很好的吻合度,整体呈上升趋势,平均增加值为0.035 ℃/a。年均温度的升高可以延长植被的生长季从而提高植被的生产力。本研究区NPP的季节变化与气温及地表太阳辐射的季节变化基本相同,在夏季7、8月份气温及太阳辐射达到最大值时月NPP也达年最大值;后随着太阳的南移,气温和太阳辐射逐渐减小,其NPP值开始下降至12—2月达到最小值。研究还发现,月际NPP的变化与太阳辐射最为相似,在2005—2008及2010年的6月份太阳辐射陡减,各植被类型的6月NPP变化出现明显的低谷。降水量对NPP的影响并不显著,可能是本研究区降水丰富,水分条件基本上能满足植被生长的需要,并不是植被生长的限制因子。针对南方丘陵山地带,过多的降水量可能会导致雨天增多,太阳辐射减少,减弱了植被的光合作用从而降低植被的生产力[48](图 3)。人类活动通过改变植被覆盖,从而影响NPP的时空分布差异:2000—2010年间研究区74.4%的区域NPP呈上升趋势,近年来植树造林、封山育林、退耕还林(草)、生态移民等植被建设工程所带来的生态效益正在呈现,南方地区大规模人工林桉树的种植在一定程度上促进了研究区森林面积的增加。NPP显著降低区域是由于经济的发展、城市化进程的加速使得人们对土地的需求不断增加,林地的大面积转出,耕地面积大量占用,城镇及工业用地的掠夺式开发使植被生长处于退化状态。
不同植被NPP值及变化幅度均不同:混交林>常绿阔叶林>常绿针叶林>落叶针叶林>落叶阔叶林>亚热带灌丛>耕地>草地。其与植被的光能最大利用率相关,林地的最大光能利用率普遍较高,灌丛、耕地次之,草地则最低。针阔混交林年均值最高,但其变化幅度最小,这可能是由于常绿落叶阔叶混交林作为喀斯特区域的顶极植被类型,内部最为稳定,其能量达到平衡状态。耕地的增长幅度明显与农业生产的提高有密切关系;草地对气温和降水量等气象因子的敏感性是草地变化幅度显著的原因;而落叶针叶林呈下降趋势主要是对全球变暖的响应。从不同植被NPP的季节变化来看,生长季一般为3—11月,最大值出现在7、8月,适宜的水热条件及太阳辐射使其生长季延长。
4.2 结论本文利用GIS/RS技术,基于MODIS遥感数据和气象数据,通过改进的CASA模型估算了南方丘陵山地带2000—2010年植被净初级生产力的时空动态变化,获得以下主要结论:
(1)研究区2000—2010年NPP平均值为117.0 TgC/a,逐年呈波动变化,上升趋势不明显,平均每年增加0.255 TgC。NPP的空间分布特征与植被类型具有较好的一致性。
(2)从植被类型来看,常绿针叶林和常绿阔叶林对研究区NPP总量贡献最大,其次是耕地,落叶针叶林因面积极小贡献最小;八种典型植被年NPP除落叶针叶林外,均呈增加趋势,草地增长趋势最明显(2.65 gC m-2 a-1),混交林增加幅度最小(1.07 gC m-2 a-1)。
(3) 研究区NPP的时空动态变化受气候波动和人类活动的影响显著。在降水量充沛的南方丘陵山地带,气温和太阳辐射变化趋势与NPP的变化有很好的吻合度,社会、经济、政策等人为因素对土地覆盖的时空变化有决定性的影响,导致了NPP变化的空间分布差异。
| [1] | Cramer W, Kicklighter D W, Bondeau A, Moore B, Churking B, Nemry B, Ruimy A, Schloss A L. Comparing global models of terrestrial net primary productivity (NPP): overview and key results. Global Change Biology, 1999, 5(S1): 1-15. |
| [2] | Field C B, Randerson J T, Malmström C M. Global net primary production: Combining ecology and remote sensing. Remote Sensing of Environment, 1995, 51(1): 74-88. |
| [3] | Keeling C D, Chin J F S, Whorf T P. Increased activity of northern vegetation inferred from atmospheric CO2 measurements. Nature, 1996, 382(6587): 146-149. |
| [4] | Field C B, Behrenfeld M J, Randerson J T, Falkowski P. Primary production of the biosphere: Integrating terrestrial and oceanic components. Science, 1998, 281(5374): 237-240. |
| [5] | Ruimy A, Kergoat L, Bondeau A. Comparing global models of terrestrial net primary productivity (NPP): analysis of differences in light absorption and light-use efficiency. Global Change Biology, 1999, 5(S1): 56-64. |
| [6] | Leith H. Primary production: terrestrial ecosystems. Human Ecology, 1973, 1(4): 303-332. |
| [7] | Uchijima Z, Seino H. Agroclimatic evaluation of net primary productivity of natural vegetations (1): Chikugo model for evaluating net primary productivity. Journal of Agricultural Meteorology, 1985, 40(4): 343-352. |
| [8] | Monteith J L. Solar radiation and productivity in tropical ecosystem. Journal of Applied Ecology, 1972, 9(3): 747-766. |
| [9] | Running S W, Coughlan J C. A general model of forest ecosystem processes for regional applications I. Hydrological balance, canopy gas exchange and primary production processes. Ecological Modelling, 1988, 42(2): 125-154. |
| [10] | Parton W J, Scurlock J M O, Ojima D S, Gilmanov T G, Scholes R J, Schimel D S, Kircner T, Menaut J C, Seastedt T, Garcia Moya E, Kamnalrut A, Kinyamario J I. Observations and modeling of biomass and soil organic matter dynamics for the grassland biome worldwide. Global Biogeochemical Cycles, 1993, 7(4): 785-809. |
| [11] | 陈利军, 刘高焕, 冯险峰. 遥感在植被净第一性生产力研究中的应用. 生态学杂志, 2002, 21(2): 53-57. |
| [12] | 张娜, 于贵瑞, 赵士洞, 于振良. 基于遥感和地面数据的景观尺度生态系统生产力的模拟. 应用生态学报, 2003, 14(5): 643-652. |
| [13] | 陈利军, 刘高焕, 冯险峰. 运用遥感估算中国陆地植被净第一性生产力. 植物学报, 2001, 43(11): 1191-1198. |
| [14] | 松下文经, 杨翠芬, 陈晋, 王勤学, 龟山哲, 田村正行. 广域空间尺度上植被净初级生产力的精确推算. 地理学报, 2004, 59(1): 80-87. |
| [15] | 彭少麟, 郭志华, 王伯荪. 利用GIS和RS估算广东植被光利用率. 生态学报, 2000, 20(6): 903-909. |
| [16] | 朱文泉, 潘耀忠, 张锦水. 中国陆地植被净初级生产力遥感估算. 植物生态学报, 2007, 31(3): 413-424. |
| [17] | Gao Z Q, Liu J Y. Simulation study of China's net primary production. Chinese Science Bulletin, 2008, 53(3): 434-443. |
| [18] | Potter C S, Randerson J T, Field C B, Matson P A, Vitousek P M, Mooney H A, Klooster S A. Terrestrial ecosystem production: A process model based on global satellite and surface data. Global Biogeochemical Cycles, 1993, 7(4): 811-841. |
| [19] | Fang J Y, Piao S L, Field C B, Pan Y D, Guo Q H, Zhou L M, Peng C H, Tao S. Increasing net primary production in China from 1982 to 1999. Frontiers in Ecology and the Environment, 2003, 1(6): 293-297. |
| [20] | DeFries R S, Field C B, Fung I, Collatz G J, Bounoua L. Combining satellite data and biogeochemical models to estimate global effects of human-induced land cover change on carbon emissions and primary productivity. Global Biogeochemical Cycles, 1999, 13(3): 803-815. |
| [21] | Lobell D B, Hicker J A, Asner G P, Field C B, Tucker C J, Los S O. Satellite estimates of productivity and light use efficiency in United States agriculture, 1982-98. Global Change Biology, 2002, 8(8): 722-735. |
| [22] | Piao S L, Fang J Y. Terrestrial net primary production and its spatio-temporal patterns in Qinghai-Xizang Plateau, China during 1982-1999. Journal of Natural Resources, 2002, 17(3): 373-380. |
| [23] | 朴世龙, 方精云, 郭庆华. 1982-1999年我国植被净第一性生产力及其时空变化. 北京大学学报: 自然科学版, 2001, 37(4): 563-569. |
| [24] | 柯金虎, 朴世龙, 方精云. 长江流域植被净第一性生产力及其时空格局研究. 植物生态学报, 2003, 27(6): 764-770. |
| [25] | 朴世龙, 方精云, 郭庆华. 利用CASA模型估算我国植被净第一性生产力. 植物生态学报, 2001, 25(5): 603-608. |
| [26] | 朱文泉, 陈云浩, 潘耀忠, 李京. 基于GIS和RS的中国植被光利用率估算. 武汉大学学报: 信息科学版, 2004, 29(8): 694-698, 714-714. |
| [27] | 董丹, 倪健. 利用CASA模型模拟西南喀斯特植被净第一性生产力. 生态学报, 2011, 31(7): 1855-1866. |
| [28] | 刘志红, Li L T, McVicar T R, VanNiel T G, 杨勤科, 李锐. 专用气候数据空间插值软件ANUSPLIN及其应用. 气象, 2008, 34(2): 92-100. |
| [29] | Hatfield J L, Asrar G, Kanemasu E T. Intercepted photosynthetically active radiation estimated by spectral reflectance. Remote Sensing of Environment, 1984, 14(1/3): 65-75. |
| [30] | Sellers P J. Canopy reflectance, Photosynthesis and Transpiration. International Journal of Remote Sensing, 1985, 6(8): 1335-1372. |
| [31] | Huemmrich K F, Goward S N. Spectral vegetation indexes and the remote sensing of biophysical parameters//Proceedings of international Geoscience and Remote Sensing Symposium(IGARSS). Institute of Electrial and Electronics Engineers, Houston Texas, 1992: 1017-1019. |
| [32] | 周广胜, 张新时. 自然植被净第一性生产力模型初探. 植物生态学报, 1995, 19(3): 193-200. |
| [33] | 周广胜, 张新时. 全球气候变化的中国自然植被的净第一性生产力研究. 植物生态学报, 1996, 20 (01): 11-19. |
| [34] | 张新时, 杨奠安, 倪文革. 植被的PE(可能蒸散)指标与植被-气候分类(三)几种主要方法与PEP程序介绍. 植物生态学与地植物学学报, 1993, 17(2): 97-109. |
| [35] | Ruimy A, Saugier B, Dedieu G. Methodology for the estimation of terrestrial net primary production from remotely sensed data. Journal of Geophysical Research, 1994, 99(D3): 5263-5263. |
| [36] | Paruelo J M, Epstein H E, Lauenroth W K, Bueke I C. ANPP estimates from NDVI for the central grassland region of the United States. Ecology, 1997, 78(3): 953-958. |
| [37] | McCrady R L, Jokela E J. Canopy dynamics, light interception, and radiation use efficiency of selected loblolly pine families. Forest Science, 1998, 44(1): 64-72. |
| [38] | Walker B, Steffen W, Canadell J, Ingram J. The Terrestrial Biosphere and Global Change: Implications for Natural and Managed Ecosystems. Cambridge: Cambridge University Press, 1999: 190-228. |
| [39] | 朴世龙, 方精云. 最近18年来中国植被覆盖的动态变化. 第四纪研究, 2001, 21(4): 294-302. |
| [40] | Churkina G, Running S W, Schloss A L. Comparing global models of terrestrial net primary productivity (NPP): the importance of water availability. Global Change Biology, 1999, 5(S1): 46-55. |
| [41] | 谷晓平, 黄枚, 季劲钧, 吴战平. 近20年气候变化对西南地区植被净初级生产力的影响. 自然资源学报, 2007, 22(2): 251-259. |
| [42] | 陶波, 李克让, 邵雪梅, 曹明奎. 中国陆地净初级生产力时空特征模拟. 地理学报, 2003, 58(3): 372-380. |
| [43] | 孙睿, 朱启疆. 气候变化对中国陆地植被净第一性生产力影响的初步研究. 遥感学报, 2001, 5(1): 58-61. |
| [44] | Schloss A L, Kicklighter D W, Kaduk J, Wittenberg U. Comparing global models of terrestrial net primary productivity (NPP): comparison of NPP to climate and the Normalized Difference Vegetation Index (NDVI). Global Change Biology, 1999, 5(S1): 25-34. |
| [45] | 贾丙瑞, 周广胜. 北方针叶林对气候变化相应的研究进展. 地球科学进展, 2009, 24(6): 668-674. |
| [46] | 王冰, 杨胜天, 王玉娟. 贵州省喀斯特地区植被净第一性生产力的估算. 中国岩溶, 2007, 26(2): 98-104. |
| [47] | 蔡睿, 徐瑞松, 陈彧, 苗莉, 王洁. 广东省植被NPP时空特征变化分析. 农机化研究, 2009, 31(2): 9-11, 16-16. |
| [48] | 蒙吉军, 王钧. 20世纪80年代以来西南喀斯特地区植被变化对气候变化的响应. 地理研究, 2007, 26(5): 857-865. |