 2015, Vol. 35
2015, Vol. 35文章信息
- 刘志理, 金光泽
- LIU Zhili, JIN Guangze
- 利用凋落物法和林木因子模拟小兴安岭阔叶红松林叶面积指数
- Using litter collection and tree data to model leaf area index in the mixed broadleaved-Korean pine forest in the Xiaoxing'an Mountains, China
- 生态学报, 2015, 35(10): 3190-3198
- Acta Ecologica Sinica, 2015, 35(10): 3190-3198
- http://dx.doi.org/10.5846/stxb201405211043
-
文章历史
- 收稿日期:2014-05-21
- 网络出版日期:2014-08-27






叶面积指数(LAI)指地表单位面积上总叶片表面积的一半[1]。叶片面积影响着植物的光合作用和蒸腾作用,因此,LAI是模拟植物冠层光合作用、蒸发散以及微气象最常用的因子之一[2, 3]。直接法测定的LAI能够代表真实LAI,经常被用于校准光学仪器法间接测定值[4, 5, 6]。在直接法中,通过收集凋落物的方法来测定森林生态系统的LAI比破坏性取样法和异速生长方程法更易实施,且不具有破坏性[7, 8, 9]。然而,相对于光学仪器法,凋落物法仍费时费力[10],因此,如何改进凋落物法使之更快捷、有效地测定森林生态系统的LAI具有重要意义。
凋落物法是指通过测定凋落物收集器中凋落物的干重,然后结合比叶面积(SLA)来测定LAI。目前,凋落物法不仅常用于测定落叶阔叶林的LAI[5, 11, 12, 13],在常绿针叶林中也得到广泛应用[14, 15, 16, 17]。SLA是影响凋落物法测定LAI精度的决定因子[7, 18, 19]。不同的树种之间,尤其是针叶树种与阔叶树种之间的SLA存在较大差异[20],因此,为准确测定整个林分的LAI,主要树种的SLA均需要测定,而不能用不同树种的平均SLA代替[21]。这就需要把收集的凋落叶按树种分类,然而不同树种凋落叶的鉴别分类需要一定的专业技能,且耗费时间,尤其是物种丰富的原始林[7]。Nasahara等[10]建议利用各树种的胸高断面积(BA)占所有树种总BA的比例代替凋落物收集器中各树种的叶凋落量占总叶量的比例来分配凋落叶。这种利用林木因子等信息来预测各树种的凋落叶的方法,极大地提高了凋落物法的有效性。林木的每木检尺主要指测定林分内树木的树种、胸径、树高、坐标等林木因子,这些因子不仅能提供树种组成、树种优势度,还能提供每棵树在林分内的空间信息。
Ishihara和Hiura[7]提出3种基于凋落物法和林分每木检尺数据的模型能够准确预测落叶阔叶林的LAI,为提高凋落物法测定落叶阔叶林LAI的效率奠定了基础。3种模型包括: 1) 平均优势度模型,即假设一个凋落物收集器内的总凋落物干重能平均分配给各树种; 2) 林分优势度模型,将凋落物按照各树种在整个样地内的相对优势度分配; 3) 局域优势度模型,将凋落物按照各树种在凋落物收集器一定距离范围内的相对优势度分配。研究结果表明3种模型预测的LAI均与实测LAI显著相关(P<0.001),最小相关系数为0.988; 局域优势模型预测的LAI与实测LAI具有最高的R2值(0.997),且比其他两个模型能更准确地预测LAI的空间变异。然而,这3种模型是否适用于常绿针叶和落叶阔叶混交的针阔混交林LAI的测定尚未得到验证。尽管此3种模型不适用于未设置凋落物收集器的林分LAI测定,但相对于其他直接法仍具有明显的优势。
利用直接法测定森林生态系统的LAI通常先测定林分内各树种的LAI,然后综合得到整个林分的LAI。各树种的LAI与胸径,BA或边材面积关系密切,破坏性取样法和异速生长方程法多是基于以上因子来预测LAI。如: 李轩然等[22]建立了胸径和叶子生物量的方程,再结合SLA测定了湿地松(Pinus elliotii)、马尾松(Pinus massoniana)和杉木(Cunninghamia lanceolata)的LAI; Guiterman等[17]通过叶面积与边材面积的异速生长方程测定美国白松(Pinus strobus)人工林的LAI,并发现凋落物法测定的LAI与各树种的BA呈线性关系。Jonckheere 等[23]通过建立叶面积与胸径或BA的相关关系测定了欧洲赤松林(Pinus sylvestris)的LAI。然而,在非破坏性条件下,国内对于如何建立LAI与BA相关关系的研究尚少。
本研究以东北东部山区的顶极植被-阔叶红松林(Pinus koraiensis)为研究对象。利用凋落物法测定该林型最大LAI时期的LAI,以此为参考值验证平均优势度模型、林分优势度模型和局域优势度模型预测针阔混交林LAI的有效性; 并以该区域的主要树种为研究对象,探讨凋落物法测定的LAI与其BA的相关关系。研究结果可为快捷、有效地测定针阔混交林的LAI提供参考,进一步为模拟该区域的生理生态过程提供科学依据。
1 材料和方法 1.1 研究区域概况野外调查在黑龙江省凉水国家级自然保护区(47° 10′ 50″ N,128° 53′ 20″ E)进行。保护区位于黑龙江省伊春市带岭区,地处小兴安岭南部达里带岭支脉的东坡,地形比较复杂,最高山脉海拔707.3 m。该区属温带大陆性季风气候,年均气温-0.3℃,年均最高气温7.5℃,年均最低气温-6.6 ℃; 年均降水量676 mm,降水集中在7月; 年积雪期130 —150 d,年无霜期100 —120 d。该区域的顶极植被群落是以红松为建群种的阔叶红松林,常绿针叶树种包括红松、冷杉(Abies nephrolepis)和云杉(Picea spp.); 落叶阔叶树种主要包括紫椴(Tilia amurensis)、五角槭(Acer mono)、枫桦(Betula costata)和裂叶榆(Ulmus laciniata)等(表 1)。
| 树种 Species | 密度 Density | 胸高断面积 Basal area | 比叶面积SLA Specific leaf area | |||||
| 样地 Plot/ (No./hm2) | 小样方 Subplot | 样地 Plot/ (m2/hm2) | 小样方 Subplot | |||||
| 范围 Range (20 m×20 m) | 标准偏差 S.D. | 范围 Range (20 m×20 m) | 标准偏差 S.D. | 均值 Mean/ (cm2/g) | 标准偏差 S.D. | |||
| S.D.为标准偏差 | ||||||||
| 红松 Pinus koraiensis | 122 (4.0) | 0—14 | 2.7 | 24.45 (56.8) | 0—2.66 | 0.57 | 83.79 | 3.74 |
| 紫椴 Tilia amurensis | 82 (2.7) | 0—21 | 3.8 | 3.46 (8.1) | 0—0.87 | 0.17 | 243.59 | 14.26 |
| 冷杉 Abies nephrolepis | 111 (3.6) | 0—15 | 3.1 | 3.42 (8.0) | 0—0.50 | 0.11 | 80.80 | 5.43 |
| 五角槭 Acer mono | 163 (5.3) | 0—15 | 3.1 | 2.49 (5.8) | 0—0.39 | 0.08 | 305.04 | 50.08 |
| 枫桦 Betula costata | 49 (1.6) | 0—16 | 3.1 | 2.02 (4.7) | 0—0.98 | 0.16 | 199.79 | 11.55 |
| 裂叶榆 Ulmus laciniata | 104 (3.4) | 0—25 | 4.4 | 1.43 (3.3) | 0—0.28 | 0.08 | 261.40 | 5.65 |
| 水曲柳 Fraxinus mandshurica | 29 (0.9) | 0—7 | 1.5 | 1.21 (2.8) | 0—0.59 | 0.11 | 338.36 | 13.63 |
| 云杉 Picea spp. | 20 (0.6) | 0—3 | 0.9 | 1.18 (2.7) | 0—0.28 | 0.08 | 59.41 | 9.70 |
| 大青杨 Populus ussuriensis | 9 (0.3) | 0—4 | 0.9 | 0.45 (1.0) | 0—0.90 | 0.11 | 198.73 | 24.72 |
| 花楷槭 Acer ukurunduense | 226 (7.3) | 0—41 | 7.4 | 0.43 (1.0) | 0—0.11 | 0.02 | 350.67 | 8.56 |
| 青楷槭 Acer tegmentosum | 111 (3.6) | 0—27 | 5.6 | 0.40 (0.9) | 0—0.07 | 0.02 | 263.26 | 32.66 |
| 春榆 Ulmus japonica | 33 (1.1) | 0—21 | 3.7 | 0.36 (0.8) | 0—0.53 | 0.07 | 179.35 | 18.05 |
| 毛榛子 Corylus mandshurica | 1150 (37.3) | 1—178 | 38.6 | 0.34 (0.8) | 0—0.06 | 0.01 | 374.89 | 66.93 |
| 其他 Others | 873 (28.3) | 14—242 | 41 | 1.36 (3.2) | 0.01—0.16 | 0.11 | - | - |
| 总计 Total | 3083 | 60—267 | 42.6 | 43.02 | 0.56—3.38 | 0.54 | - | - |
研究样地位于凉水典型阔叶红松林9 hm2监测样地的中心区,面积为160 m × 160 m,分为64个20 m × 20 m的小样方。2010年8月份,对于样地内胸径 ≥1 cm 的物种进行每木检尺,主要测定胸径、树高、坐标等。在每个小样方的中心处,设置一个凋落物收集器,共计64个。凋落物收集器用直径为8 mm的铁丝和尼龙网围成(孔径1 mm,深0.5—0.6 m),网口为正方形,面积为0.5 m2,凋落网底离地面0.5 m左右。
1.2 凋落物法测定叶面积指数 1.2.1 凋落物的收集凋落物收集主要在2010年9月初至2011年8月初进行。其中,2010年9月至12月和2011年5月至8月初,每月初收集1次; 而2011年1月至4月,因样地内积雪过多,收集难度大,该段时间内不进行凋落物的收集,即2011年5月初收集的凋落物是2010年12月至2011年4月期间内凋落的。收集的凋落物及时带回实验室,将凋落叶按树种进行分类并测其湿重。随机选取一定数量的样品,烘干至恒重后测其干重。再结合各树种的SLA得到各样点各树种各时期凋落叶产生的LAI。
1.2.2 比叶面积的测定SLA指单位干重的叶面积[24]。研究表明同一树种的SLA存在季节性差异[18, 25]。因此,为减小SLA季节性差异对估测LAI产生的影响,本研究分别在8月、9月、10月测定样地内主要树种的SLA。本研究测定了红松、冷杉、云杉、紫椴、五角槭、枫桦、裂叶榆、水曲柳(Fraxinus mandshurica)、大青杨(Populus ussuriensis)、花楷槭(Acer ukurunduense)、青楷槭(Acer tegmentosum)、春榆(Ulmus japonica)及毛榛子(Corylus mandshurica)共3种针叶树种和10种阔叶树种的SLA。阔叶和针叶树种采用不同方法测定其SLA。阔叶树种,各树种从凋落物收集器内随机选取50个健康平整的样叶,利用扫描仪测定样叶的叶面积,不平整的样叶需浸在水中直至平整; 针叶树种,各树种在凋落收集器内随机选取400个样针,利用排水法测定其半表面积[26],操作细节参照了Liu等[27]的文献。将样叶烘干至恒重后,分别称重(精确到0.001 g)。根据SLA定义即可得到各树种3个时期的SLA。对于未测定SLA的时期,采用3次的均值。对于未测定SLA的阔叶树种,其SLA采用10种阔叶树种的均值。
1.2.3 叶面积指数的测定本研究关注2010年阔叶红松林最大LAI时期即8月初的LAI。阔叶红松林内阔叶和针叶树种8月初的LAI根据不同时期的凋落物数据结合SLA计算。各阔叶树种8月初的LAI通过累加落叶季节(2010年9月至11月)凋落叶产生的LAI得到。对于针叶树种,利用以下公式可得到林冠内的总LAI[28]:

式中,LAIi-total为针叶展出i年后林冠内存留针叶的总LAI,LAIi为林冠内针叶展出i年后存留的LAI,i为针叶展出后存活的年数。研究区域的阔叶红松林为成熟林,可假设每年新生针叶的总数量保持不变,任何一年内观测的LAIi都可代表该林分的平均状态。因此,本研究以1a为周期(2010年9月至2011年8月初),先测定1年内凋落针叶产生的LAI,然后乘以针叶的平均叶寿命得到2010年8月初各针叶树种的LAI。本研究通过实地采集样枝的方法获得红松、冷杉和云杉针叶的平均叶寿命分别为(3.08 ± 0.25)(平均值±标准偏差)a、(3.68 ± 0.52)a、(3.91 ± 0.27)a,具体测定方法参照刘志理等[28]的文献。综合阔叶和针叶树种的LAI即可得到整个林分8月初的LAI,该方法得到的LAI为实测LAI,用于检验模型预测LAI的有效性。
1.3 模型预测叶面积指数参照Ishihara和Hiura[7]和Nasahara等[10]的研究结果,本研究利用各树种的BA来评定各树种的优势度。
1.3.1 平均优势度模型该模型假设凋落物收集器i内的总凋落叶干重(LFi,g/m2)能够平均分配给13个树种,即利用以下公式可得到凋落物收集器i内所有树种的总LAI(LAIi)[7]:
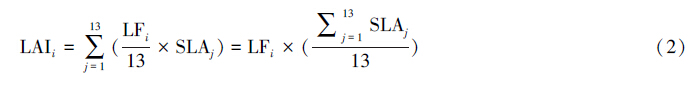
式中,SLAj为树种j的比叶面积。LFi为凋落物收集器i内的总凋落叶干重(即各时期凋落叶总和),其中阔叶树种为2010年9月至2010年11月内所有凋落叶,而针叶树种一直存在凋落现象,因此收集器i内针叶树种的总凋落叶是根据2010年9月至2011年8月内的总凋落针叶乘以其针叶平均叶寿命计算得到。
1.3.2 林分优势度模型该模型根据各树种在整个样地内的相对优势度来预测凋落物收集器i对应的总LAI (LAIi)[7]:

式中,Dj为树种j在整个样地内的相对优势度,即树种j的胸高断面积(BAj,m2)占整个样地内13个树种总BA的比例,根据下式得到:
1.3.3 局域优势度模型

该模型是根据各树种在距离凋落物收集器一定范围内的相对优势度来预测凋落物收集器i对应的总LAI(LAIi)[7]:

式中,Dij为树种j在距离凋落物收集器i 10m距离内的相对优势度,根据下式计算得到:

Staelens等[29]的研究结果表明凋落物收集器内的凋落叶总量与以收集器为中心一定半径范围内树木的总BA显著相关,且半径在1 m到10 m内其相关性随半径增加而增加,而半径继续增加其相关性无明显变化。Ishihara和Hiura[7]的研究进一步表明10 m为最佳距离。因此,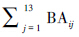 为以凋落物收集器i为中心,半径为10 m范围内13个树种的总BA。
为以凋落物收集器i为中心,半径为10 m范围内13个树种的总BA。
SLA的测定是凋落物法测定LAI中的关键步骤,同时加大了凋落物法的实施难度,尤其是测定物种丰富的森林生态系统中各树种的SLA费时费力。因此,本研究基于12个树种的SLA,检验测定SLA的树种数量对于LAI测量精度的影响,根据下式计算凋落物收集器i的总LAI(LAIi)[7]:
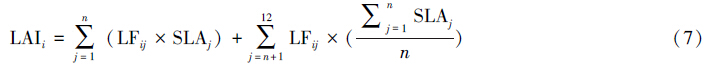
右式中第一部分为凋落物收集器i内n个树种(j = 1,2,3,…,n,n 最大为11)的总LAI,第二部分为估测的剩余凋落叶产生的LAI,由剩余凋落叶干重除以n个已知树种的平均SLA得到。当n = 12时,第二部分为0,公式(7)计算得到实测LAI。根据树种的BA占样地内总BA的比例(即相对优势度)由高到低依次选择树种,即j = 1时,所选树种为相对优势度最高的红松,依次类推(表 1)。本研究中大青杨未用于SLA有效性的测定,因为其总BA主要是一棵大树的贡献(BA占总BA的79%),这使之凋落叶的分布过于集中,缺乏代表性。
为检验公式(7)估测LAI的准确性,将之与实测LAI进行回归分析,为方便比较,截距设置为0。检验参数为斜率和调整后的决定系数(R2)。
1.5 叶面积指数与胸高断面积的回归分析以64个20 m × 20 m的小样方为单位,以红松、冷杉、紫椴、五角槭、枫桦和裂叶榆为对象,根据每木检尺数据计算小样方内各树种的BA,然后与凋落物收集器内对应凋落叶得到的LAI进行回归分析。
2 结果与分析 2.1 模型比较3种模型预测的LAI与实测LAI均显著相关(P<0.01)(图 1)。局域优势度模型预测的LAI与实测LAI具有最大的R2及最小的RMSE,分别为0.97和0.42,表明该模型比平均优势度模型和林分优势度模型能更准确地预测LAI及其空间变异。平均优势度模型和林分优势度模型预测的LAI与实测LAI具有相同的R2值(0.76),但前者与实测LAI的RMSE略大于后者,值分别为1.10和1.09。实测LAI,平均优势度模型,林分优势度模型及局域优势度模型预测的LAI分别为6.88(S.D. = 2.27)、12.44(4.23)、7.86(2.62)及7.41(2.32) (表 2),表明3种模型预测的LAI均高于实测LAI。平均优势度模型预测LAI的效果最差,预测值比实测值高估81%; 林分优势度模型预测效果稍好,比实测值高估14%; 局域优势度模型预测效果最好,与实测值的差异小于10%,表明在预测LAI时,应考虑LAI的空间特性,不同方法测定LAI均具有较大标准偏差(表 2),说明阔叶红松林的LAI存在明显的空间变异。

|
| 图1 实测叶面积指数与3种模型预测的叶面积指数的回归分析(虚线为1∶1线) Fig.1 Regression analysis between measured and predicted LAI (leaf area index) for three models (n=64). The dotted lines represent the 1∶1 relationship |
| 数值 Values | 叶面积指数 LAI/(m2/m2) | 差异Difference/% | |||||
| 凋落物法 Litter collection method | 平均优势度模型 Equal dominance model | 林分优势度模型 Stand dominance model | 局域优势度模型 Local dominance model | 平均优势度模型 Equal dominance model | 林分优势度模型 Stand dominance model | 局域优势度模型 Local dominance model | |
| 差异(%)=(模型预测叶面积指数-实测叶面积指数)/实测叶面积指数×100% | |||||||
| 平均值 Mean | 6.88 | 12.44 | 7.86 | 7.41 | 81 | 14 | 9 |
| 标差 S.D. | 2.27 | 4.23 | 2.62 | 2.32 | 30 | 19 | 9 |
总体来看,随测定SLA树种数量的增加,斜率、调整后的R2与1的距离均越来越近(2个树种时除外)(图 2)。当选择1—7个树种的SLA计算LAI时,预测精度波动较大,而当选择≥8个树种的SLA计算LAI时,斜率、调整后的R2分别大于0.970和0.986,其测量精度高于97%,表明最少选择8个树种的SLA才能准确地测定阔叶红松林的LAI。

|
| 图2 实测叶面积指数与选择不同数量树种的比叶面积计算的叶面积指数间的斜率,调整后的决定系数 (R2) Fig.2 Slopes of the regression line between measured LAI and LAI calculated using SLA of n dominant species,adjusted coefficients of determination (R2) |
图 3表明利用凋落物法测定6个树种的LAI与其BA均显著相关(P<0.01)。在每个小样方内均有红松分布(n = 64,图 3),主要源于红松是该区域的建群种(相对优势度为56.8%)。总体来看,LAI与BA的相关性在针叶树种和阔叶树种间存在明显差异,例如红松和冷杉,其LAI和BA回归方程的指数分别为0.7972和0.9539; 相对而言,阔叶树种间LAI和BA回归方程的指数均小于针叶树种,如裂叶榆对应的指数最大为0.6814。裂叶榆的LAI与其BA具有最优相关性,R2 = 0.93,而枫桦、紫椴、冷杉、红松和五角槭,R2分别为0.89、0.81、0.81、0.78和0.67。

|
| 图3 小兴安岭6种主要树种的叶面积指数与其胸高断面积的回归分析 Fig.3 Regression analysis between LAI and basal area for six major species in the Xiaoxing′an Mountains,China |
平均优势度模型、林分优势度模型及局域优势度模型预测落叶阔叶林LAI的准确性均大于91%[7],高于预测针阔混交林(阔叶红松林)的LAI,主要源于针叶树种和阔叶树种间的SLA存在明显差异。本研究中不同树种SLA的变异系数大于Ishihara和Hiura[7]的研究结果,二者的值分别为46.4%和17.9%,此外,本研究中未考虑冠层中叶片的垂直结构对SLA产生的影响。平均优势度模型预测针阔混交林的LAI的效果最差,比实测值高估81%,表明平均优势度模型不适于预测针阔混交林LAI。在具备主要树种的SLA及BA条件下,林分优势度模型能够用于针阔混交林LAI的预测,精度达86%。局域优势度模型预测针阔混交林LAI的效果最优,精度高于90%,表明该模型不仅适用于预测落叶阔叶林的LAI,在针阔混交林中也适用; 然而该模型需要额外测定树木在林分中的空间信息(坐标)[7]。此外,为准确估测阔叶红松林LAI,应考虑测定树种的SLA,当测定树种数量过少时会出现低估或高估LAI的现象(图 2),Ishihara和Hiura[7]也得到类似结论; 但Kalácska等[21]发现基于一些树种的平均SLA预测LAI时总会出现高估现象,与树种数量无关,这些现象可能主要源于林分的物种组成及树种特性的差异。本研究表明,最少测定8个主要树种的SLA,其BA占12个物种总BA的95%,才能准确预测针阔混交林的LAI(精度高于97%)。林分优势度模型和局域优势度模型均不需要将凋落叶按树种进行分类,这大大提高了凋落物法的效率。
目前,异速生长方程法也是测定森林生态系统LAI常用的直接方法[30, 31],但相对于局域优势度模型法,需要耗费更多的人力财力,因为不仅同一地域不同树种间的异速生长方程存在差异,同一树种不同地域间的异速生长方程也可能存在差异[7, 26, 32]。然而局域优势度模型受凋落物收集器的限制,不适用于未设置收集器林分LAI的测定。相对而言,光学仪器法能方便、快捷地测定森林生态系统的LAI,但其存在明显低估现象,精度需要直接测定值进行校准已得到广泛报道[15, 33, 34, 35]。因此,如何提高直接法的效率或如何提高模型预测的精度应给予更多关注。
本研究基于凋落物法,建立了小兴安岭6个主要树种的LAI与其BA的回归关系,该方法避免了破坏性取样。研究表明,6个树种的LAI与BA均显著相关(P<0.01),最小R2为0.67。其他学者也得到类似结论,如Deblonde等[36]通过采伐样树,拟合叶面积与胸高处边材面积的方程,直接估测了加拿大松树林的脂松(Pinus resinosa)和班克松(Pinus banksiana)的LAI,发现林分LAI与平均BA相关性很强(R2 = 0.98); Neumann等[12]通过凋落物法估测了加拿大安大略湖南部山杨(Populus grandidentata)-红枫(Acer rubum)混交落叶林的LAI,得到LAI与各树种的BA具有很强的相关性(R2 = 0.994)。这些研究结果表明根据BA能够有效地预测树种或林分的LAI,同时也为基于凋落物法建立LAI与BA的回归关系提供了理论支持。本研究中虽考虑了SLA的季节性差异,但未考虑其是否存在空间变异。而有研究表明利用凋落物法测定LAI时应考虑SLA的空间变异性,如Bouriaud等[19]的研究表明计算LAI时忽略SLA的空间变异能导致8%—24%的误差。因此,为提高凋落物法测定LAI的准确性,SLA的空间变异性在今后研究中应被考虑。
| [1] | Chen J M, Black T A. Defining leaf area index for non-flat leaves. Plant, Cell & Environment, 1992, 15(4): 421-429. |
| [2] | Asner G P, Scurlock J M O, A Hicke J. Global synthesis of leaf area index observations: implications for ecological and remote sensing studies. Global Ecology and Biogeography, 2003, 12(3): 191-205. |
| [3] | Saigusa N, Yamamoto S, Murayama S, Kondo H. Inter-annual variability of carbon budget components in an AsiaFlux forest site estimated by long-term flux measurements. Agricultural and Forest Meteorology, 2005, 134(1/4): 4-16. |
| [4] | Chen J M, Rich P M, Gower S T, Norman J M, Plummer S. Leaf area index of boreal forests: theory, techniques, and measurements. Journal of Geophysical Research, 1997, 102(D24): 29429-29443. |
| [5] | Eriksson H, Eklundh L, Hall K, Lindroth A. Estimating LAI in deciduous forest stands. Agricultural and Forest Meteorology, 2005, 129(1/2): 27-37. |
| [6] | Bréda N J J. Ground-based measurements of leaf area index: a review of methods, instruments and current controversies. Journal of Experimental Botany, 2003, 54(392): 2403-2417. |
| [7] | Ishihara M I, Hiura T. Modeling leaf area index from litter collection and tree data in a deciduous broadleaf forest. Agricultural and Forest Meteorology, 2011, 151(7): 1016-1022. |
| [8] | Jonckheere I, Fleck S, Nackaerts K, Muys B, Coppin P, Weiss M, Baret F. Review of methods for in situ leaf area index determination: Part I. Theories, sensors and hemispherical photography. Agricultural and Forest Meteorology, 2004, 121(1/2): 19-35. |
| [9] | Chianucci F, Cutini A. Estimation of canopy properties in deciduous forests with digital hemispherical and cover photography. Agricultural and Forest Meteorology, 2013, 168: 130-139. |
| [10] | Nasahara K N, Muraoka H, Nagai S, Mikami H. Vertical integration of leaf area index in a Japanese deciduous broad-leaved forest. Agricultural and Forest Meteorology, 2008, 148(6/7): 1136-1146. |
| [11] | Cutini A, Matteucci G, Mugnozza G. Estimation of leaf area index with the Li-Cor LAI 2000 in deciduous forests. Forest Ecology and Management, 1998, 105(1/3): 55-65. |
| [12] | Neumann H H, Den Hartog G, Shaw R H. Leaf area measurements based on hemispheric photographs and leaf-litter collection in a deciduous forest during autumn leaf-fall. Agricultural and Forest Meteorology, 1989, 45(3/4): 325-345. |
| [13] | 郭志华, 向洪波, 刘世荣, 李春燕, 赵占轻. 落叶收集法测定叶面积指数的快速取样方法. 生态学报, 2010, 30(5): 1200-1209. |
| [14] | 苏宏新, 白帆, 李广起. 3类典型温带山地森林的叶面积指数的季节动态: 多种监测方法比较. 植物生态学报, 2012, 36(3): 231-242. |
| [15] | 王宝琦, 刘志理, 戚玉娇, 金光泽. 利用不同方法测定红松人工林叶面积指数的季节动态. 生态学报, 2014, 34(8): 1956-1964. |
| [16] | Sprintsin M, Cohen S, Maseyk K, Rotenberg E, Grünzweig J, Karnieli A, Berliner P, Yakir D. Long term and seasonal courses of leaf area index in a semi-arid forest plantation. Agricultural and Forest Meteorology, 2011, 151(5): 565-574. |
| [17] | Guiterman C H, Seymour R S, Weiskittel A R. Long-term thinning effects on the leaf area of Pinus strobus L. as estimated from litterfall and individual-tree allometric models. Forest Science, 2012, 58(1): 85-93. |
| [18] | Majasalmi T, Rautiainen M, Stenberg P, Lukeš P. An assessment of ground reference methods for estimating LAI of boreal forests. Forest Ecology and Management, 2013, 292: 10-18. |
| [19] | Bouriaud O, Soudani K, Bréda N. Leaf area index from litter collection: impact of specific leaf area variability within a beech stand. Canadian Journal of Remote Sensing, 2003, 29(3): 371-380. |
| [20] | Hoch G, Richter A, Körner C. Non-structural carbon compounds in temperate forest trees. Plant, Cell & Environment, 2003, 26(7): 1067-1081. |
| [21] | Kalácska M, Calvo-Alvarado J C, Sánchez-Azofeifa G A. Calibration and assessment of seasonal changes in leaf area index of a tropical dry forest in different stages of succession. Tree Physiology, 2005, 25(6): 733-744. |
| [22] | 李轩然, 刘琪璟, 蔡哲, 马泽清. 千烟洲针叶林的比叶面积及叶面积指数. 植物生态学报, 2007, 31(1): 93-101. |
| [23] | Jonckheere I, Muys B, Coppin P. Allometry and evaluation of in situ optical LAI determination in Scots pine: a case study in Belgium. Tree Physiology, 2005, 25(6): 723-732. |
| [24] | Gower S T, Kucharik C J, Norman J M. Direct and indirect estimation of leaf area index, fAPAR, and net primary production of terrestrial ecosystems. Remote Sensing of Environment, 1999, 70(1): 29-51. |
| [25] | Nouvellon Y, Laclau J P, Epron D, Kinana A, Mabiala A, Roupsard O, Bonnefond J M, Le Maire G, Marsden C, Bontemps J D. Within-stand and seasonal variations of specific leaf area in a clonal Eucalyptus plantation in the Republic of Congo. Forest Ecology and Management, 2010, 259(9): 1796-1807. |
| [26] | Chen J M. Optically-based methods for measuring seasonal variation of leaf area index in boreal conifer stands. Agricultural and Forest Meteorology, 1996, 80(2/4): 135-163. |
| [27] | Liu Z L, Jin G Z, Qi Y J. Estimate of leaf area index in an old-growth mixed broadleaved-korean pine forest in northeastern china. PLoS One, 2012, 7(3): e32155. |
| [28] | 刘志理, 金光泽, 周明. 利用直接法和间接法测定针阔混交林叶面积指数的季节动态. 植物生态学报, 2014, 38(8): 843-856. |
| [29] | Jeroen S, Lieven N, Sebastiaan L. Predicting the spatial distribution of leaf litterfall in a mixed deciduous forest. Forest Science, 2004, 50(6): 836-847. |
| [30] | Gower S T, Norman J M. Rapid estimation of leaf area index in conifer and broad-leaf plantations. Ecology, 1991, 72(5): 1896-1900. |
| [31] | 马泽清, 刘琪璟, 曾慧卿, 李轩然, 陈永瑞, 林耀明, 张时煌, 杨风亭, 汪宏清. 南方人工林叶面积指数的摄影测量. 生态学报, 2008, 28(5): 1971-1980. |
| [32] | Gower S T, Reich P B, Son Y. Canopy dynamics and aboveground production of five tree species with different leaf longevities. Tree Physiology, 1993, 12(4): 327-345. |
| [33] | 赵传燕, 沈卫华, 彭焕华. 祁连山区青海云杉林冠层叶面积指数的反演方法. 植物生态学报, 2009, 33(5): 860-869. |
| [34] | Qi Y J, Jin G Z, Liu Z L. Optical and litter collection methods for measuring leaf area index in an old-growth temperate forest in northeastern China. Journal of Forest Research, 2013, 18(5): 430-439. |
| [35] | 刘志理, 金光泽. 小兴安岭白桦次生林叶面积指数的估测. 生态学报, 2013, 33(8): 2505-2513. |
| [36] | Deblonde G, Penner M, Royer A. Measuring leaf area index with the LI-COR LAI-2000 in pine stands. Ecology, 1994, 75(5): 1507-1511. |