 2015, Vol. 35
2015, Vol. 35文章信息
- 孙涛, 刘瑞鹏, 李兴欢, 毛子军, 黄唯, 董利利
- SUN Tao, LIU Ruipeng, LI Xinghuan, MAO Zijun, HUANG Wei, DONG Lili
- 模拟氮沉降对东北地区兴安落叶松树干呼吸的影响
- Effects of simulated nitrogen deposition on stem respiration of Larix gmelinii Rupr in Northeastern China
- 生态学报, 2015, 35(11): 3684-3691
- Acta Ecologica Sinica, 2015, 35(11): 3684-3691
- http://dx.doi.org/10.5846/stxb201308042017
-
文章历史
- 收稿日期:2013-8-4
- 网络出版日期:2014-6-12






氮元素是影响森林生态系统生产力的主要限制因子[1, 2]。但自工业革命以来,由于人类活动向大气中排放了大量的氮化物,导致这些氮化物在大气中不断累积并向陆地和水域生态系统沉降[3]。预计到2050年,全球氮沉降量将比1995年高出1倍,增加至200Tg N/a[4]。目前我国已成为继欧洲和北美之后的第三大氮沉降区,氮沉降现象在局部地区非常严重,而且伴随着人类活动的进一步加强,我国氮沉降正处于持续增加的状态[5, 6]。大气氮沉降的不断增加,将会对陆地生态系统的结构和功能产生不同程度的影响。
树木的自养呼吸可消耗自身叶片光合作用固定碳的50%以上[7],对维持森林生态系统碳平衡具有非常重要的作用。其中,树干呼吸消耗的碳占树木自养呼吸的14%—48%[8, 9, 10],其等于或有时已高于叶片呼吸释放的碳通量[11]。另外,树干呼吸也是树木生长活力及碳代谢状况的重要物候学指标之一[12, 13]。树干释放的CO2的来源和去向十分复杂,受多种因素的制约,主要包括生物因素和非生物因素,前者如树干的生长速率、液流密度、非结构性碳水化合物含量、氮含量、直径和边材厚度等[9, 12, 14, 15],后者如树干的温度、大气CO2浓度、树干内的氧气浓度、土壤湿度及养分等[11, 16, 17, 18, 19, 20, 21]。其中氮元素含量与树干呼吸密切相关,这是因为植物组织中90%的氮以蛋白质的形式存在[22],树木的生命活动大约消耗总维持呼吸的20%[23]。因此,大气氮沉降的持续增加会对树干呼吸产生重要影响,然而关于这方面的研究还很缺乏,且研究结果不一致。Maier报道施肥后火炬松的树干呼吸速率显著增加,与之伴随的是树干生长速率和氮含量都显著增加,而且发现树干生长速率和氮含量可解释树干基础呼吸季节动态的75%[14]。而别的研究结果与之相反,发现施肥后在没有引起树干生长加快的情况下,树干呼吸速率也没有显著变化[24, 25]。
落叶松(Larix gmelinii)是我国东北林区重要的建材树种,落叶松人工林已构成了东北地区主要人工林类型。目前Wang等[13]、石新立等[18]、许飞等[20]和王秀伟等[26]已对兴安落叶松树干呼吸的日变化和季节变化、种内差异、温度敏感性、与液流速度及胸径的关系进行了系统研究,但尚未开展关于氮素添加对落叶松树干呼吸影响方面的研究。我国东北地区的氮沉降量虽然低于较为严重的中南部地区,但近年来随着化肥施用量的逐渐增加等人类活动的影响,氮沉降量也在持续增长,可能会对森林生态系统碳平衡产生重要影响。我们通过对黑龙江帽儿山2011年大气年氮湿沉降进行监测,发现该地区氮湿沉降为19.16 kg N hm-2 a-1,其结果要明显高于刘世荣[27]在1992年在该区测定的氮湿沉降值(12.9 kg N ha-1)。本研究以东北典型造林树种兴安落叶松为研究对象,在兴安落叶松人工林内进行了不同水平的氮沉降实验,旨在分析氮沉降对落叶松树干呼吸通量的影响,并探讨其影响机理,为全球变化尤其在氮沉降持续增加的背景下,理解温带森林碳循环过程及其模型构建提供基础数据和科学依据。
1 研究方法 1.1 研究地概况本试验地设立在黑龙江省尚志市帽儿山林场老山试验站(45°20′N,127°34′E),地处长白山系张广才岭西北部小岭余脉,平均海拔340 m。该地区气候属于大陆性季风气候,年降雨量600—800 mm,雨量主要集中于7、8月份,年蒸发量1093.9 mm,年相对湿度70%;年平均温度2.7℃,全年大于或等于10℃的积温为2526℃,年平均总日照时数2471.3 h。早霜一般在8月末出现,晚霜截止到5月末,无霜期约120—140 d。地带性土壤为典型暗棕壤[13]。
该研究选取40年生兴安落叶松人工林为研究对象,主要乔木层树种为落叶松,伴有少量的水曲柳、白桦、胡桃楸和紫椴等,土壤为典型暗棕色森林土壤。土壤表层(0—10 cm)的有机质、全氮和磷分别为(13.5±0.83)%、(0.96±0.12)%和(0.15±0.01)%;土壤表层(0—10 cm)的pH值为(6.28±0.09),土壤容重为(0.85±0.06) g/cm3。
1.2 样地设置和处理2011年4月在兴安落叶松人工林内设立12个10 m×20 m 的标准样地。样方之间留有15m左右的缓冲带以防止相互之间干扰。实验分为4个处理组,分别为对照处理(CK)、低氮处理(LN)、中氮处理(MN)和高氮处理(HN),每个处理组分成3个重复,即同一林分同一处理由随机选择的3个样方组成。在2011年5月至10月份,2012年5月份至10月份间,每月底喷洒硝酸铵(NH4NO3),每个样方每次将所需的氮肥溶解在50 L自来水中,然后采用背式喷雾器人工有序的均匀喷洒在林地,对照样方喷施等量的自来水。连续2a内共喷洒12次氮肥,生长季内,每月喷洒氮量保持一致,分别按0、50、100和150 kg N hm-2 a-1浓度进行施氮处理。
1.3 样木选择为减少树干呼吸的种内变异,在每块样地内严格按照以下标准筛选出3株落叶松作为测定样木:(1)树干胸径在(20±1.5) cm范围之内;(2)树高和冠幅基本一致;(3)树干通直,生长状态正常;(4)土壤条件排水较为良好。
1.4 树干呼吸和温度的测定于2012年4月底至10月底测定树干呼吸,半个月左右测定一次,生长季内共测定12次。采用石新立等[18]的测定方法,测定前,在选取的样木正北面距离地面1.3 m处安装1个内径为10.2 cm,高5 cm的PVC呼吸环。安装前,首先除去松脱的表层树皮、苔藓等杂物,避免伤及树皮活组织,影响树干呼吸测定;然后,按照要安装的树干表面弧度将呼吸环的一端打磨成形状与树干表面相吻合的弧形;利用100%中性硅酮胶将制打磨好的PVC环固定在树干表面上,并保持PVC呼吸环在整个测定期间位置不变且不漏气。
利用 LI-6400 便携式 CO2分析系统(LI-COR Inc. USA)进行树干呼吸的测定。考虑到树干呼吸具有明显的日变化格局,为更准确的比较树干呼吸对不同水平氮沉降的响应,将每次测定限定在晴天的10:00—12:00时间段内进行。测定时,直接在LI-6400-09同化室固定好的PVC环上进行测定,每次测定3个循环。同步,在PVC环上方5 cm处放置接触式温度记录仪测定树干表皮层下1 cm深处的温度。
1.5 树干氮素含量的测定在生长季较为旺盛的7月末,利用内径为5 mm的生长锥在每株样木PVC呼吸环的下方5 cm处钻取边材最外侧2 cm的树干样品,钻取之前首先需将树干表层的树皮轻轻除掉,并在取样之后立即用直径像吻合的树枝堵住树干伤口。取回的树干样品65℃恒温条件下至少烘48 h。采用半微量凯氏定氮法进行树干氮含量的测定,即称磨细烘干的树干样品(0.25 mm 筛)0.1 g,置于100 mL的消化管中,用浓硫酸和硝酸混合样消煮后,使用凯氏定氮仪进行蒸馏并用标准酸溶液滴定[28, 29, 30]。
1.6 数据分析由于树干表面的不规则性导致所测定的树干表面积和呼吸环的体积可能会不一致,因此所测得的树干呼吸速率(μmol CO2 m-2 s-1)需要进行校正[17, 18, 31],校正采用以下公式:

其中:

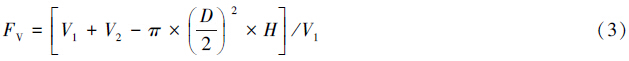
式中,RS为校正后的树干呼吸(μmol CO2 m-2 s-1),RM为校正前LI-6400系统测定的树干呼吸,S为系统默认的同化室面积(81.7 cm2),S0为实际测定的树干表面积(cm2),V1为同化室容积(991 cm3),V2为呼吸环的实际体积(cm3);D为呼吸环直径(10.2 cm),H为呼吸环高度(5 cm)。
采用公式(4)拟合树干呼吸速率与树干温度间的关系[32]:

温度系数Q10值的计算公式如下:

式中,RS为树干呼吸速率(μmol m-2 s-1);T为树干温度(℃);R0和β为常数。
数据分析采用SPSS13.0完成。利用LSD多重检验法检验不同处理间的差异显著性。RS生长季内的累积碳通量是基于实测数据建立的RS统计模型与连续树干温度监测数据相结合的方法进行估测。统计模型所需要的温度参数来自于在研究样地安装的温度自动记录仪Thermo Recorder Tr-71U (T&D Corp.Japan),每隔30 min记录1次树干表皮层下1 cm深处的温度。生长季期间的碳通量根据上述模型以天为步长累加而得。
2 结果 2.1 树干呼吸的季节动态不同氮处理条件下的树干呼吸速率均存在明显的季节变化,整个生长季内的呼吸速率呈单峰曲线,与树干温度的变化格局基本一致(图 1)。不同氮处理条件下的树干温度差异不限制(P > 0.05)。树干呼吸速率从春季4月末开始,随着树干温度的升高也逐渐上升,在7月底均到达峰值,8月份以后随着树干温度的下降,呼吸速率也开始逐渐降低,10末达到最低值。生长季内,对照、低氮、中氮、和高氮处理下的树干呼吸速率最小值分别为1.33、1.60、1.85 μmol m-2 s-1和1.73 μmol m-2 s-1,最高值分别为4.71、5.01、5.38 μmol m-2 s-1和5.96 μmol m-2 s-1。相应地,树干呼吸的离散程度也呈现出类似的季节变化趋势,7月份的标准误差是10月份的3.59倍。
其中,CK、LN、MN和HN分别表示对照处理(0 kg N hm-2 a-1)、低氮处理(50 kg N hm-2 a-1)、中氮处理(100 kg N hm-2 a-1)和高氮处理(150 kg N hm-2 a-1)。

|
| 图1 兴安落叶松树干温度与呼吸速率的季节变化特征 Fig.1 Seasonal variations of stem temperature and stem respiration rate of Larix gmelinii |
经过连续2个生长季的施氮处理,均显著提高了落叶松的树干呼吸速率(图 1b)。生长季内,对照、低氮、中氮、和高氮处理条件下的平均树干呼吸速率分别为(2.95±0.57)、(3.16±0.72)、(3.45±0.93) μmol m-2 s-1和(3.82±0.82) μmol m-2 s-1。而且随氮处理水平增加,生长季内树干表面释放的CO2通量也呈现逐渐上升的趋势,在高氮处理条件下达到峰值,为86.5g C/m2,比对照高出了将近30%(图 2)。高氮处理下的树干表面CO2通量与对照的差异达到显著水平(P < 0.05),但与低氮和中氮处理下的差异不显著。中氮处理条件下的碳通量也与对照的差异达到显著水平(P < 0.05),但与其它处理条件下的差异均不显著。低氮处理下的碳通量与各处理间的差异均未达到显著水平。

|
| 图2 生长季内不同氮处理条件下的树干表面CO2通量比较 Fig.2 Comparisons of the estimated stem surface CO2 flux among different N treatments in the growing season |
对不同氮处理条件下的树干呼吸速率与树干温度进行的相关性分析表面,呼吸速率均与树干温度有显著性的正相关,并且树干1 cm深处的温度可以解释树干呼吸速率64%—82%的变化(表 1)。
研究发现,温度敏感性(Q10)对不同浓度的氮沉降的响应也不同,随着氮处理浓度增加,Q10值逐渐变大(表 1)。对照、低氮、中氮、和高氮处理下的Q10值分别为1.67、1.80、2.01和2.54,高氮处理条件下的Q10与对照、低氮的差异达到显著水平(P < 0.05),但与中氮处理下的差异不显著。中氮处理条件下的Q10也与对照、低氮的差异达到显著水平(P < 0.05)。低氮处理条件下的Q10与对照组间的差异未达到显著水平,与其它各组间的差异均达到显著水平(P < 0.05)。
| 处理 Treatment | 回归方程 Regression model | Q10 | R2 | P | n |
| CK | y =1.085e0.051T | 1.665 | 0.64 | < 0.001 | 30 |
| LN | y =1.352e0.059T | 1.804 | 0.75 | < 0.001 | 30 |
| MN | y =1.493e0.070T | 2.014 | 0.82 | < 0.001 | 28 |
| HN | y =1.620e0.093T | 2.535 | 0.73 | < 0.001 | 26 |
其中,CK、LN、MN和HN分别表示对照处理(0 kg N hm-2 a-1)、低氮处理(50 kg N hm-2 a-1)、中氮处理(100 kg N hm-2 a-1)和高氮处理(150 kg N hm-2 a-1)。
2.4 树干呼吸速率与树干氮元素含量的关系随着氮处理浓度提高,树干氮含量也随之上升。其中对照、低氮、中氮、和高氮处理下氮元素的浓度分别为1.00、1.32、2.02和2.35 g/kg。高氮和中氮处理条件下的树干氮含量与对照、低氮的差异均达到显著水平(P < 0.05),但两者之间的差异不显著。低氮处理条件下的树干氮含量也与对照组间的差异达到显著水平(P < 0.05)。对树干呼吸速率和树干氮浓度的关系进行线性回归分析,结果表明树干氮含量与树干呼吸速率之间的相关性较弱,树干氮浓度仅解释了生长季内树干呼吸速率38.3%的变化。

|
| 图3 树干呼吸速率与树干氮浓度之间的关系 Fig.3 The relationship between stem respiration rates and stem nitrogen concentration among different N treatments in the growing season |
本研究结果表明,测定期间树干呼吸速率表现为明显的单峰型季节变化模式,生长季的树干呼吸显著高于非生长季。而且不同氮浓度处理均不同程度的促进了落叶松树干呼吸速率,其中以高氮浓度处理条件下树干表面释放的CO2通量最大,这与以往的研究结果相一致[14, 22, 24, 25]。树干呼吸主要由提供能量合成新组织的生长呼吸和维持活细胞正常生命活动的维持呼吸两部分组成[14]。本研究中氮沉降促进树干呼吸的可能原因主要有两个:一是氮元素增加促进了植物组织的生长,从而引起植物呼吸尤其是生长呼吸的增加[33]。目前大多数研究认为温带森林生长主要是氮元素的限制[34],大气氮沉降通过提高土壤的有效氮水平,进而促进了植物的生长[35],而植物的生长速率与树干的生长呼吸存在密切的正相关关系[14, 24]。这很好的解释了本研究中生长季旺盛季节,氮处理促进了树干呼吸速率的现象;二是外界氮输入导致了植物细胞蛋白质含量的增加。植物组织中90%的氮参与了蛋白质的合成,外界氮输入之后会使植物组织蛋白质合成数量增加,而这些蛋白质的修复和更新需要大量的能量支持,进而促使树干呼吸尤其是维持呼吸的上升[22]。这与本研究中生长季末期,氮处理仍提高了树干呼吸速率这一结果相吻合。
本文研究还首次比较了不同氮处理条件下树干呼吸的温度敏感性(Q10)指数,结果表明随着氮输入增加,Q10也随之增加,这可能是由于树干呼吸中维持呼吸和生长呼吸对氮沉降的响应模式不同引起的[36]。树干组织氮含量对呼吸的影响(解释38.3%的变化)可能也会间接影响树干呼吸对温度的敏感性。尽管不同氮处理的树干呼吸对温度的响应程度不一,但总体上两者均存在极显著的指数函数关系(P < 0.01)。树干温度是影响树干呼吸的重要环境因子,一方面是由于在一定范围内温度升高增强了参与树干呼吸作用的酶活性,促进了树干呼吸速率[24];另一方面,主要是温度升高促进了植物组织细胞呼吸、降低了CO2在水中的溶解度、提高了CO2 的扩散系数,从而导致树干表面释放更多的CO2通量[37]。另外,本研究基于季节尺度计算而得到的树干呼吸温度敏感性与以往温带森林研究的结果范围相符合[16, 19, 25, 37],但要明显高于石新立等[18]在该地区测定的兴安落叶松树干呼吸温度敏感性结果,尽管采用的研究方法相一致。导致不同研究中Q10变异较大的原因可能由于本研究中树干呼吸的测定期间为4—10月份,测定间隔为15 d左右,而石新立等[18]的测定期间为5—9月份,测定时间尺度不同,直接会造成结果的很大差异;另外,本研究所选取样木的胸径在20 cm左右,明显小于石新立等[18]测定样木的胸径(波动在15.5—39.3 cm,平均为27.6 cm)。
随着氮输入增加,树干的氮含量也逐渐升高,这与以往的研究结果相一致[14, 22, 24]。例如Maier对火炬松林连续3a施肥后,发现氮输入样地的树干氮含量比对照高出了1.5倍左右[14]。这可能是因为每月喷洒的NH4NO3为可溶性氮,该部分氮很容易被植物组织吸收利用,进而导致树干氮含量升高。但是,目前很多研究,即使都采用同一方法,其结果仍然表明树干氮含量与树干呼吸之间的关系存在较大的差异。例如,本文与Lavigne等[38]很多研究的结果都表明树干氮含量与树干呼吸速率之间的相关较弱,而Vose等[9]的研究表明树干氮含量与树干呼吸速率之间存在显著的正相关关系。导致树干氮含量与树干呼吸之间相关较弱的原因之一是由于树干呼吸释放的CO2的来源和去向十分复杂,实际上采用红外线气体分析仪LI-6400同化室所测得的变量只是实际树干呼吸产生的CO2的一部分,而剩余的部分CO2可能还溶解于树干液流中,或被带至冠层上方,或被树皮光合作用所利用[15, 37];其二是进入植物组织的氮素并没有向蛋白质分配,而是以游离氨基酸的形态贮存着,这部分氮将可能不会对呼吸速率造成影响[9]。
通过模拟氮沉降对兴安落叶松树干呼吸的影响,发现施肥2a后,随着氮输入的增多,树干呼吸速率、树干氮含量和温度敏感性都呈现逐渐增加的趋势。可见,氮沉降是影响温带森林树干呼吸速率非常重要的因素之一。
| [1] | LeBauer D S, Treseder K K. Nitrogen limitation of net primary productivity in terrestrial ecosystems is globally distributed. Ecology, 2008, 89(2): 371-379. |
| [2] | 李德军, 莫江明, 方运霆, 彭少麟, Gundersen P. 氮沉降对森林植物的影响. 生态学报, 2003, 23(9): 1891-1900. |
| [3] | Vitousek P M, Aber J D, Howarth R W, Likens G E, Matson P A, Schindler D W, Schlesinger W H, Tilman G D. Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications, 1997, 7(3): 737-750. |
| [4] | Galloway J N, Townsend A R, Erisman J W, Bekunda M, Cai Z C, Freney J R, Martinelli L A, Seitzinger S P, Sutton M A. Transformation of the nitrogen cycle: recent trends, questions, and potential solutions. Science, 2008, 320(5878): 889-892. |
| [5] | Galloway J N, Dentener F J, Capone D G, Boyer E W, Howarth R W, Seitzinger S P, Asner G P, Cleveland C C, Green P A, Holland E A, Karl D M, Michaels A F, Porter J H, Townsend A R, Vörösmarty C J. Nitrogen cycles: past, present, and future. Biogeochemistry, 2004, 70(2): 153-226. |
| [6] | Liu X J, Zhang Y, Han W X, Tang A H, Shen J L, Cui Z L, Vitousek P, Erisman J W, Goulding K, Christie P, Fangmeier A, Zhang F S. Enhanced nitrogen deposition over China. Nature, 2013, 494(7438): 459-462. |
| [7] | Ryan M G. Effects of climate change on plant respiration. Ecological Applications, 1991, 1(2): 157-167. |
| [8] | 曾小平, 彭少麟, 赵平.广东南亚热带马占相思林呼吸量的测定. 植物生态学报, 2000, 24(4): 420-424. |
| [9] | Vose J M, Ryan M G. Seasonal respiration of foliage, fine roots, and woody tissues in relation to growth, tissue N, and photosynthesis. Global Change Biology, 2002, 8(2): 182-193. |
| [10] | 肖复明, 汪思龙, 杜天真, 陈龙池, 于小军. 湖南会同林区杉木人工林呼吸量测定. 生态学报, 2005, 25(10): 2514-2519. |
| [11] | Edwards N T, Hanson P J. Stem respiration in a closed-canopy upland oak forest. Tree Physiology, 1996, 16(4): 433-439. |
| [12] | Edwards N T, Tschaplinski T J, Norby R J. Stem respiration increases in CO2-enriched sweetgum trees. New Phytologist, 2002, 155(2): 239-248. |
| [13] | Wang W J, Yang F J, Zu Y G, Wang H M, Kentaro T, Kaichiro S, Takayoshi K. Stem respiration of a larch (Larix gmelinii) plantation in Northeast China. Acta Botanica Sinica, 2003, 45(12): 1387-1397. |
| [14] | Maier C A. Stem growth and respiration in loblolly pine plantations differing in soil resource availability. Tree Physiology, 2001, 21(16): 1183-1193. |
| [15] | Teskey R O, Saveyn A, Steppe K, McGuire M A. Origin, fate and significance of CO2 in tree stems. New Phytologist, 2008, 177(1): 17-32. |
| [16] | 王淼, 武耀祥, 武静莲. 长白山红松针阔叶混交林主要树种树干呼吸速率. 应用生态学报, 2008, 19(5): 956-960. |
| [17] | 王淼, 姬兰柱, 李秋荣, 肖冬梅, 刘海良. 长白山地区红松树干呼吸的研究. 应用生态学报, 2005, 16(1): 7-13. |
| [18] | 石新立, 王传宽, 许飞, 王兴昌. 四个温带树种树干呼吸的时间动态及其影响因子. 生态学报, 2010, 30(15): 3994-4003. |
| [19] | 王文杰, 王慧梅, 祖元刚, 李雪莹, 小池孝良. 林木非同化器官与土壤呼吸的温度系数Q10值的特征分析. 植物生态学报, 2005, 29(4): 680-691. |
| [20] | 许飞, 王传宽, 王兴昌. 东北东部14个温带树种树干呼吸的种内种间变异. 生态学报, 2011, 31(13): 358-3589. |
| [21] | Etzold S, Zweifel R, Ruehr N K, Eugster W, Buchmann N. Long-term stem CO2 concentration measurements in Norway spruce in relation to biotic and abiotic factors. New Phytologist, 2013, 197(4): 1173-1184. |
| [22] | Ryan M G. Foliar maintenance respiration of subalpine and boreal trees and shrubs in relation to nitrogen content. Plant, Cell and Environment, 1995, 18(4): 765-772. |
| [23] | Wieser G, Bahn M. Seasonal and spatial variation of woody tissue respiration in a Pinus cembra tree at the alpine timberline in the central Austrian Alps. Trees, 2004, 18(5): 576-580. |
| [24] | Stockfors J, Linder S. Effect of nitrogen on the seasonal course of growth and maintenance respiration in stems of Norway spruce trees. Tree Physiology, 1998, 18(3): 155-166. |
| [25] | Ryan M G, Hubbard R M, Pongracic S, Raison R J, McMurtrie R E. Foliage, fine-root, woody-tissue and stand respiration in Pinus radiata in relation to nitrogen status. Tree Physiology, 1996, 16(3): 333-343. |
| [26] | 王秀伟, 毛子军, 孙涛, 吴海军. 春、秋季节树干温度和液流速度对东北3树种树干表面CO2 释放通量的影响. 生态学报, 2011, 31(12): 3358-3367. |
| [27] | 刘世荣. 兴安落叶松人工林生态系统营养元素生物地球化学循环特征. 生态学杂志1992, 11(5): 1-6. |
| [28] | Sun T, Mao Z J. Functional relationships between morphology and respiration of fine roots in two Chinese temperate tree species. Plant and Soil, 2011, 346(1/2): 375-384. |
| [29] | Sun T, Mao Z J, Dong L L, Hou L L, Song Y, Wang X W. Further evidence for slow decomposition of very fine roots using two methods: litterbags and intact cores. Plant and Soil, 2013, 366(1/2): 633-646. |
| [30] | Sun T, Mao Z J, Han Y Y. Slow decomposition of very fine roots and some factors controlling the process: a 4-year experiment in four temperate tree species. Plant and Soil, 2013, 372(1/2): 445-458. |
| [31] | Zha T, Kellomǎki S, Wang K Y, Ryyppǒ A, Niinistǒ S. Seasonal and annual stem respiration of Scots pine trees under boreal conditions. Annals of Botany, 2004, 94(6): 889-896. |
| [32] | Davidson E A, Belk E, Boone R D. Soil water content and temperature as independent or confounded factors controlling soil respiration in a temperate mixed hardwood forest. Global Change Biology, 1998, 4(2): 217-227. |
| [33] | Vitousek P M, Aber J D, Howarth R W, Likens G E, Matson P A, Schindler D W, Schlesinger W H, Tilman D G, Human alteration of the global nitrogen cycle: sources and consequences. Ecological Applications, 1997, 7(3): 737-750. |
| [34] | Vitousek P M, Howarth R W. Nitrogen limitation on land and sea: how can it occur? Biogeochemistry,1991, 13(2): 87-115. |
| [35] | Frink C R, Waggoner P E, Ausubel J H. Nitrogen fertilizer: retrospect and prospect. Proceedings of the National Academy of Sciences of the United States of America, 1999, 96(4): 1175-1180. |
| [36] | Bredu S A, Yokota T, Hagihara A. Temperature effect on maintenance and growth respiration coefficients of young, field- grown Hinoki cypress (Chamaecyparis obtusa). Ecological Research, 1997, 12(3): 357-362. |
| [37] | McGuire M A, Cerasoli S, Teskey R O. CO2 fluxes and respiration of branch segments of sycamore (Platanus occidentalis L.) examined at different sap velocities, branch diameters and temperatures. Journal of Experimental Botany, 2007, 58(3): 2159-2168. |
| [38] | Lavigne M B, Ryan M G. Growth and maintenance respiration rate of aspen, black spruce and jack pine stems at northern and southern BORE AS sites. Tree Physiology, 1997, 17(8/9): 543-551. |