 2015, Vol. 35
2015, Vol. 35文章信息
- 齐麟, 赵福强
- QI Lin, ZHAO Fuqiang
- 不同采伐强度对阔叶红松林主要树种空间分布格局和物种空间关联性的影响
- Impact of logging intensity on the spatial distribution and association of dominant tree species in a broadleaved Korean pine mixed forest
- 生态学报, 2015, 35(1): 46-55
- Acta Ecologica Sinica, 2015, 35(1): 46-55
- http://dx.doi.org/10.5846/stxb201405311129
-
文章历史
- 收稿日期:2014-05-31
- 修订日期:2014-11-14






森林采伐作为人类对森林生态系统最大的干扰活动显著的影响着森林生态系统的结构和功能[1, 2, 3]。采伐的影响不仅仅是对一些具有经济价值的树种进行移除,它还对存留的林木的生长、林下植被动态以及土壤产生衍生的影响[4, 5, 6]。因此,不同的干扰方式能够从很多方面影响植被的恢复,其中最重要的就是改变群落的组成和结构。由于采伐干扰往往对森林的更新和重建过程产生影响,并且干扰体系(包括采伐强度、次数以及采伐轮回期)能够驱动森林生态系统转向一个新的平衡状态,因而,干扰的强度和干扰后的物种格局在次生林的演替速度与方向上起决定性作用[7]。一项研究表明:在采伐40a后,尽管物种丰富度可以恢复到老龄林的水平,但幼树和小树的多度随着采伐强度的增加而逐渐增大,物种组成也随着采伐强度发生变化[8]。这可能是因为采伐后留下的林隙能够为新个体的成功定植和生长提供更多的生态位。采伐强度会对森林群落的恢复速度产生重大的影响。因此,研究不同采伐强度对采伐干扰对森林群落结构的影响有助于揭示采伐干扰如何影响森林群落,也有助于探讨干扰对森林群落组配过程的影响。
近年来,利用分析物种的空间格局在研究森林群落形成的生态学过程 (如物种扩散、种内和种间竞争、干扰、环境异质性等)、种群的生物学特性及其与环境因子之间的相互关系[9, 10, 11, 12]等方面取得了许多进展。但这些研究大多数都设置在未受干扰的原始森林中,旨在了解自然状态下,森林群落中的物种空间分布格局和物种之间的空间关联性[13, 14, 15],对于干扰事件对森林生态系统中物种的空间分布、种间关系影响的研究却相对缺乏。尤其是经过采伐这样的人为干扰以后,由于采伐前在森林群落中占据绝对优势的大径级冠层乔木被全部或部分去除,采伐后的物种组成、种群的年龄结构以及保留林木个体空间格局都发生了较大的变化,这些变化不仅会改变物种、个体之间的关系,还会影响到整个群落的动态和生态系统的演替速度和方向。因此,开展此方面研究,不仅可以明确森林群落中物种空间分布和种间关系对干扰的响应,更有助于加深我们对干扰后森林恢复、演替过程中,森林群落结构、种间关系变化过程的理解。
深入了解干扰对森林群落中种间关系、结构动态的影响对森林生态系统的经营管理具有重要意义[16]。它可以为制定合理的森林采伐、抚育方案,提高森林经营管理的可持续性提供可靠的科学依据[17, 18]。东北林区是我国重要的木材产区,森林蓄积量占全国森林总蓄积量的1/3[19]。该区的地带性植被为阔叶红松(Pinus koraiensis)林。该植被类型不仅生物量巨大,生物多样性也很高,是我国东北重要的生态屏障[19]。该区的森林经营活动频繁,经历了多年的不合理利用,森林质量下降严重[20]。为了保护阔叶红松林这一宝贵的资源,自天保工程实施,该区全面禁止皆伐,调减采伐量,对红松等珍稀濒危物种也实施了禁伐的措施。许多学者针对择伐的采伐强度对森林群落的物种多样性、森林蓄积量的影响开展了大量研究,取得了大量的相关成果[21, 22, 23]。然而,阔叶红松林中主要树种如红松、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandschurica)等的天然更新状况依旧不容乐观[17]。关于不同采伐强度对森林结构动态、种间关系以及更新的影响等方面的研究亟待开展。为了明确上述问题,本研究利用空间点格局分析的方法,对经历不同采伐强度的阔叶红松林中主要树种的空间分布、种间关系以及更新状况进行了研究,以期加深我们对干扰后森林群落结构变化的驱动机理的理解,为制定合理的森林经营方案提供可靠的理论依据。
1 研究区概况与研究方法 1.1 研究区概况研究地点位于吉林省白山市抚松县露水河林业局东升林场。该区位于长白山西北麓,与长白山自然保护区毗邻。海拔在450—1400 m,森林覆盖率达94%。该区属于北温带大陆性气候,是吉林省最寒冷的地区。春季昼夜温差大;夏季短,温热多雨;秋季凉爽,多晴天;冬季长,干燥寒冷。该区年平均气温 4. 6℃,年平均降水量883. 4 mm,年日照时数2259 h,无霜期140d。土壤为火山母质形成的暗棕壤,土层深厚,湿润。地带性顶极植物群落为阔叶红松林,属于长白山植物区系,主要乔木树种有红松、鱼鳞云杉(Piceajezoensis Carr. var. m icrosperma)、山杨( Populus davidiana)、山槐(Albizia kalkora)、黄檗(Phellodendron amurense)、白桦(Betula platyphylla)、暴马丁香(Syringa reticulata)、千金榆(Carpinus cordata)、色木槭 (Acermono)、青楷槭 (Acer tegmentosum)、水曲柳、胡桃楸(Juglans mandshurica)、紫椴 (Tilia amurensis)、冷杉 (Abies nephrolepis)、春榆(Ulmus davidiana Planch var. japonica)、蒙古栎等。灌木主要有毛榛子(Corylus fargesii)、东北山梅花(Philadelphus schrenkii)、忍冬 (Lonicera japonica)等,草本以山茄子(Brachybotrys paridiformis)、木贼(Equisetum hyemale)等为主,多样性丰富。
1.2 样地设置研究采用典型取样方法,选择海拔、坡向、坡度相近,地势平坦,经不同采伐方式以及强度采伐,并自然恢复30a左右且采伐前林型为原始阔叶红松林的样地3块,并选择一块未经采伐的原始阔叶红松林作为对照。在每块样地中选择一块地势平坦、面积为 1 hm2 (100 m ×100 m)的正方形样方。样方设置如表 1所示。
| 样地 Sites | 采伐前森林类型 The forest type before | 当前森林类型 The forest type now | 采伐强度/% Selective logging | 采伐时间 Selective logging time |
| CON | 原始阔叶红松林 | 原始阔叶红松林 | 0 | |
| LSC | 原始阔叶红松林 | 阔叶红松择伐林 | 15 | 1980′s |
| MSC | 原始阔叶红松林 | 阔叶红松择伐林 | 30 | 1980′s |
| CC | 原始阔叶红松林 | 白桦林 | 皆伐 | 1980′s |
在每块样方内 ,对所有胸径 ≥2 cm的乔木进行编号、挂牌、记录树种、测量胸径、目测树高、并以样地左下角为坐标原点测量所有胸径 ≥2 cm的乔木相对于坐标原点的平面直角坐标系的相对坐标(以 X,Y形式表示 )。
将胸径在2—8cm且树高大于1.5m的个体定义为幼树,胸径在8—20cm的个体定义为中等径级树(简称中树)。
1.4 乔木个体空间点格局分析本研究应用Ripley′s K函数分析不同物种、径级个体的空间分布格局与种间空间关联性。利用Kest函数分析不同物种在采伐后的空间分布格局的变化;利用Kmulti函数来分析不同物种、不同径级个体空间关联性。为了便于横向比较,将此函数做了如下转化:
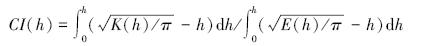
式中,CI为聚集指数;K为K函数(Kest或Kmulti);h为距离; 当CI>1时,表示该物种呈显著聚集分布,或者表示两个类群的空间相关性为正,即两物种相吸引; 当1<CI<1时,表示该物种为随机分布,或者两个类群无空间相关性; 当CI<-1时,表示该物种为规则分布,或者两个类群相排斥,即两个物种是竞争关系。
空间点格局分析由R语言中相关的程序包完成。
2 研究结果 2.1 采伐对阔叶红松林乔木群落结构的影响在未经采伐的原始阔叶红松林(CON)中,成年的(DBH>20 cm)红松(101株/hm2)、蒙古栎(24株/hm2)以及水曲柳(11株/hm2)和紫椴(41株/hm2)的密度较高,特别是红松,其密度占主林层乔木密度的50%以上。在林下层(DBH<8cm),暴马丁香(66株/hm2)、千金榆(120株/hm2)、花楷槭(58株/hm2)、青楷槭(96株/hm2)以及色木槭(81株/hm2)和假色槭(177株/hm2)的幼树占林下层全部物种密度的80%以上。
主林层中占优势的乔木物种(红松、紫椴、水曲柳、蒙古栎),除紫椴外,其余各物种幼树的密度极低,更新情况较差(图 1-COM)。轻度择伐(采伐强度15%)对阔叶红松林乔木树种总体的密度影响不大,但减少了蒙古栎、紫椴等用材树种成年树的密度(蒙古栎7株/hm2;紫椴17株/hm2)。白桦幼树,槭树类的幼树总和明显高于原始阔叶红松林。主要乔木树种的更新情况与原始阔叶红松林相似(图 1-LSC)。在中等强度择伐样地(MSC)中,各种径级的白桦都高于轻度择伐样地。红松、水曲柳、蒙古栎以及紫椴的成年树密度低于强度择伐样地,而这些树种的幼树密度高于低强度择伐样地,更新情况得到明显改善(图 1-MSC)。在皆伐样地(CC)中,在采伐前占据绝对优势的乔木物种都已被采伐,速生物种白桦取代了原有的森林。槭树类等喜阴的物种的密度低于择伐后的次生阔叶红松林。红松、蒙古栎、水曲柳、紫椴等主要乔木树种的幼树密度明显高于择伐后的次生阔叶红松林(图 1-CC)。

|
| 图 1 不同择伐强度样地中各树种年龄结构 Fig. 1 Age structure of 4 sites following different selective logging intensity CON:原始阔叶红松林;LSC:低强度择伐阔叶红松林(15%采伐强度);MSC: 中等强度择伐后阔叶红松林(30%采伐强度);CC: 皆伐后自然恢复白桦林 |
综上所述,采伐减少了冠层主要乔木成年个体的密度,且随着采伐强度的增加,影响越明显。在一定程度上,采伐促进了一些物种,如槭树类、白桦等。但一些树木如红松蒙古栎等只有在皆伐样地中幼树密度有明显增加。
2.2 不同采伐强度对阔叶红松林主要物种空间分布格局的影响采伐后,各物种个体在样地中空间格局发生了明显的变化,且不同的采伐强度所造成的影响也不同(图 2)。 在原始阔叶红松林中,假色槭、青楷槭、白桦在0—25m的尺度上呈显著的聚集分布;紫蒙古栎在5—25m的尺度上表现为显著的聚集分布,但聚集程度较低;色木槭在0—7.5m的尺度内表现为较弱的;蒙古栎在5—25m的尺度内表现为聚集分布,聚集程度随着尺度的增加而降低。水曲柳、红松以及春榆在0—25m尺度上表现为随机分布;蒙古栎在0—5m尺度上表现为随机分布;色木槭在7.5—25m的尺度上为随机分布。低强度的择伐没有明显改变阔叶红松林中主要乔木树种的空间分布格局。各物种的空间分布格局与未受采伐的原始阔叶红松林中的相似(图 3)。中等强度的择伐,增加了假色槭在0—7.5m的聚集性,降低了其在7.5—25m尺度的聚集性;色木槭的空间分布格局有随机分布变为显著的聚集分布;红松、春榆、水曲柳的空间分布格局变为聚集程度较低的显著聚集性分布;蒙古栎空间分布格局变为随机分布;中等强度择伐还增加了紫椴空间分布格局的聚集程度(图 3)。在皆伐后的样地中,假色槭的空间分布格局虽然依然是聚集分布,但在5—25m尺度内聚集程度显著低于其他3块样地;青楷槭的空间分布格局的聚集性增加,显著高于其他3块样地。水曲柳、蒙古栎、红松、春榆的空间分布聚集程度显著增加(图 3)。

|
| 图 2 不同采伐强度样地中主要乔木物种的空间分布 Fig. 2 Spatial distribution pattern of major tree species in broadleaved Korean pine mixed forest following selective logging CON:原始阔叶红松林;LSC:低强度择伐阔叶红松林(15%采伐强度);MSC: 中等强度择伐后阔叶红松林(30%采伐强度);CC: 皆伐后自然恢复白桦林 |

|
| 图 3 不同采伐强度后阔叶红松林主要树种空间分布 Fig. 3 Effect of logging intensity on tree spatial distribution DI: 空间分布指数;CON:原始阔叶红松林;LSC:低强度择伐阔叶红松林(15%采伐强度);MSC: 中等强度择伐后阔叶红松林(30%采伐强度);CC: 皆伐后自然恢复白桦林;ENV-UP: 上包迹线;ENV-LOW:下包迹线; 当模拟曲线在上包迹线上方时该物种为显著的聚集分布,在下包迹线下方是为显著的规则分布,在包迹线之间为随机分布 |
在未经采伐的原始阔叶红松林中,成年白桦与紫椴幼树的空间关联性在10—25m的尺度上为显著的负相关,与红松、蒙古栎和水曲柳幼树在0—25m尺度内无明显空间相关性;红松成年树与水曲柳幼树在15—25m尺度内呈现显著的负相关,与紫椴幼树在15—25m尺度上呈现显著的正相关,与白桦、蒙古栎在0—25m尺度无空间相关关系;蒙古栎成年树与白桦幼树在5—25m尺度内呈显著的负相关关系,与水曲柳幼树在0—25m尺度内呈正相关关系,但相关性并不高,与红松幼树、紫椴幼树在0—25m尺度内无显著空间相关关系。成年水曲柳和紫椴与白桦、红松、蒙古栎以及紫椴的幼树在0—25m尺度内均无显著空间关联(图 4)。在轻度择伐样地中,成年白桦与紫椴在15—25m尺度内无显著空间相关关系;成年红松与白桦幼树在7.5—22.5m尺度内呈显著负相关关系,与紫椴幼树在0—5m内呈显著正相关关系;成年水曲柳与紫椴幼树在10—17.5m尺度内呈显著正相关关系;其余各物种空间关联性与原始林中相近(图 4)。在中度择伐样地中,成年白桦与红松幼树在10—25m尺度上呈显著的正相关,且相关的程度随尺度的增加而增加,与水曲柳幼树在15—25m尺度内呈显著的负相关关系,与紫椴幼树在0—15m尺度内呈显著负相关关系;成年蒙古栎与白桦幼树在15—25m尺度内显著正相关,与红松幼树在0—25m尺度内呈显著正相关,与水曲柳幼树在15—25m尺度内显著负相关,与紫椴幼树在0—25m尺度内无显著相关关系;水曲柳成年树与红松幼树在10—25m尺度内呈显著负相关;成年紫椴与红松幼树在0—25m尺度内呈显著正相关,与水曲柳幼树在5—25m尺度内呈显著正相关关系(图 4)。在皆伐样地中,成年白桦与红松幼树在5—25m尺度内呈显著正相关,与水曲柳幼树在10—25m尺度内呈显著正相关;与紫椴幼树在0—25m尺度内无显著空间相关性;皆伐样地中无红松成年树;蒙古栎成年树与白桦幼树在0—25m尺度内呈显著的负相关关系,与红松幼树在0—25m尺度内呈显著的负相关关系,与水曲柳幼树在0—20m尺度内呈显著负相关,在20—25m尺度内显著正相关,与紫椴幼树在20—25m尺度内显著正相关;成年水曲柳与白桦幼树在0—25m尺度内无显著相关,与红松幼树在0—10m尺度显著正相关,在20—25m尺度显著负相关;紫椴成年树与白桦幼树和红松幼树在0—25m尺度显著正相关与蒙古栎幼树在10—25m尺度内显著正相关,与水曲柳幼树在0—5m尺度显著负相关,在5—25m尺度内显著正相关(图 4)。

|
| 图 4 不同采伐强度对阔叶红松林主要树种种间空间关联性的影响 Fig. 4Effect of logging intensity on spatial association of dominate tree special CON:原始阔叶红松林;LSC:低强度择伐阔叶红松林(15%采伐强度);MSC: 中等强度择伐后阔叶红松林(30%采伐强度);CC: 皆伐后自然恢复白桦林;ENV-UP: 上包迹线;ENV-LOW:下包迹线; 幼树: 2cm<DBH<8cm; 成年树:DBH>20cm;当模拟曲线在上包迹线时俩物种的空间关系为显著的正相关,在下包迹线下方则表示为显著负相关,在包迹线之间为无显著相关性 |
不同强度的采伐还使阔叶红松林主要乔木树种种内空间关联性发生了改变。在原始林中,多数物种的幼树都与同种成年树成正相关或者不相关的关系,如假色槭、青楷槭、水曲柳等。轻度择伐(15%)并没有显著影响各物种个体的种内空间关联性。中等强度的择伐使假色槭的幼树与成年树由原始林中的物空间关联性转变为显著的负相关关系。皆伐对阔叶红松林主要乔木树种的种内空间关联性影响最大。水曲柳、蒙古栎、红松、春榆、紫椴以及白桦在皆伐样地中的种内空间关联性都较其他样地有显著变化(图 5)。

|
| 图 5 不同采伐强度对阔叶红松林主要树种处内空间关联性的影响 Fig. 5 Effect of logging intensity on intraspecific spatial association CI: 空间关联度指数;CON:原始阔叶红松林;LSC:低强度择伐阔叶红松林(15%采伐强度);MSC: 中等强度择伐后阔叶红松林(30%采伐强度);CC: 皆伐后自然恢复白桦林;ENV-UP: 上包迹线;ENV-LOW:下包迹线; 当模拟曲线在上包迹线上方时该物种大树(DBH>20cm)与小树(2cm<DBH<8cm)的空间关联性为显著的正相关,在下包迹线下方则表示为显著负相关关系,在包迹线之间为无显著相关性 |
乔木个体的空间分布格局、空间关联性可以较好的反映个体、物种之间的竞争关系[24, 25]。在本研究中,轻度择伐对阔叶红松林群落(包括:个体密度、物种组成以及个体的空间格局和空间关联性)造成的影响最小。在采伐30a后,次生阔叶红松林的森林群落结构特征已经基本恢复到采伐前的状态。然而,在原始阔叶红松林中,尽管成年的红松、蒙古栎、水曲柳以及紫椴等用材树种占据着绝对优势,但这些树种的幼树在样地中的密度却很低,更新情况很差(图 1)。造成这种格局的可能原因是,在这两类样地中,阔叶红松林中主要乔木树种的成年树中占据着绝大部分的资源(阳光、水、养分等),使得这些树种的幼树难以获得足够的生存空间和资源。轻度择伐林后的阔叶红松林的状况与原始阔叶红松林的类似,表明尽管轻度择伐对原始阔叶红松林的扰动较小,尽管使采伐后的次生阔叶红松林群落结构得以迅速的恢复,但主要乔木树种的更新状况并未得到改善。
中等强度的择伐较为明显的减少了阔叶红松林中主要乔木树种中较大径级(DBH>20cm)个体的密度,林下更新情况得到明显改善。这表明,中等强度的的择伐释放出了较多的被大径级乔木所占据的空间和资源,使其为更新苗所用。有研究表明,森林经过择伐后,尽管幼苗的数量要少于采伐前,但幼苗的成活率显著提高,因此,幼树的密度要高于未采伐过的森林[26]。除此之外,采伐后的次生阔叶红松林的群落结构特征与原始阔叶红松林群落结构特征相差不大。这表明,中等强度的择伐不仅可以维持阔叶红松林群落结构特征,还可以一定程度的促进更新。
皆伐去除了阔叶红松林中所有的乔木树种,使其演替回到初始阶段。先锋物种白桦迅速占据了采伐迹地。在此类样地中,尽管红松、蒙古栎、水曲柳等顶级群落的优势物种的更新情况极好,但森林群落要恢复到采伐前的结构和功能要经历一个完整的演替过程,可能所需要的时间很长。
3.2 采伐物种选择对阔叶红松林群落结构、物种空间关联性的影响除择伐的强度外,择伐时所选择的物种也是影响采伐后阔叶红松林更新、群落结构动态的重要因素[27]。因为,在森林生态系统中,个体之间、物种之间的关系不尽相同,有的呈现出聚集趋势,有的呈排斥趋势,采伐时选择不同的物种,势必会对保留木造成影响。在本研究中,通过采伐后的群落结构可以看出,轻度择伐样地中,主要采伐的是阔叶树种如水曲柳、蒙古栎等,红松则被保留。在此样地中,不仅红松的更新较差,蒙古栎和水曲柳的幼树密度也很低。而在中度择伐样地中,红松、蒙古栎、水曲柳等乔木的成年树密度均显著低于原始阔叶红松林和轻度择伐样地,而这几个物种的幼树却高于它们。这中情况不仅仅与采伐的强度有关,还很可能与是否采伐了红松有关。有研究表明,红松的成年树会对自身的更新和其他物种如水曲柳和蒙古栎的更新产生抑制作用[28]。在本研究中,对红松成年树与各物种幼树的空间关联性分析中也观察到了这种现象。因此,对阔叶树红松林中成年红松密度的调控,会对阔叶红松林群落动态产生重要的影响。然而,研究区现阶段对红松采取禁伐的政策,该政策尽管使红松这一珍稀物种得到最大的保护,但对其种群的动态的长期稳定性可能产生不利的影响。
综上所述,不同的采伐强度会对阔叶红松林的群落结构动态,个体的空间分布格局以及空间关联性产生显著的影响,中等的采伐强度(30%的蓄积量)既可以使阔叶红松林的群落结构在较短的时间内得到较好的恢复,又可以促进主要乔木树种的更新。择伐时所选择不同的采伐物种会改变群落内个体之间的空间关联性,进而改变群落的结构动态。因此,进行采伐设计时,采伐的强度应控制在30%左右,采伐的轮回期应大于30a。此外,还需要考虑采伐时物种的选择以及物种之间的关系。在研究区,应把成年红松的密度控制在合理的密度内,以促进红松和其他阔叶树种的更新,维持阔叶红松林生态系统结构的动态平衡。
| [1] | 吴蒙蒙, 王立海, 侯红亚, 刘德洲. 采伐强度对阔叶红松林空间结构的影响. 东北林业大学学报, 2013, 41(9): 6-9. |
| [2] | Valverde-Barrantes O J, Rocha O J. Logging impacts on forest structure and seedling dynamics in a Prioria copaifera (Fabaceae) dominated tropical rain forest (Talamanca, Costa Rica). Revista De Biología Tropical, 2014, 62(1): 347-357. |
| [3] | Arbainsyah, de Iongh H H, Kustiawan W, de Snoo G R. Structure, composition and diversity of plant communities in FSC-certified, selectively logged forests of different ages compared to primary rain forest. Biodiversity and Conservation, 2014, 23(10): 2445-2472. |
| [4] | Dulamsuren C, Khishigjargal M, Leuschner C, Hauck M. Response of tree-ring width to climate warming and selective logging in larch forests of the Mongolian Altai. Journal of Plant Ecology, 2013, 7(1): 24-38. |
| [5] | Wang H, Shao G F, Dai L M, Xu D, Xu H Y, Wang F. An impacts of logging operations on understory plants for the broadleaved/Korean pine mixed forest on Changbai mountain, China. Journal of Forestry Research, 2005, 16(1): 27-30. |
| [6] | 杨秀云, 韩有志, 宁鹏, 武小钢. 采伐干扰对华北落叶松林下土壤水分、pH和全氮空间变异的影响. 土壤学报, 2011, 48(2): 356-365. |
| [7] | Burivalova Z, Şekercioglu Ç H, Koh L P. Thresholds of logging intensity to maintain tropical forest biodiversity. Current Biology, 2014, 24(16): 1893-1898. |
| [8] | 齐麟, 代力民, 于大炮, 周莉, 钟磊, 苏东凯. 采伐对长白山阔叶红松林乔木群落的影响. 湖南农业科学, 2009, (8): 129-132. |
| [9] | Fangliang H, Legendre P, LaFrankie J V. Distribution patterns of tree species in a Malaysian tropical rain forest. Journal of Vegetation Science, 1997, 8(1): 105-114. |
| [10] | Druckenbrod D L, Shugart H H, Davies I. Spatial pattern and process in forest stands within the Virginia piedmont. Journal of Vegetation Science, 2005, 16(1): 37-48. |
| [11] | Nathan R. Long-distance dispersal of plants. Science, 2006, 313(5788): 786-788. |
| [12] | Martínez I, Wiegand T, González-Taboada F, Obeso J R. Spatial associations among tree species in a temperate forest community in North-western Spain. Forest Ecology and Management, 2010, 260(4): 456-465. |
| [13] | Condit R, Ashton P S, Baker P, Bunyavejchewin S, Gunatilleke S, Gunatilleke N, Hubbell S P, Foster R B, Itoh A, LaFrankie J V, Lee H S, Losos E, Manokaran N, Sukumar R, Yamakura T. Spatial patterns in the distribution of tropical tree species. Science, 2000, 288(5470): 1414-1418. |
| [14] | 张健, 郝占庆, 宋波, 叶吉, 李步杭, 姚晓琳. 长白山阔叶红松林中红松与紫椴的空间分布格局及其关联性. 应用生态学报, 2007, 18(8): 1681-1687. |
| [15] | Wang X G, Comita L S, Hao Z Q, Davies S J, Ye J, Lin F, Yuan Z Q. Local-scale drivers of tree survival in a temperate forest. PloS One, 2012, 7(2): e29469, doi: 10.1371/journal.pone.0029469. |
| [16] | 叶尔江·拜克吐尔汉, 张春雨, 赵秀海. 天山云杉林优势种空间格局及其空间关联性. 林业资源管理, 2013, (3): 58-62. |
| [17] | 刘宪钊, 陆元昌, 周燕华. 退化次生林恢复过程中群落结构和生态位动态. 生态学杂志, 2010, 29(1): 22-28. |
| [18] | 缪宁, 刘世荣, 史作民, 马姜明, 王晖. 强度干扰后退化森林生态系统中保留木的生态效应研究综述. 生态学报, 2013, 33(13): 3889-3897. |
| [19] | Yu D P, Zhou L, Zhou W M, Ding H, Wang Q W, Wang Y, Wu X Q, Dai L M. Forest management in northeast China: history, problems, and challenges. Environmental Management, 2011, 48(6): 1122-1135. |
| [20] | 屈红军, 牟长城. 东北地区阔叶红松林恢复的相关问题研究. 森林工程, 2008, 24(3): 17-20. |
| [21] | 邱仁辉, 陈涵. 择伐作业对常绿阔叶林乔木层树种结构及物种多样性的影响. 中国生态农业学报, 2005, 13(3): 158-161. |
| [22] | 郑丽凤, 周新年. 择伐强度对天然林树种组成及物种多样性影响动态. 山地学报, 2008, 26(6): 699-706. |
| [23] | 陈辉荣, 周新年, 蔡瑞添, 胡喜生, 巫志龙, 周成军. 天然林不同强度择伐后林分空间结构变化动态. 植物科学学报, 2012, (3): 230-237. |
| [24] | Petritan I C, Marzano R, Petritan A M, Lingua E. Overstory succession in a mixed Quercus petraea-Fagus sylvatica old growth forest revealed through the spatial pattern of competition and mortality. Forest Ecology and Management, 2014, 326: 9-17. |
| [25] | Ledo A, Cañellas I, Barbeito I, Gordo F J, Calama R A, Gea-Izquierdo G. Species coexistence in a mixed Mediterranean pine forest: Spatio-temporal variability in trade-offs between facilitation and competition. Forest Ecology and Management, 2014, 322: 89-97. |
| [26] | Wang G, Liu F. The influence of gap creation on the regeneration of Pinus tabuliformis planted forest and its role in the near-natural cultivation strategy for planted forest management. Forest Ecology and Management, 2011, 262:413-423. |
| [27] | Tatsumi S, Owari T, Kasahara H, Nakagawa Y J. Individual-level analysis of damage to residual trees after single-tree selection harvesting in northern Japanese mixedwood stands. Journal of Forest Research, 2014, 19(4): 369-378. |
| [28] | 徐振邦, 戴洪才, 李昕, 章依平, 郭杏芬, 戴力民, 杨发柱. 主要伴生树种叶粉对紫椴幼苗生长的影响. 植物生态学与地植物学学报, 1993, 17(2): 175-182. |