 2015, Vol. 35
2015, Vol. 35文章信息
- 陈颖涵, 林元烧, 郑连明, 曹文清
- CHEN Yinghan, LIN Yuanshao, ZHENG Lianming, CAO Wenqing
- 北部湾北部水母类群集结构与数量分布
- The assemblage and abundance distribution of jellyfish in northern Beibu Gulf
- 生态学报, 2015, 35(10): 3382-3393
- Acta Ecologica Sinica, 2015, 35(10): 3382-3393
- http://dx.doi.org/10.5846/stxb201307011812
-
文章历史
- 收稿日期:2013-07-01
- 网络出版日期:2014-05-30






水母类(jellyfish)作为胶质类浮游动物(gelatinous zooplankton)的一大类群,包括水螅水母类(Hydromedusae)、管水母类(Siphonophora)、立方水母(Cubomedusae)、钵水母类(Scyphomedusae)和栉水母类(Ctenophore)[1],种类多,数量大,是海洋浮游动物的重要类群之一。近年来,水母的爆发已成为全球海洋关注的热点[2, 3, 4, 5],我国东海北部和黄海海区的水母爆发有加剧的趋势[6]。水母食性较广,几乎无选择地摄食一切可获得的浮游生物[7],当其数量剧增时,会对生物群落产生显著影响[8, 9, 10],改变海洋生物群落结构和生态系统的能量传递机制,从以硅藻→甲壳类浮游动物→鱼类转变为以甲藻→原生动物和微型浮游动物→水母为主的传递方式,引起了海洋生态体制的转换[11],部分能量从原来的鱼类传递到水母类,影响了渔业产量和渔业资源的可持续利用,造成经济和生态损失。我国学者已对不同海域的水母类进行了种类组成、数量分布与群落结构[12, 13, 14, 15, 16]等许多相关研究。由于水母在某些半封闭海湾的生态作用尤为突出,是影响浮游动物种群数量和鱼类补充量的重要因子[17],而北部湾是我国东南海域重要的半封闭海湾,其优良的环境和丰富的生物资源是环北部湾社会、经济、人口等发展的重要支撑,因此有必要对北部湾的水母类进行生态学研究。有关北部湾水母类的调查研究。许振祖[18]介绍了海南岛及邻近海域的水螅水母类的种类,包括花水母目、软水母目、硬水母目、筐水母目和管水母目共52 种水母,记录了[19]北部湾花水母亚纲6新种;郭东晖等[20]进行了夏、冬两季北部湾水母类的生态研究,共记录4 类187 种,种类数夏季高于冬季,主要优势种为拟细浅室水母和双生水母,杜飞雁等[21]报道了北部湾67 种栉水母类的种类记录,并记录主辐特古水母(Tregoubovia perradialis sp.nov.)、泡真囊水母(Euphysora vacuola sp.nov.)和波腺侧丝水母(Helgicirrha sinuatus sp.nov.)3新种。本文利用国家“908”专项的调查资料,对该海域水母类的种类组成、数量分布和群集结构特征进行分析,为进一步探讨北部湾生态系统结构与功能提供研究资料。
1 材料与方法本文材料系国家“908”专项北部湾综合调查专项(908-01-ST09)于2006年夏季(7月15日—8月7日)、冬季(12月25日—翌年1月22日)、2007年春季(4月12日—5月1日)和秋季(10月14日—11月15日)在北部湾海域的综合调查结果。为了解北部湾北部的生物群落结构和生态系统功能,上述专项中选取北部海域40个测站(20.0556°—21.4842° N,108.0381°—109.8225° E,图 1)进行专题探讨。浮游动物样品采集与实验室处理均按照《海洋生物生态调查技术规程》[22]进行,30 m以浅海域用浅水Ⅰ型网(孔径0.505 mm,内径50 cm,网长145 cm),30 m以深海域用大型浮游生物网(孔径0.505 mm,内径80 cm,网长280 cm)由底层至表面垂直拖取,样品现场用5%福尔马林溶液固定。从浮游动物样品中挑取水母,按个体计数法在体式解剖镜下鉴定并计数,丰度以单位水体内的个体数(个/m3)表示。

|
| 图 1 北部湾北部浮游生物采样站位(示等深线) Fig.1 Sampling stations of plankton in northern Beibu Gulf(Depth) |
优势度(Y)计算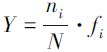
式中,ni为第i种的个体数;fi为该种在各位站出现的频率;N为总的个体数。优势种以Y ≥0.02[23]为优势种划分标准。
聚类分析:将北部湾北部水母类按出现频率降序排列,舍去出现频率<3%的种类,并将各水母丰度进行平方根转换。计算各站位间的Bray-Curtis相似性指数,并以此为基础构建相似性矩阵,在此基础上,用类平均法对站位进行聚类。应用非参数的相似性分析检验各聚类分组的差异显著性。
相似百分比(SIMPER)分析:将北部湾北部水母类的每个种对群集之间非相似性贡献的百分比(累计贡献率取90%)降序排列,确定对北部湾北部水母类群集划分起主要作用的种。
去趋势典范对应(DCCA)分析:对北部湾北部主要水母类(SIMPER分析结果)物种丰度和环境因子进行DCCA分析,数据经对数转换:
Yi=ln(Xi+1)
式中,Xi为水母丰度,Yi为转换后的数值。以了解影响北部湾北部水母类主要种类分布的环境因子。
聚类分析和相似百分比分析采用Primer5.0软件包;去趋势典范对应分析采用Canoco4.5软件包。
本文使用的水深、水温、盐度和叶绿素a等环境因子参数由厦门大学“908”专项办公室提供。
2 结果 2.1 水母群集组成和种数时空分布本区共记录水母类4类125种(包括未定种),以水螅水母类为主(99种),占总种数的79.2%。春季记录水母49种,夏季83种,秋季49种,冬季56种,种数的季节变化与调查区表层平均水温的变化基本一致(图 2)。

|
| 图 2 北部湾北部水母类群集组成与表层温度 Fig.2 Group composition of jellyfish assemblage and surface temperature in northern Beibu Gulf |
本区水母类可分为沿岸暖温性、沿岸暖水性和大洋暖水性3种生态类群,沿岸暖温种很少,只有蝶水母(Ocyropsis crystallina)1种;沿岸暖水种占总种数61.6%,代表种有拟细浅室水母(Lensia subtiloides)、双生水母(Diphyes chamissonis)和球型侧腕水母(Pleurobrachia globosa)等;大洋暖水种占总种数37.6%,代表种有四叶小舌水母(Liriope tetraphylla)、半口壮丽水母(Aglaura hemistoma)和两手筐水母(Solmundella bitentaculata)等。
水母类种数平面分布格局如图 3所示,总体表现为20 m以浅的雷州半岛-广西近岸海域种数较少,20 m以深的远岸海域种数较多,其中夏季水母类各站位种数分布较均匀,基本都在10种以上。

|
| 图 3 北部湾北部水母类种数四季平面分布(示表层温度) Fig.3 Seasonal horizontal distributions of species of jellyfish in northern Beibu Gulf(surface temperature) |
水母类周年平均丰度为61.9 个/m3,其中夏季(24.2 个/m3)>春季(14.3 个/m3)>冬季(13.7 个/m3)>秋季(11.0 个/m3),丰度的季节变化为单峰型。
水母类丰度平面分布的时空变化如图 4所示,调查海域西北部丰度较高,涠洲岛以北海域丰度较低,基本<10.0 个/m3。四季丰度的高值区分布有所不同,夏季和冬季位于调查海域西北部,春季和秋季位于调查海域西部偏中部区域。

|
| 图 4 北部湾北部水母类丰度的四季平面分布(个/m3)(示表层盐度) Fig.4 Seasonal horizontal distributions of abundance of jellyfish in northern Beibu Gulf(surface salinity) |
以优势度≥0.02为依据,水母类四季优势种有拟细浅室水母、双生水母、球型侧腕水母、四叶小舌水母等7种水母,其中管水母类的拟细浅室水母和双生水母为主要优势种。优势种季节性消长现象明显,拟细浅室水母夏季丰度最大,秋季减少,冬季为全年最低;双生水母春季丰度最大,夏季和秋季减少,冬季增加;四叶小舌水母在秋季大量出现,其他季节丰度都比较低(图 5)。

|
| 图 5 北部湾北部水母类主要优势种数量的四季变化 Fig.5 Seasonal changes of abundance of dominant species of jellyfish in northern Beibu Gulf |
拟细浅室水母:春季和夏季数量多,高值区位于调查海域的西北部,其中夏季丰度最大值达78.7 个/m3;丰度低值区位于铁山港海域(<1.0 个/m3)(图 6)。

|
| 图 6 拟细浅室水母丰度的四季平面分布(示表层盐度)(个/m3) Fig.6 Seasonal horizontal distribution of abundance of Lensia subtiloides(Surface Salinity) |
双生水母:春季和冬季数量多,其中春季高值区位于调查海域西偏北部(丰度最大值为36.9 个/m3),琼州海峡西向入海口出现小范围密集区(丰度最大值为36.7 个/m3);冬季密集区位于调查海域西北部(丰度最大值为27.1 个/m3),铁山港海域零星出现(<1.0 个/m3)(图 7)。

|
| 图 7 双生水母丰度的四季平面分布(示表层盐度)(个/m3) Fig.7 Seasonal horizontal distribution of abundance of Diphyes chamissonis(Surface Salinity) |
四叶小舌水母:秋季数量多,在调查海域的分布范围广,密集区位于调查海域的西部,丰度最大值为24.8 个/m3,铁山港海域没有或零星出现(<1.0 个/m3)(图 8)。

|
| 图 8 四叶小舌水母丰度的四季平面分布(示表层盐度)(个/m3) Fig.8 Seasonal horizontal distribution of abundance of Liriope tetraphylla(Surface Salinity) |
此外球型侧腕水母夏季数量最多(114.7 个/m3),密集区位于涠洲岛以北附近海域(丰度最大值为37.9 个/m3)和铁山港海域(丰度最大值为20.8 个/m3),调查海域西南部没有或零星出现(<1.0 个/m3)。春季(22.0 个/m3)、秋季(33.4 个/m3)和冬季(48.1 个/m3)数量则相对较少。
2.4 群集结构选取重要环境因子(深度(D,m)、表层水温(Ts,℃)、中层水温(Tm,℃)、底层水温(Tb,℃)、表层盐度(Ss)、中层盐度(Sm)、底层盐度(Sb)、表层叶绿素a浓度(Chas,mg/m3)、中层叶绿素a浓度(Cham,mg/m3) 和底层叶绿素a浓度(Chab,mg/m3) 共10个因子)分析它们对水母类的群集结构的影响。四季各群集的主要种和特有种如表 1所示。
| 群集 Assemblages 季节Seasons |
Ⅰ | Ⅱ | Ⅲ | |||
| 主要种 Main species |
特有种 Unique species |
主要种 Main species |
特有种 Unique species |
主要种 Main species |
特有种 Unique species |
|
| 春季 Spring |
Solmundella bitentaculata Euphysora knides Voragonema pedunculata |
Diphyes chamissonis Lensia subtiloides Aglaura hemistoma Euphysora knides Nanomia bijuga |
Diphyes chamissonis Aglaura hemistoma Pleurobrachia globosa Nanomia bijuga Lensia subtilis Bassia bassensis Clytia folleata |
|||
| 夏季 Summer |
Pleurobrachia globosa Diphyes subtiloides Eirene ceylonensis Phialucium benga |
Nanomia bijuga Diphyes chamissonis Solmundella bitentaculata |
Lensia subtiloides Diphyes chamissonis Cunina peregrina Liriope tetraphylla Aglaura hemistoma |
Cunina peregrina | ||
| 秋季 Autumn |
Autumn Liriope tetraphylla | Liriope tetraphylla Lensia subtiloides Diphyes chamissonis Pleurobrachia globosa |
Lensia subtiloides Diphyes chamissonis Pleurobrachia globosa Cunina peregrina Enneagonum hyalinum Nanomia bijuga |
|||
| 冬季 Winter |
Clytia folleata | Diphyes chamissonis Aglaura hemistoma Lensia subtiloides Pleurobrachia globosa Nanomia bijuga Euphysora verrucosa |
Diphyes chamissonis Aglaura hemistoma Lensia subtiloides Pleurobrachia globosa Nanomia bijuga Euphysora verrucosa Liriope tetraphylla Solmundella bitentaculata |
|||
春季:群集Ⅰ包括广西沿岸的B05、B21和B40测站,群集Ⅱ分布在远岸海域(图 9)。Anosim分析结果表明,群集间的差异极显著(R=0.628,P=0.001)。

|
| 图 9 北部湾北部水母类丰度-站位聚类及群落结构的划分 Fig.9 Non-metric multidimensional scaling of dominant species and Community Classification of jellyfish in northern Beibu Gulf |
按对相似性贡献大小(以下同),SIMPER分析表明,群集Ⅰ的主要种依次为两手筐水母、刺胞真囊水母(Euphysora knides)和太阳水母(Voragonema pedunculata),其中刺胞真囊水母和太阳水母基本只分布在群集 Ⅰ,位于DCCA排序图(图 10)低温低盐区域,说明该群集受沿岸流影响;群集Ⅰ的主要种为双生水母等5种

|
| 图 10 北部湾北部水母类的主要种类-重要环境因子DCCA排序(累计贡献率取90%) Fig.10 DCCA based on dominant species of jellyfish and environment variables in northern Beibu Gulf |
水母,其中双生水母和球型侧腕水母等7种水母为该群集的特有种,在图中低温低盐和高温高盐区域均有分布,说明该群集受沿岸流和外海水共同影响的混合性特点。此外双生水母离原点较近,说明其生态适应性较广。
夏季:群集Ⅰ分布在雷州半岛-广西沿岸海域,群集Ⅱ包括B26、B30和J10测站,群集Ⅲ分布在远岸海域(图 9)。Anosim分析结果表明群集间的差异极显著(R=0.596,P=0.001)。
SIMPER分析表明,群集Ⅰ的主要种为球型侧腕水母、拟细浅室水母、锡兰和平水母(Eirene ceylonensis)和短腺和平水母(Phialucium benga),其中锡兰和平水母基本只分布在群集Ⅰ,位于DCCA排序图(图 10)低温低盐区域,说明该群集受沿岸流影响;群集Ⅱ的主要种为双小水母(Nanomia bijuga)、双生水母和两手筐水母,其中长囊无棱水母(Sulculeolaria chuni)和双小水母基本只分布在群集Ⅱ,位于图中高温高盐区域,说明该群集主要受外海水影响;群集Ⅲ的主要种为拟细浅室水母、双生水母、异摇篮水母(Cunina peregrina)、四叶小舌水母和半口壮丽水母,其中异摇篮水母为该群集的特有种,位于图中高温高盐区域,表明该群集所在海区受外海水的影响特征。
秋季:群集Ⅰ包括广西沿岸的B21和B38测站,群集Ⅱ几乎分布于整个调查海域(图 9),Anosim分析结果表明两群集间的差异极显著(R=0.764,P=0.001)。
SIMPER分析表明群集I的主要种为四叶小舌水母,短腺和平水母和细颈和平水母(Eirene menoni)基本只分布在群集Ⅰ,位于DCCA排序图(图 10)低盐区域,说明该群集主要受沿岸流影响;群集Ⅱ的主要种为四叶小舌水母、拟细浅室水母、双生水母和球型侧腕水母,其中拟细浅室水母、双生水母、球型侧腕水母、异摇篮水母、晶莹九角水母(Enneagonum hyalinum)和双小水母为群集Ⅱ的特有种,在图中高温高盐和低温低盐区域均有分布,反映了该群集受沿岸流和外海水共同影响的混合性特点。
冬季:群集I包括B36和B40站位,群集Ⅱ分为涠洲岛以北的沿岸群集Ⅱ1和涠洲岛以南的远岸群集Ⅱ2(图 9),Anosim分析表明两群集间的差异极显著(R=0.819,P=0.001),其中群集Ⅱ1和Ⅱ2之间的差异极显著(R=0.771,P=0.001)。
SIMPER分析表明,群集Ⅰ的主要种为单囊美螅水母(Clytia folleata),此外刺胞真囊水母基本只分布在群集Ⅰ,属于近岸种,该群集受沿岸流影响;群集Ⅱ的主要种为双生水母和半口壮丽水母等6种水母,该6种水母和四叶小舌水母、两手筐水母为该群集的特有种,属近岸种或大洋种,群集Ⅱ受沿岸流和外海水的共同影响。对比群集Ⅱ1与Ⅱ2,半口壮丽水母和疣真囊水母(Euphysora verrucosa)只分布在Ⅱ2,位于DCCA排序图(图 10)中高温高盐区域,说明群集Ⅱ2所在海区受外海水影响。
3 讨论 3.1 水母类种类组成特点共记录水母4类125种,其中水螅水母类99种,管水母类17种,钵水母类4种,栉水母类5种。水螅水母类是一类以近岸浅海性为主的浮游动物[24],浅海陆架区的环境条件适合水螅水母的繁育[24],本研究海域调查站位水深均在60 m以浅,因此本区水螅水母类较多,与整个北部湾[25](148种)相比相差不大;管水母类大多属热带大洋性种类,而北部湾终年受南海次表层暖流的影响,湾南部的环境条件更适合管水母的繁育,北部湾北部管水母类种数占整个北部湾[25]管水母类种数的28.8%。
本区水母类可分为沿岸暖温性、沿岸暖水性和大洋暖水性3种生态类群。
沿岸暖温类群 以低温低盐为特征,分布在温度17—20 ℃的低盐近岸水域,本区只记录蝶水母1种,于冬季出现在防城港附近的B21站。
沿岸暖水类群 以高温低盐为特征,分布在温度在20—25 ℃的低盐沿岸水和混合水海域,以水螅水母类为主,共77种。水螅水母类的代表种有端粗范式水母(Vannuccia forbesii)、脊状镰螅水母(Zanclea costata )和锡兰和平水母等;栉水母类的代表种为球型侧腕水母;管水母类的代表种有拟细浅室水母和双生水母。该类群对本调查海域水母类总丰度的变化起主导作用,在调查海区占绝对优势。
大洋暖水类群 以高温高盐为特征,分布在温度>25 ℃的高盐外海水流经海域,共47种。其中分布较广、数量较多的包括四叶小舌水母、半口壮丽水母、两手筐水母和异摇篮水母,主要出现于混合水区或沿岸水边界的混合水区,这与台湾海峡西南部海域的研究结果相似[26]。林茂[27]认为四叶小舌水母和半口壮丽水母对暖流余脉有指示作用。异双生水母(Diphyes dispar)只出现在夏季盐度>34涠洲岛附近的B29站位,为大洋暖水狭布种,可作为高温高盐的南海外海水入侵的指示种。
3.2 水母类不同生态类群的演替本调查海域位于北部湾北部,北部湾为亚热带海湾,湾内有沿岸流和外海水2个水系。沿岸流包括广西沿岸流和越南沿岸流[28],居于调查海域沿岸一带;外海水为南海暖流余脉,居于海南岛西南侧或近底层。沿岸流和外海水的混合区形成混合水,混合水的范围取决于两股水系的消长程度[26]。
水母类种类组成与不同生态类型的演替与水系的消长密切相关。春季大洋暖水性生态类群分布范围在21.00°N以南的外海水海域;夏季由于西南季风的影响,外海水向北推移,21.30°N以南的大部分海区主要受外海水控制,形成混合水区,大洋暖水种的种类数量不断增加,主要分布在混合水区或混合水区的靠岸边界;秋冬季东北季风逐渐强盛,外海水开始向东南方向退回,大洋暖水种数量减少。沿岸暖水性生态类群的演替主要取决于水温的变化。冬季表层水温由远岸向近岸逐渐降低,21.30°N以北的近岸水域表层水温低于20 ℃,出现沿岸暖温性种类;夏、秋两季整个调查海域处于高温期,沿岸暖水性类群种数显著增加,分布于近岸和混合水区。
3.3 水母类丰度的平面分布特点及优势种生态适应性拟细浅室水母、双生水母和四叶小舌水母四季的数量变化决定了北部湾北部水母类的数量变化,水母类丰度的时空分布特征与主要优势种的生态适应性密切相关。
拟细浅室水母为沿岸暖水种,主要分布于沿岸和近海低盐水域[29, 30],是调查海域最主要的优势种,春季和夏季数量多,在夏季形成高峰,密集中心位于调查海域西北部(30.3—31.3 ℃),夏季水母类的丰度分布与该种分布基本一致;冬季水温偏低,拟细浅室水母数量明显减少。
双生水母为沿岸暖水种,是我国东南近海的优势种,除渤海之外,在我国近海均有分布[29]。优势度仅次于拟细浅室水母,数量冬季和春季占绝对优势,密集中心位于调查海域西北部(17.9—23.2 ℃)和琼州海峡西向入海口(22.9—23.60 ℃),冬季和春季水母类的丰度与该种分布基本一致;夏季和秋季水温高,双生水母数量明显减少。与拟细浅室水母相比,双生水母的适温范围较广。
四叶小舌水母为大洋暖水种[31],是秋季的绝对优势种,密集中心位于调查海域西部(盐度31.8—32.3),秋季水母类的丰度与该种分布基本一致。该种在我国从黄海至南海均有分布,是东海[31]、厦门港[32]、台湾海峡[26]和南海北部[25]水螅水母类的优势种。许振祖[26]表明,四叶小舌水母具有广盐性,主要分布于混合水区或沿岸水边界的混合水区。秋季北部湾北部表层盐度30.4—33.2,平均盐度31.3,调查海域混合区范围为全年最广,为四叶小舌水母的生长提供了良好的条件。
3.4 水母类群集结构特点综合聚类分析和DCCA分析的结果,北部湾北部水母类可分为等深线20 m以浅的低盐群集和等深线20 m以深的高盐群集,群集间有显著差异。低盐群集主要受广西-越南沿岸流的影响,高盐群集受沿岸流和外海水的共同影响,两群集的盐度分界基本在32.5左右。
低盐群集:主要分布于沿岸水域,其出现和数量变化主要受沿岸水的影响,根据水温的不同[29],可分为沿岸暖温生态类群和沿岸暖水生态类型。低盐群集的代表种为沿岸暖水生态类型中管水母类的拟细浅室水母和双生水母,徐兆礼[30]认为这两种水母的分布可作为沿岸水影响的指标。该群集对本调查海域水母类的丰度变化起主导作用。
高盐群集:主要分布于外海水流经的区域(包括混合水区),其出现和数量变化主要受外海水的影响,秋季该群集分布于调查区的绝大部分海域,混合水区范围全年最大。代表种为四叶小舌水母和半口壮丽水母。林茂[27]认为这两种水母对暖流余脉有指示作用。此外巴斯水母、双小水母和长囊无棱水母均属于该类群。林茂[33]表明这3种水母是南海中部管水母类的常见种,张金标[34]指出巴斯水母和长囊无棱水母是受外海暖水影响的共有种。本次调查巴斯水母、双小水母和长囊无棱水母是高盐群集的特征种,可以认为对南海暖流进入北部湾有指示作用。
| [1] | Steele J H, Thorpe S A, Turekian K K. Marine Biology: A Derivative of the Encyclopedia of Ocean Sciences. 2nd ed. Burlington: Academic Press, 2009. |
| [2] | Graham W M, Pagès F, Hamner W M. A physical context for gelatinous zooplankton aggregations: a review. Hydrobiology, 2001, 155(1/3): 199-212. |
| [3] | Mills C E. Jellyfish blooms: are populations increasing globally in response to changing ocean conditions? Hydrobiologia, 2001, 451(1/3): 55-68. |
| [4] | Uye S. Blooms of the giant jellyfish Nemopilema nomurai: a threat to the fisheries sustainability of the East Asian Marginal Seas. Plankton and Benthos Research, 2008, 3(S1): 125-131. |
| [5] | 张芳, 孙松, 李超伦. 海洋水母类生态学研究进展. 自然科学进展, 2009, 19(2): 121-130. |
| [6] | 程家骅, 李圣法, 丁峰元, 严利平. 东、黄海大型水母暴发现象及其可能成因浅析. 现代渔业信息, 2004, 19(5): 10-12. |
| [7] | 马喜平, 凡守军. 水母类在海洋食物网中的作用. 海洋科学, 1998, (2):38-41. |
| [8] | Greve W. The 1989 German Bight invasion of Muggiaea atlantica. ICES Journal of Marine Science, 1994, 51(4): 355-358. |
| [9] | Purcell J E, Grover J J. Predation and food limitation as causes of mortality in larval herring at a spawning ground in British Columbia. Marine Ecology Progress Series, 1990, 59: 55-61. |
| [10] | Schneider G, Behrends G. Top-down control in a neritic plankton system by Aurelia aurita medusae-a summary. Ophelia, 1998, 48(2): 71-82. |
| [11] | 单秀娟, 庄志猛, 金显仕, 戴芳群. 长江口及其邻近水域大型水母资源量动态变化对渔业资源结构的影响. 应用生态学报, 2011, 22(12): 3321-3328. |
| [12] | 黄加祺, 许振祖. 东山岛周围海域各类水母的分布. 台湾海峡, 1996, 15(4): 363-367. |
| [13] | 廖秀丽, 李纯厚, 杜飞雁, 林钦, 贾晓平. 2003-2005年大亚湾水螅水母类生态研究. 海洋环境科学, 2006, 25(S1): 48-52. |
| [14] | 马喜平, 高尚武. 渤海水母类生态的初步研究——种类组成、数量分布与季节变化. 生态学报, 2000, 20(4): 533-540. |
| [15] | 王真良. 黄海区水母类的生态研究. 黄渤海海洋, 1996, 14(1): 41-50. |
| [16] | 张芳, 杨波, 张光涛. 胶州湾水母类生态研究 Ⅱ. 优势种丰度的时空分布. 海洋与湖沼, 2005, 36(6): 518-526. |
| [17] | 马喜平, 孙松, 高尚武. 胶州湾水母类生态的初步研究 Ⅰ. 群落结构及其年季变化. 海洋科学集刊, 2000, 42: 91-99. |
| [18] | 许振祖. 海南岛及鄰近海区浮游动物的调查研究——Ⅰ. 水螅水母类. 厦门大学学报: 自然科学版, 1965, 12(1): 90-110. |
| [19] | 许振祖. 北部湾花水母亚纲六新种记述//胡建宇, 杨圣云. 北部湾海洋科学研究论文集(第 1 辑). 北京: 海洋出版社, 2008: 222-229. |
| [20] | 郭东晖, 黄加祺, 许振祖. 北部湾夏、冬两季浮游动物生态学研究Ⅱ 水母类//胡建宇, 杨圣云. 北部湾海洋科学研究论文集(第 1 辑). 北京: 海洋出版社, 2008: 222-229. |
| [21] | 杜飞雁, 许振祖, 黄加祺, 郭东晖. 中国南海北部北部湾水母类调查及三新种记述 (刺胞动物门). 动物分类学报, 2012, 37(3): 506-519. |
| [22] | 梁玉波, 宁修仁, 王长云. 海洋生物生态调查技术规程. 北京: 海洋出版社, 2006. |
| [23] | 徐兆礼, 陈亚瞿. 东黄海秋季浮游动物优势种聚集强度与鲐鲹渔场的关系. 生态学杂志, 1989, 8(4): 13-15. |
| [24] | 陈洪举, 刘光兴. 夏季长江口及邻近海域水母类生态特征研究. 海洋科学, 2010, 34(4): 17-24. |
| [25] | 郑连明. 北部湾水母类生态学研究及中国近海和平水母科分子系统分析 [D]. 厦门: 厦门大学, 2008. |
| [26] | 许振祖. 台湾海峡西南部水螅水母的生态研究. 海洋学报, 1983, 5(1): 91-101. |
| [27] | 林茂, 张金标. 台湾海峡西部海域水螅水母类和栉水母类的生态研究. 海洋学报, 1989, 11(5): 621-628. |
| [28] | 陈波. 北部湾水系形成及其性质的初步探讨. 广西科学院学报, 1986, 2(2): 92-95. |
| [29] | 沈国英, 施并章. 海洋生态学. 北京: 科学出版社, 2002. |
| [30] | 徐兆礼, 张金标, 蒋玫. 东海管水母类生态研究. 水产学报, 2003, 27(S1): 82-90. |
| [31] | 徐兆礼, 张金标, 王云龙. 东海水螅水母类生态研究. 水产学报, 2003, 27(S1): 91-97. |
| [32] | 林茂, 张金标. 厦门港及邻近海域水螅水母类、管水母类和栉水母类的生态研究. 海洋学报, 1989, 11(4): 493-500. |
| [33] | 林茂. 南海中部管水母类生态的初步研究. 海洋学报, 1992, 14(2): 99-105. |
| [34] | 张金标, 林茂. 南海管水母类的生态地理学研究. 海洋学报, 1997, 19(4): 121-131. |