 2015, Vol. 35
2015, Vol. 35文章信息
- 王晓雨, 于大炮, 周莉, 周旺明, 吴志军, 郭焱, 包也, 孟莹莹, 代力民
- WANG Xiaoyu, YU Dapao, ZHOU Li, ZHOU Wangming, WU Zhijun, GUO Yan, BAO Ye, MENG Yingying, DAI Limin
- 长白山北坡林线岳桦种群空间分布格局
- Spatial patterns of a treeline Betula ermanii Cham. population on the north slope of Changbai Mountain
- 生态学报, 2015, 35(1): 116-124
- Acta Ecologica Sinica, 2015, 35(1): 116-124
- http://dx.doi.org/10.5846/stxb201405221055
-
文章历史
- 收稿日期:2014-05-22
- 修订日期:2014-11-10






2. 中国科学院大学, 北京 100049
2. University of Chinese Academy of Sciences, Beijing 100049, China
高山林线常被认为是全球气候变化的理想监测器[1]。所有自然林线植被具有的趋同特征之一即乔木树种的树高随海拔升高而显著降低[2],低矮灌木逐渐取代乔木生活型占据高山植被带。由直立乔木到低矮灌木的生活型变化,常被认为是林线植物能够适应高海拔胁迫环境的关键。
种群空间格局是指种群内个体的空间分布方式或配置特点,是植物种群在群落中相对位置可以定量化描述的基本特征之一[3]。种群分布格局是植物种群生物学特性对环境条件长期适应和选择的结果[4, 5]。对于高山林线地带不同生活型岳桦之间格局关系的研究,有利于了解该地区种群分布规律及预测未来气候变化下的种群过程演化,以及揭示乔木生活型不能占据高山生命带的机理。
长白山北坡岳桦在海拔1950m左右形成急变型林线[6],种群总体呈斑块状分布于过渡带内,优势树种岳桦在过渡带地区表现出树高生长受限、矮曲生长等特征。以往关于长白山林线岳桦种群格局的研究表明林线岳桦呈聚集分布[7],但该地区更为典型的乔木生活型和矮曲灌木生活型空间分布规律的内容还未见报道。本研究采用点格局分析方法,研究长白山北坡林线岳桦种群总体分布格局,以及不同生活型岳桦的格局特征,探讨可能影响岳桦种群分布的生物学过程。
1 资料与研究方法 1.1 研究区概况研究区位于长白山北坡(41°31′—42°28′N,127°9′—128°55′E)的岳桦苔原交错带。该区气候属于受季风影响的温带大陆性气候,冬季寒冷漫长,夏季温暖多雨而短暂,年平均气温-2.3—-3.8℃,年降雨量967—1400mm,积雪期199—219d。土壤为山地生草森林土。由于地形以及其他形式的胁迫如风、雪等因素的影响,海拔2000m以上仅有岳桦一种乔木形成优势,呈斑块状镶嵌分布于苔原过渡带,并逐渐向上扩展其分布区。
1.2 研究方法 1.2.1 样地设置与植被调查2006年9月,在长白山北坡海拔2000m左右的岳桦苔原过渡带区域建立一块80m×80m的固定样地。对样方内岳桦进行每木调查,具体调查指标包括植株的高度(H)、胸径 (距地表 1.3m处的树干直径,DBH);对于所有高度小于1.3m的植株,测定相应的基径(地面根颈部位的树干直径,D);同时记录每个植株在样地中的坐标位置(x,y)。将上述数据整理为空间点集,每个坐标点同时带有H、DBH或D,以及分枝属性信息。
|
主干个数
Tree No. | 树高等级H/m
Tree height class | 分类结果Class | |||||
| 树高H/m | 地径D/cm | 胸径DBH/cm | 个体数
Number | 所占比例/%
Percent | 分类单元
Class unit | ||
| 表中“—”代表无实际意义或无法计算的指标, s1:具有独立主干且树高大于3m的老树/乔木(n=215); s2:具有独立主干树高介于0.5m和3m之间的中树(n=1646); s3:树高小于0.5m的幼苗(n=101); s4:具有2个以上主干或分枝的灌木生活型(n=921) | |||||||
| n=1 | H>3 | 4.246±1.072 | — | 4.954±3.1 | 215 | 7.46 | s1乔木/老树 |
| n=1 | 0.5<H≤3 | 1.467±0.593 | 1.276±0.455 | 0.9±0.913 | 1646 | 57.09 | s2中树 |
| n=1 | H≤0.5 | 0.452±0.083 | 0.885±0.315 | — | 101 | 3.5 | s3幼苗 |
| n≥2 | — | 2.041±1.221 | 1.441±0.565 | — | 921 | 31.95 | s4灌木型 |
计算足够小的空间区域上点的密度可以实现对岳桦种群总体分布格局的预判。本文采用估计函数为:

Morishita Index(MI)是表示种群聚集程度的指标之一,通常仅用一个MI来描述固定样方内种群的分布格局。而通过连续分割固定样方,得到不同样方大小的MI,可以将空间聚集程度与尺度信息联系在一起。具体方法为将一个大样方逐步分割为2×2,3×3,…,n×n个小样方,通过统计落入样方内的个体数计算一组连续的MI。

本文采用点格局分析方法分析岳桦个体的空间分布。使用的二阶统计量包括:Ripley′K,
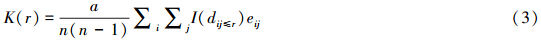
空间点集呈明显的非均匀分布时,适合使用g(r)指数,也称pair correction function或 简称pcf。相对于Ripley′K函数,该估计值的方差相对稳定。

通过Monte-Carlo拟合检验确定置信区间,拟合方法为以完全随机分布的种群为基本模型,拟合一系列随机坐标点,此过程重复99次,得到95%置信区间,即图中上、下包迹线之间的范围。本文所有数据处理、格局分析、统计图制作均通过R软件实现。
2 结果与分析 2.1 种群生活史特征与乔、灌两种生活型样地内共有植株2883棵(表 1),岳桦平均树高为1.822m,最小树高0.1m,最大树高8.50 m,树高总体呈偏峰分布,峰值位于1.3m处(图 1);3个四分位点分别为1.00,1.55、2.20 m。分布频数在1.50—3.00 m范围内迅速下降。除树体矮小外,高山岳桦相对于低海拔岳桦,具有一种独特的生活型即矮曲灌木。对样地内的矮曲灌木进行分枝数的统计(图 2),测得的最大分枝数为23个,其中具有一个明显主干的植株占总株数的68.05%。

|
| 图 1 种群内所有个体树高值的密度分布 Fig. 1 Density distribution of three height of every individual Betula in population |

|
| 图 2 岳桦种群总体在不同尺度下的聚集指数MI Fig. 2 Plot of Morishita Index of increasing diameter of quadrat |
考虑到过渡带地区独特的气候条件下难以通过树龄、或者DBH区分岳桦龄级结构,而树高特征能更好地反应岳桦树体大小,根据林线的最新定义,树高大于3m的植株才能被视为“树”[8]。本研究利用树高结构代替年龄结构,将树高指标和分枝数作为种群生活史阶段与生活型的划分依据,将样地内所有岳桦植株分为s1:具有独立主干且树高大于3m的老树/乔木(n=215);s2:具有独立主干树高介于0.5m和3m之间的中树(n=1646);s3:树高小于0.5m的幼苗(n=101);s4:具有2个以上主干或分枝的灌木生活型(n=921)。

|
| 图 3 岳桦种群4个生活史阶段的空间分布核密度估计 Fig. 3 Kernel estimation of distribution for different life stages-related Betula 颜色越深代表分布密度越大,颜色越浅代表分布密度越小 |
种群总体的MI图(图 4)显示,0—60m范围内,MI始终大于1,则0—60m内的各个尺度,岳桦种群均表现为聚集分布;MI在0—10m的小尺度内MI值迅速减小,10—60m较大尺度上MI有所降低,但基本稳定在1以上,岳桦种群在该尺度上仅表现为低强度的聚集。

|
| 图 4 各生活史阶段的空间格局 Fig. 4 Spatial patterns of different life stages s1: 老树;黑色粗实线代表实际观察值;黑色粗虚线代表完全随机分布条件下的g(r);灰色区域为95%置信区间;s2: 中树,其他同s1;s3: 幼苗;s4:矮曲灌木型 |
核密度估计图表明(图 3)岳桦种群大部分以斑块状镶嵌分布于样地内,但不同生活史阶段的岳桦分布格局仍有差别。
g(r)统计量定量分析4种生活史阶段的岳桦在各个尺度上的格局(图 4),本研究中的分析范围皆为0—20m。老树在0—16m区间内表现出明显的聚集分布,16—18m为随机分布,18—20m为均匀分布;中树在0—20m区间内一直表现为聚集分布,且随尺度增大,聚集趋势逐渐减小;幼苗在0—7m区间内表现为明显的聚集分布,7—19m大致为随机分布,19—20m为均匀分布;矮曲灌木型在0—5m范围内表现为聚集分布,5—20m为随机分布。
几种生活史阶段均在不同尺度上表现出格局差异,且在小尺度上大部分表现为聚集,为了量化4种类型的总体聚集强度g(r),构造一个新的统计量CI,CI=(观察值-理论值)/(置信上限-理论值)。据此得到(图 5),相比于s4、s3,s2、s1表现出更明显的聚集趋势。

|
| 图 5 各生活史阶段的整体聚集趋势 Fig. 5 Clustering trend of different life stages s1: 老树;图中黑色虚线代表-1,1的区间;黑色粗实现代表观察值;灰色粗虚线代表聚集程度较弱;黑色粗实现代表聚集程度较强;s2: 中树;其他同s1;s3: 幼苗;s4: 矮曲灌木型 |
为了分析4种类型聚集程度之间的差异,首先提取g(r)统计量进行配对t检验,由于g(r)统计量为连续数值,选取其中511个值进行检验,结果(表 2)表明几种类型之间聚集度有显著差别。
| 类型Type | t统计量 | mean difference | P | df |
| s1—s2 | -21.9085 | -3.993731 | < 0.001 | 511 |
| s1—s3 | 12.4781 | 1.974787 | < 0.001 | 511 |
| s1—s4 | 13.4009 | 1.678653 | < 0.001 | 511 |
| s2—s3 | 35.5429 | 5.968518 | < 0.001 | 511 |
| s2—s4 | 35.3676 | 5.672384 | < 0.001 | 511 |
| s3—s4 | -3.3264 | -0.2961342 | < 0.001 | 511 |
利用gcross()双变量检验进行空间关联分析结果表明(图 6),在0—10m的较小尺度上,s1与s2、s1与s4、s2与s3、s2与s4均表现为正关联;s1与s3、s3与s4表现为无明显促进或排斥;在10—20m的较大尺度上,各阶段之间皆表现为无明显促进或排斥关系。

|
| 图 6 各类型之间的空间关联 Fig. 6 Spatial correlation of multiple types x轴代表空间尺度,y轴为双变量g(r)值,虚线中间的区域为完全随机分布的置信区间 |
本研究中具有一个明显主干的乔木型岳桦和灌木型岳桦分别占总体的7.46%和31.95%,对总体树高特征的研究表明该研究区内岳桦树高生长受限,且存在一个关键的树高区间1.55—2.5m。王晓东[9]等对同一区域岳桦的研究结果表明,岳桦在径级18cm 时达到树高生长的极限,说明林线处树高生长明显受环境限制,这与本研究的结果一致。目前关于树高生长受限机理的研究多集中于成熟假说、营养限制假说、呼吸假说以及水力限制假说[10, 11, 12, 13],其中水力限制假说受到更多关注,Koch[14]等认为重力以及运输路径限制了叶片大小和光合能力,最终引起树高生长受限。功能生态学观点认为乔木到灌木的优势生活型变化有利于矮曲植物体和分生组织在白天获得高于气温的温度和维持植物生长所需的正热量平衡[8],低矮的植物体和密集的地被层可使植被脱离冠层以上恶劣气候的作用,隔绝周围气候因素的影响[15]。关于高山植物与微气候的研究认为,低矮的灌木、草本或苔原植物与乔木相比,更容易隔绝周围气候因素的影响,也更能够在叶冠中积累热量[16]。此外,如果认为优势生活型的选择是植物主动适应、权衡(Trade-off)的结果,在高生长不能为植物带来光获取的优势,反而增加其他胁迫风险如风雪侵袭的情况下,植物适应性地选择灌木型以抵御胁迫条件[17],有可能是高海拔地区灌木取代乔木生活型的原因。
3.1.2 岳桦种群分布格局本研究中岳桦种群表现为0—10m较小尺度上的聚集分布和10—20m较大尺度上的均匀分布,这与王晓春等利用地统计学方法分析材积数据得到的分布趋势相同[7]。某一尺度上的空间格局是一个或几个生态过程共同作用的结果,较大尺度上的格局更有可能是物种分布异质性、斑块性以及不同的环境条件决定,而较小尺度上的格局可能是不同程度的种内、种间竞争、种子扩散限制等因素所致。长白山北坡林线岳桦分布受地形因子的影响尤其明显,地形可能与其他形式的低温胁迫结合在一起,影响岳桦种群分布格局。对样地的融雪格局持续监测结果显示,5月末的融雪期,大岳桦集中于雪斑边缘,而在没有雪斑覆盖的裸露地段,少有岳桦分布,融雪格局与岳桦分布格局相关性较好。相比于裸地,雪被下的土壤温度降低了岳桦用于抵抗低温胁迫的碳投资。此外,有研究认为岳桦种群径级结构和土壤养分在林线内空间分异明显,岳桦通过不断调整生活型降低土壤限制积极应对气候变化[18]。
3.1.3 不同生活史阶段的空间格局与关联不同生活史阶段岳桦的分布格局可以在一定程度上反应种群的发育过程,在不区分海拔的情况下,长白山北坡岳桦在其发育过程中由集群分布发展为随机分布[6]。本研究中林线岳桦表现出不同的趋势,0—5m的小尺度下各个阶段均表现为聚集分布,5—20m范围内中树和老树表现为相对聚集,表明其对空间的异质性选择效果最强,相反幼苗表现为随机分布,其对空间异质性选择的效果相对较弱。此外相对于乔木型,灌木型岳桦表现为相对随机的分布,这种聚集程度和空间选择的差异可能指示了矮曲、直立岳桦之间对环境适应性选择的差异,矮曲、灌木代表的“紧凑生活型”可能具有一定的适应性优势,而乔木更多地体现在占据有利位置。岳桦发育过程中可能存在一个特定的时期,个体适应性地选择一种合适的生存策略,如果有大树或群丛的保护,以直立主干的形式迅速生长,如果在没有隐蔽保护的条件下,以灌木生活型减少与周围环境因子的耦合,保证生存。年轮年代学方法得出乌拉尔山4个区域的优势树种具有从多主干到独立主干的趋势,且认为这种趋势与冬季降雪增多导致的相对温暖环境有关[19, 20],表明可能存在这一生活型转化的特定阶段。
树木个体之间的相互影响具有一定的作用范围,考虑物种的空间分布关联时更为关注较小尺度。本研究中0—10m的较小尺度上,除s1与s3、s3与s4表现为无明显促进或排斥外,其他类型间均表现出促进作用。值得关注的是幼苗与老树、灌木型之间均无明显促进作用,仅与中树存在显著正关联。而仅有中树与其他类型都表现为正相关,可能的原因是中树所占比例过大,本文中划分方法没有合理区分开生活史的各阶段,或者岳桦树高生长受环境因素影响较大。
3.2 结论(1)长白山北坡林线岳桦树高生长受到限制,1.5—3.0m是一个关键的树高指标。
(2)相比于老树、中树,幼苗和灌木型岳桦更为均匀,对空间的异质性选择更弱。
(3)林线岳桦发育过程中,存在一个生活型分离的重要阶段。
| [1] | Körner C. A re-assessment of high elevation treeline positions and their explanation. Oecologia, 1998, 115(4): 445-459. |
| [2] | Körner C. Alpine Plant Life: Functional Plant Ecology of High Mountain Ecosystems. Berlin: Springer, 2003: 3-40. |
| [3] | Tirado R, Pugnaire F I. Community structure and positive interactions in constraining environments. Oikos, 2005, 111(3): 437-444. |
| [4] | Gittins R. Canonical Analysis: A Review with Applications in Ecology. Berlin: Springer, 1985. |
| [5] | 周国英, 陈桂琛, 魏国良, 韩友吉, 祝存冠. 青海湖地区芨芨草群落主要种群分布格局研究. 西北植物学报, 2006, 26(3): 579-584. |
| [6] | 于大炮, 唐立娜, 王绍先, 王顺忠, 代力民. 长白山北坡植被垂直带群落交错区的定量判定. 应用生态学报, 2004, 15(10): 1760-1764. |
| [7] | 王晓春, 韩士杰, 邹春静, 周晓峰. 长白山岳桦种群格局的地统计学分析. 应用生态学报, 2002, 13(7): 781-784. |
| [8] | Körner C. High elevation treelines//Körner C, ed. Alpine Treelines. Berlin: Springer, 2012. |
| [9] | 王晓东, 刘慧清. 长白山突变型林线岳桦对气候变化的响应. 山地学报, 2011, 29(5): 551-560. |
| [10] | Drake J E, Raetz L M, Davis S C, DeLucia E H. Hydraulic limitation not declining nitrogen availability causes the age-related photosynthetic decline in loblolly pine (Pinus taeda L.). Plant, Cell & Environment, 2010, 33(10): 1756-1766. |
| [11] | Ryan M G, Yoder B J. Hydraulic limits to tree height and tree growth. BioScience, 1997, 47(4): 235-242. |
| [12] | Thomas S C. Photosynthetic capacity peaks at intermediate size in temperate deciduous trees. Tree Physiology, 2010, 30(5): 555-573. |
| [13] | Thomas S C, Winner W E. Photosynthetic differences between saplings and adult trees: an integration of field results by meta-analysis. Tree Physiology, 2002, 22(2/3): 117-127. |
| [14] | Koch G W, Sillett S C, Jennings G M, Davis S D. The limits to tree height. Nature, 2004, 428(6985): 851-854. |
| [15] | Grace J, Berninger F, Nagy L. Impacts of climate change on the treeline. Annals of Botany, 2002, 90(4): 537 -544. |
| [16] | Scherrer D, Körner C. Infra-red thermometry of alpine landscapes challenges climatic warming projections . Global Change Biology, 2010, 16(9): 2602-2613. |
| [17] | Körner C, Paulsen J, Pelaez-Riedl S. A bioclimatic characterization of Europe's alpine areas//Nagy L, Grabherr G, Körner C, Thompson D B A, eds. Alpine Biodiversity in Europe. Berlin: Springer, 2003: 13-28. |
| [18] | 王晓东, 刘惠清. 长白山北坡岳桦林线变动的水热条件分析. 地理科学进展, 2011, 30(3): 313-318. |
| [19] | Hagedorn F, Shiyatov S G, Mazepa V S, Devi N M, Grigor'ev A A, Bartysh A A, Fomin V V, Kapralov D S, Terent'ev M, Bugman H, Rigling A, Moiseev P A. Treeline advances along the Urals mountain range-driven by improved winter conditions?. Global Change Biology, 2014, 20(11): 3530-3543. |
| [20] | Aune S, Hofgaard A, Söderström L. Contrasting climate- and land-use-driven tree encroachment patterns of subarctic tundra in northern Norway and the Kola Peninsula. Canadian Journal of Forest Research, 2011, 41(3): 437-449. |