 2015, Vol. 35
2015, Vol. 35文章信息
- 金慧, 赵莹, 赵伟, 尹航, 秦立武, 崔凯峰, 李晓杰, 黄祥童
- JIN Hui, ZHAO Ying, ZHAO Wei, YIN Hang, QIN Liwu, CUI Kaifeng, LI Xiaojie, HUANG Xiangtong
- 长白山牛皮杜鹃群落物种多样性的海拔梯度变化及相似性
- Elevational changes in species diversity and similarity analysis of a Rhododendron chrysanthum community on Changbai Mountain
- 生态学报, 2015, 35(1): 125-133
- Acta Ecologica Sinica, 2015, 35(1): 125-133
- http://dx.doi.org/10.5846/stxb201405120971
-
文章历史
- 收稿日期:2014-05-12
- 修订日期:2014-11-14






牛皮杜鹃(Rhododendron chrysanthum Pall.)为杜鹃花科多年生常绿小灌木,株高10—25cm,径横卧。主要生长于长白山海拔1300—2650m[1, 2, 3],属长白山高山苔原带生态系统中的优势种群[4]。目前,对牛皮杜鹃大多集中在遗传结构、组织培养和植株解剖等方面的研究[4, 5, 6, 7]。关于牛皮杜鹃群落结构和生物多样性方面鲜有报道,尤其是牛皮杜鹃群落在不同海拔梯度的物种组成更是未见报道。物种多样性是表征群落学的重要指标,在反映植物群落的生境差异、群落结构组成和稳定性程度等方面具有重要的意义[8, 9],高山植物群落物种多样性随海拔梯度的变化始终是生态学家的研究热点[10]。本文以长白山牛皮杜鹃群落为研究对象,综合调查牛皮杜鹃群落物种组成及生物多样性沿海拔梯度的变化,揭示牛皮杜鹃群落生物多样性的变化规律,旨在深入了解牛皮杜鹃群落物种多样性的海拔分布格局,为牛皮杜鹃群落生物多样性的保护和管理提供理论依据。
1 研究区域概况研究区位于长白山北坡海拔1900—2150m的岳桦-苔原过渡带和海拔2150m以上高山苔原带。气候条件属于湿润性亚高山气候,冬季寒冷多风,夏季温凉短暂。年平均气温低,生长季节短。岳桦-苔原过渡带土壤为火山熔岩或火山浮石上发育起来的山地生草森林土,土质贫瘠,土层较薄[11]。高山苔原带土壤受到成土母岩和风化壳类型的制约,表层出现泥炭化和粗腐殖质的积累,土壤表现为粗骨性和薄层性[12]。特殊的环境条件发育着特殊的植被类型,在岳桦-苔原过渡带,岳桦 (Betula ermanii)与长白落叶松(Larix olgensis)、东北赤杨(Alnus mandshurica)等混生,林下灌木种类较多,主要有牛皮杜鹃(Rhododendron chrysanthum)、西伯利亚刺柏(Juniperus sibirica)、笃斯越桔 (Vaccinium uliginosum)等[10];高山苔原带植被矮小,灌木匍匐生长,呈座垫状,耐严寒,抗风力强,根系浅而发达,地下部分明显多于地上部分[12]。调查的牛皮杜鹃群落植物组成随海拔变化有所不同,主要植物种类为牛皮杜鹃(Rhododendron chrysanthum)、松毛翠(Phyllodoce caerulea ) 、笃斯越桔(Vaccinium uliginosum) 、苞叶杜鹃(Rhododendron bracteatum)、宽叶仙女木(Dryas octopetala var.asiatica)、圆叶柳(Salix crenata) 、毛山菊(Dendran themum zawadskii var.alpinum)、高山罂粟(Papaver alpinum) 等。
2 研究方法 2.1 样地设置与调查以长白山北坡海拔1900—2600m牛皮杜鹃群落为研究区域,根据研究区域的立地条件,采用样地调查法,自海拔1900m至2600m利用GPS定位,分别在10个不同海拔地段具有代表性的牛皮杜鹃群落设置样地,样地面积为20m×20m,在样地内选取8个面积5m×5m有代表性的小样方用于灌木层物种多样性的调查,再取8个面积为1m×1m的小样方用于草本层物种多样性的调查。调查内容包括样地经纬度、海拔高度、坡度等基本概况以及灌木、草本植物的种数、株高、地径、冠幅、株数等。样地概况见表 1所示。
|
海拔
Elevation/m | 经度
Longitude | 纬度
Latitude | 坡度
Slope/(°) | 坡向
Aspect/(°) | 群落高
Community height/m | 群落外貌
Community physiognomy |
| 1926 | 128°04′15.38″ | 42°03′37.41″ | 25 | 130 | 13 | 林型主要为岳桦-牛皮杜鹃林,灌木以牛皮杜鹃为主,伴生蓝靛果忍冬、花楸;牛皮杜鹃为散生片状分布 |
| 1949 | 128°04′19.55″ | 42°03′38.64″ | ≥5 | 140 | 12 | 片段化群落外围,乔木以岳桦为主,牛皮杜鹃为优势灌木,草本层不发达 |
| 1986 | 128°03′58.0″ | 42°03′27.50″ | 15 | 175 | 13 | 乔木为岳桦,牛皮杜鹃均匀分布,群落稳定 |
| 1992 | 128°03′57.2″ | 42°03′25.10″ | 7—10 | 158 | 8 | 岳桦上限,岳桦稀疏,牛皮杜鹃矮小,均块状分布 |
| 2010 | 128°03′57.9″ | 42°03′14.50″ | 15 | 65 | 5 | 岳桦上限,牛皮杜鹃片状分布,有小块状生长不良 |
| 2250 | 128°04′25.37″ | 42°02′28.32″ | 35 | 40 | 0.30 | 沟谷上沿,牛皮杜鹃多分布在中断沟谷底部 |
| 2350 | 128°04′21.97″ | 42°02′27.97″ | 10 | 100 | 0.31 | 沟谷边缘分布牛皮杜鹃,而沟谷外牛皮杜鹃少量分布,以笃斯越桔为主 |
| 2450 | 128°04′19.44″ | 42°02′11.82″ | 30 | 35 | 0.28 | 牛皮杜鹃斑块状分布,灌木总盖度85% |
| 2528 | 128°04′18.12″ | 42°02′05.70″ | 35 | 175 | 0.13 | 牛皮杜鹃盖度达80%,矮小,偶见毛毡杜鹃、苞叶杜鹃 |
| 2598 | 128°04′20.98″ | 42°01′54.97″ | 10 | 95 | 0.15 | 有河流冲刷小沟谷,斑块状群落外围镶嵌式分布 |
研究中调查数据处理与统计分析,均采用SPSS17.0软件和Microsoft Office Excel 2003软件完成。
2.2.1 重要值与综合优势比[13]重要值 VIt=(相对盖度+相对频度+相对密度)/3
综合优势比 SDR4t=(相对多度+相对盖度+相对频度+相对高度)/4
2.2.2 群落α多样性[13](1)物种丰富度指数(D1,D2)
Gleason指数
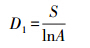
Marglef指数
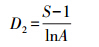
(2)Shannon-Wiener物种多样性指数(SW)
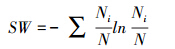
(3)修正后的Simpson生态优势度指数(SN)
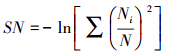
(4)种间机遇机率Hurlbert指数(H)

(5)Pielou群落均匀度(R)
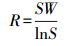
(1)Whittaker指数(βw)
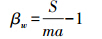
(2)Cody指数(βC)

(3)Routledge指数(βR)
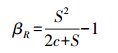
采用Jaccard相似性公式计算相似性系数[13]:

调查的牛皮杜鹃群落中(表 2),共包含灌木种类14种,草本种类96种。从不同海拔的物种总数上看,低海拔高度时牛皮杜鹃群落的物种数最多,达到34种。随着海拔的升高,物种总数呈先减少再增加再减少的趋势,主要与长白山岳桦上线和高山苔原带特殊的地形条件有关。海拔2000m为长白山岳桦上线,岳桦带植物逐渐消失而仅出现部分适合生存的苔原带植物,物种总数相对减少,而海拔2300m左右为苔原带植物生长的最佳生境,物种总数最多。
|
植被层次 Vegetation Layer | 物种数目number of species | |||||||||
| 1926 | 1949 | 1986 | 1992 | 2010 | 2250 | 2350 | 2450 | 2528 | 2598 | |
| 灌木层Shrub layer | 6 | 2 | 7 | 4 | 3 | 7 | 7 | 7 | 7 | 6 |
| 草本层Herb layer | 28 | 23 | 7 | 8 | 11 | 23 | 24 | 20 | 13 | 16 |
| 总计Total | 34 | 25 | 14 | 12 | 14 | 30 | 31 | 27 | 20 | 22 |
从灌木层的重要值特征与综合优势比(表 3)可知,牛皮杜鹃在整个群落中的重要值和优势度最高,低于2000m的牛皮杜鹃群落,灌木物种越桔在各海拔梯度均有分布,西伯利亚刺柏除1949m没有出现,其它样方均有分布。这一结果显示,长白山岳桦与苔原过渡带调查的牛皮杜鹃群落中,牛皮杜鹃种群为优势种群,越桔为次优种群。
|
灌木树种
Shrub species | 重要值与综合优势比VIi and SDR4i/% | |||||||||
| 1926 | 1949 | 1986 | 1992 | 2010 | 2250 | 2350 | 2450 | 2528 | 2598 | |
|
牛皮杜鹃
Rhododendron hrysanthum | 72.2
(57.7) | 87.3
(87.2) | 60.1
(49.3) | 64.3
(55.9) | 100.0
(100.0) | 41.9
(39.0) | 26.7
(24.7) | 37.9
(36.2) | 39.6
(37.1) | 17.3
(22.3) |
|
越桔
Vaccinium vitisidaea | 2.2
(2.7) | 12.7
(12.8) | 17.4
(14.0) | 21.9
(18.4) | — | — | — | — | — | — |
|
笃斯越桔
Vaccinium uliginosum | — | — | 1.7
(2.6) | 11.8
(13.7) | — | 10.7
(11.2) | 22.3
(24.9) | 6..4
(8.2) | 3.5
(5.5) | 6.9
(9.7) |
|
西伯利亚刺柏
Juniperus sibirica | 8.9
(10.0) | — | 5.2
(8.7) | 2.1
(12.0) | — | — | — | — | — | — |
|
花楸
Sorbus pohuashanensis | 5.4
(1.6) | — | — | — | — | — | — | — | — | — |
|
蓝靛果忍冬
Lonicea caerulea | 4.7
(7.5) | — | 1.7
(7.3) | — | — | — | — | — | — | — |
|
林奈木
Linnaea borealis | 2.3
(2.5) | — | 12.5
(12.1) | — | — | — | — | — | — | — |
|
毛毡杜鹃
Rhododendron confertissimum | — | — | — | — | — | 3.1
(4.7) | 16.0
(14.4) | 1.0
(3.3) | 4.1
(6.3) | 3.7
(5.9) |
|
松毛翠
Phyllodoce caerulea | — | — | — | — | — | 19.5
(18.7) | 5.3
(7.1) | 21.2
(18.9) | 19.6
(17.7) | — |
|
兴安杜鹃
Rhododendron dauricum | — | — | 1.4
(6.0) | — | — | — | — | — | — | — |
|
苞叶杜鹃
Rhododendron redowskianum | — | — | — | — | — | 10.4
(10.1) | 10.2
(10.2) | 15.0
(14.0) | 15.4
(15.3) | 6.6
(7.8) |
|
宽叶仙女木
Dryas octopetala var. asiatica | — | — | — | — | — | 5.2
(5.9) | 18.6
(16.4) | 5.7
(6.2) | 7.1
(7.5) | 43.8
(35.2) |
|
圆叶柳
Salix crenata | — | — | — | — | — | 9.3
(10.4) | 0.9
(2.3) | 12.7
(13.3) | 10.8
(10.5) | 21.7
(19.2) |
海拔2000m以上的数据显示,随着海拔的升高,牛皮杜鹃群落中灌木物种组成发生明显变化,越桔、西伯利亚刺柏等物种全部消失,随之出现苞叶杜娟、宽叶仙女木、圆叶柳等灌木。达到高海拔时,物种组成基本稳定。牛皮杜鹃种群作为群落的优势种群,除海拔2598m无松毛翠,其它海拔梯度上松毛翠的重要值和优势度均较高,作为群落中的次优势种存在。各群落类型的分布主要受海拔和坡度、坡位、生境条件等小地形因素控制。
3.2 牛皮杜鹃群落沿海拔梯度的生物多样性特征 3.2.1 群落α多样性的垂直变化植物群落作为物种种群对环境反应的集合体,其群落本身特征随物种组成变化而变化。α多样性是表征植被群落内物种组成特征的一项重要指标(表 4)。由于长白山特殊地理环境,水热条件严酷,致使灌木物种分布有限,草本物种类型较为丰富。研究发现,牛皮杜鹃群落相同海拔高度,草本层的物种多样性普遍高于灌木层的物种多样性。
灌木层α多样性结果显示,除丰富度指数D2变化不明显,其他各指数沿海拔梯度的变化基本表现出一致性,自海拔1926—1986m,多样性指数先降低后升高,1986m后再次降低,到达海拔2010m降到最低点后多样性指数开始回升,直至海拔2250m,生物多样性指数的变化趋于平缓。说明沿海拔的升高,到达高山苔原带后,适应高山苔原带特殊生境条件的物种增多,海拔2250—2528m范围内,达到最为适宜的气候条件,群落的各多样性指数除生态优势度指数SN外,其他指数的变化趋于平稳状态,物种组成相对较为稳定。海拔2528m以上由于高海拔区域气候寒冷,土层渐薄,火山灰层加厚,限制了植物的生存,生物多样性再次呈降低趋势。研究结果与长白山特殊的地理环境有关,1949—2010m为岳桦上线,2010m以上为高山苔原带。
|
海拔/m
Elevation | 层次
Layer | Simpson
生态优势度指数 (SN) | Shnannon
物种多样性指数 (SW) | Gleason
丰富度指数 (D1) | Marglef
丰富度指数 (D2) | Pielou
均匀度指数 (R) | Hurlbert
种间机遇指数 (H) |
| 1926 | 灌木shurb | 0.625 | 0.925 | 1.731 | 0.786 | 0.516 | 0.466 |
| 草本herb | 2.911 | 3.069 | 13.465 | 4.251 | 0.921 | 0.947 | |
| 1949 | 灌木shurb | 0.251 | 0.381 | 0.577 | 0.194 | 0.550 | 0.223 |
| 草本herb | 2.576 | 2.821 | 11.061 | 3.130 | 0.900 | 0.925 | |
| 1986 | 灌木shurb | 0.890 | 1.222 | 2.020 | 0.823 | 0.628 | 0.590 |
| 草本herb | 2.618 | 1.694 | 3.366 | 0.984 | 0.870 | 0.786 | |
| 1992 | 灌木shurb | 0.744 | 0.950 | 1.154 | 0.472 | 0.685 | 0.526 |
| 草本herb | 2.911 | 1.622 | 3.847 | 1.094 | 0.780 | 0.947 | |
| 2010 | 灌木shurb | 0.000 | 0.000 | 0.289 | 0.000 | 0.000 | |
| 草本herb | 2.618 | 2.296 | 5.290 | 1.761 | 0.957 | 0.894 | |
| 2250 | 灌木shurb | 1.395 | 1.639 | 2.020 | 0.862 | 0.842 | 0.753 |
| 草本herb | 2.819 | 2.968 | 11.061 | 3.639 | 0.947 | 0.943 | |
| 2350 | 灌木shurb | 0.638 | 1.724 | 2.020 | 0.855 | 0.886 | 0.806 |
| 草本herb | 2.894 | 3.034 | 11.542 | 3.474 | 0.955 | 0.946 | |
| 2450 | 灌木shurb | 1.447 | 1.629 | 2.020 | 0.840 | 0.837 | 0.765 |
| 草本herb | 2.529 | 2.755 | 9.618 | 3.019 | 0.920 | 0.922 | |
| 2528 | 灌木shurb | 1.435 | 1.650 | 2.020 | 0.819 | 0.848 | 0.762 |
| 草本herb | 2.215 | 2.367 | 6.252 | 1.966 | 0.923 | 0.893 | |
| 2598 | 灌木shurb | 1.274 | 1.482 | 1.731 | 0.831 | 0.827 | 0.722 |
| 草本herb | 2.493 | 2.607 | 7.694 | 2.128 | 0.940 | 0.918 |
牛皮杜鹃群落草本层的α多样性指数显示,生态优势度指数SN、种间机遇指数H的变化趋势不明显,而物种多样性指数SW、丰富度指数D和均匀度指数R沿海拔梯度的变化大致表现相同的变化趋势。自海拔1926m开始,先降低后升高,到达2350m时为最大值,随后继续下降,到达2528m后再次升高,但是此阶段变化不显著。
3.2.2 群落α多样性指数间的相关及回归分析在群落多样性梯度上,在不同的空间水平,群落各种α多样性指数对于群落物种多样性的贡献不同。
根据相关分析结果可知(表 5):牛皮杜鹃群落各α多样性指数间呈P<0.01水平极显著正相关性,由于各变量之间存在着高度相关性,对物种多样性指数(SW)与其他各指数之间进行了回归分析:
| SW | SN | D1 | D2 | R | H | |
| **代表P<0.01水平的差异显著性 | ||||||
| SW | 1 | |||||
| SN | 0.877* * | 1 | ||||
| D1 | 0.910* * | 0.811* * | 1 | |||
| D2 | 0.920* * | 0.795* * | 0.992* * | 1 | ||
| R | 0.863* * | 0.763* * | 0.651* * | 0.650* * | 1 | |
| H | 0.903* * | 0.864* * | 0.681* * | 0.699* * | 0.904* * | 1 |
SW与生态优势度指数SN的回归方程
Y=-0.049+0.985X
SW与丰富度指数D1的回归方程
Y=-3.107+4.368X
SW与丰富度指数D2的回归方程
Y=-0.809+1.306X
SW与均匀度指数R的回归方程
Y=0.541+0.148X
SW与种间机遇指数H的回归方程
Y=0.250+0.264X
从对群落物种多样性SW的贡献上看,在整个牛皮杜鹃群落中,物种丰富度对群落的多样性指数贡献率最大,表现为丰富度指数>种间机遇指数>生态优势度指数>群落均匀度指数。
3.2.3 群落β多样性的垂直变化牛皮杜鹃群落β多样性沿海拔梯度基本呈波形变化,牛皮杜鹃群落β多样性极值基本出现在群落的交错地带。表 6显示,牛皮杜鹃群落的草本层β多样性指数普遍高于灌木层β多样性指数,这意味着在牛皮杜鹃群落物种沿海拔梯度的替换速率上,草本植物高于灌木物种。Routledge指数的变化趋势不显著,而Cody指数的变化灌木层和草本层基本相同。随着海拔的逐渐升高,Cody指数在海拔1986m和海拔2250m处,出现两次极值。第一处极值反映了高山岳桦带向高山苔原带的过渡;第二处极值反映了草本层物种的构成差异较大。海拔2250m以上灌木层的Whittaker 多样性指数和Cody指数逐渐趋于平稳,说明海拔上段群落灌木层之间的差异和变化较小。β指数波动与相邻样方间的水平距离引起的物种构成变化大小有关[15]。
|
海拔区间/m
Elevation range | 层次
Layer | Whittaker指数
(βw) | Cody指数
(βC) | Routledge指数
(βR) |
| 1926—1949 | 灌木 | 6.000 | 2.000 | 11.739 |
| 草本 | 3.174 | 12.500 | 90.275 | |
| 1949—1986 | 灌木 | 1.000 | 2.500 | 11.739 |
| 草本 | 12.714 | 16.500 | 93.633 | |
| 1986—1992 | 灌木 | 2.500 | 1.500 | 10.686 |
| 草本 | 11.000 | 6.000 | 95.000 | |
| 1992—2010 | 灌木 | 3.667 | 1.500 | 13.000 |
| 草本 | 7.727 | 7.500 | 92.305 | |
| 2010—2250 | 灌木 | 1.000 | 4.000 | 13.000 |
| 草本 | 3.174 | 14.000 | 92.852 | |
| 2250—2350 | 灌木 | 1.000 | 0.000 | 9.955 |
| 草本 | 3.000 | 10.500 | 90.275 | |
| 2350—2450 | 灌木 | 1.000 | 0.000 | 9.955 |
| 草本 | 3.800 | 11.000 | 90.432 | |
| 2450—2528 | 灌木 | 1.000 | 0.000 | 9.955 |
| 草本 | 6.385 | 7.500 | 90.798 | |
| 2528—2598 | 灌木 | 1.333 | 0.500 | 10.147 |
| 草本 | 5.000 | 3.000 | 90.432 |
采用Jaccard相似性系数作为不同海拔间群落相似性的度量标准,并加以分析。如表 7所示,海拔2250m与海拔2450m群落相似性系数为0.583;海拔2350m与海拔2528m群落相似性系数为0.545;海拔2450m与海拔2528m群落相似性系数为0.516,海拔2528m与海拔2598m群落相似性系数为0.680,以上各海拔牛皮杜鹃群落均呈现中等相似水平;海拔1926m与海拔1949m群落相似性系数为0.311;海拔1926m与海拔1992m相似性系数为0.278,海拔2250 m与海拔2350m、2528m、2598m之间,海拔2350m与海拔2450m、2598m之间,海拔2450m与海拔2592m之间群落相似性系数均在0.25—0.50之间,呈中等不相似水平;其他各群落之间的相似性系数均在0.00—0.25间,属于极不相似水平。海拔梯度间生境及群落结构差异性越大,物种多样性变化越明显,海拔高度接近的群落间相似性系数较高。
|
海拔/m Elevation | 1926 | 1949 | 1986 | 1992 | 2010 | 2250 | 2350 | 2450 | 2528 |
| 括号内数字表示两样地重叠的植物种类数 | |||||||||
| 1949 | 0.311(10) | ||||||||
| 1986 | 0.127(8) | 0.147(5) | |||||||
| 1992 | 0.278(10) | 0.156(5) | 0.238(1) | ||||||
| 2010 | 0.200(8) | 0.147(5) | 0.217(5) | 0.300(6) | |||||
| 2250 | 0.085(5) | 0.078(4) | 0.048(2) | 0.105(4) | 0.100(3) | ||||
| 2350 | 0.048(3) | 0.037(2) | 0.098(4) | 0.132(5) | 0.071(3) | 0.452(12) | |||
| 2450 | 0.070(4) | 0.061(3) | 0.051(2) | 0.083(3) | 0.051(2) | 0.583(21) | 0.450(11) | ||
| 2528 | 0.038(2) | 0.023(1) | 0.097(3) | 0.143(4) | 0.063(2) | 0.471(16) | 0.545(18) | 0.516(9) | |
| 2598 | 0.037(2) | 0.044(2) | 0.091(3) | 0.133(4) | 0.059(2) | 0.405(15) | 0.472(17) | 0.400(14) | 0.680(11) |
(1)牛皮杜鹃群落沿海拔梯度的变化逐渐形成以灌木为主、多年生草本植物为辅的群落结构类型,牛皮杜鹃作为优势种群,次优种群随着海拔升高而发生改变。说明牛皮杜鹃群落物种组成受海拔和坡度、坡位、生境条件等多因素控制。
(2)群落α多样性具有海拔梯度、纬度梯度等多种环境梯度格局。海拔梯度作为环境梯度格局中的主导梯度[16],引起相关学者的极度重视。天山南麗山前平原植物群落物种多样性随海拔的增加而增加[17];关帝山海拔2500—2755m 的灌丛草甸群落物种多样性随着海拔的升高综合多样性呈下降趋势[18]; 青海高寒草甸地区海拔3840—4435m物种多样性呈先增加后减少的趋势[19]。关于牛皮杜鹃群落生物多样性研究中,在1926—2010m海拔区间内,α多样性指数的极值出现在了海拔1986m,灌木层α多样性指数在该处出现最大值,而草本层α多样性指数在该处出现最小值。主要由于海拔1986m作为长白山岳桦上限,处于与高山苔原带的交界处,物种组成发生着明显的变化,适合该处生境生存的草本植物较少,而高山灌木相对的适应性更强。在2010—2598m海拔区间内,在2250m出现极值后趋于平缓,该海拔地段适生物种已经趋于稳定,人为干扰较小。
牛皮杜鹃群落间各α多样性指数间呈P<0.01水平极显著正相关性。在整个牛皮杜鹃群落中,物种丰富度对群落多样性的贡献率最大。这与以往的大多研究相一致,柳妍妍等在天山南坡中段高寒草地的研究结果表明,群落的物种丰富度指数对多样性的贡献率大于其他指数[20]。杨利民等在松嫩平原草地的研究结论显示群落物种多样性受丰富度指数的影响大于受均匀度指数的影响[21]。
(3)群落中物种β多样性反映了一定距离水平上群落物种组成的差异以及环境梯度上群落物种组成的更替。研究发现物种β多样性除了灌木层的Whittaker 多样性指数略有不同,其他指数沿海拔梯度同样在1986m和2250m出现了两个极值,但大体呈波形变化,由于长白山岳桦林带向高山苔原带过渡带物种变化速率较快,β多样性出现一个极大值。2250m适生物种的种类再次发生变化,物种变化速率较快,β多样性第二次出现极大值,而后趋于平缓。而灌木层的Whittaker 多样性指数和Cody指数均反映了以2250m为分界,两侧的物种构成的相似性和垂直替代。
(4)不同海拔梯度间生境及群落结构差异性较大,物种多样性变化相对较为明显。海拔高度接近的群落间相似性系数较高,说明海拔是影响牛皮杜鹃群落差异的主要因素,种群扩散与种群定居均受海拔高度的影响,植物群落的物种组成随着海拔的变化而发生变化。
(5)从牛皮杜鹃物种多样性的海拔格局来看,海拔相近的区域,由于不同的地形条件致使物种多样性间的差异,研究未对海拔以外的影响因子加以分析,但是较小海拔范围内其它因素的影响不容忽视。因此,以后的研究中应针对牛皮杜鹃群落物种多样性在大小尺度上的差异进行研究方法的改进,对物种多样性沿海拔梯度的同一尺度变化以及地形等其他因子对物种多样性的影响有待于进一步研究。
| [1] | 刘景双. 长白山高山苔原牛皮杜鹃群落物质循环的研究. 生态学报, 1993, 13(1): 96-98. |
| [2] | 钱宏. 长白山高山冻原植被. 森林生态系统, 1992, 6: 72-95. |
| [3] | 周繇, 朱骏义, 于俊林. 中国长白山观赏植物彩色图志. 吉林: 吉林教育出版社, 2005: 378-378. |
| [4] | 宫宇, 刘宪虎, 张春影, 李美善, 项美玲. 不同地区牛皮杜鹃叶片解剖结构比较分析. 延边大学农学学报, 2010, 32(1): 22-25. |
| [5] | 杨志勇. 牛皮杜鹃遗传多样性的研究[D]. 长春: 吉林大学, 2009. |
| [6] | 徐颖, 金灿, 宗成文, 于秋艳, 曹后男. 长白山牛皮杜鹃组培快繁及初步驯化栽培研究. 江苏农业科学, 2010, (2): 47-49. |
| [7] | 丁洪玲, 宫宇, 李美善, 许明子, 刘宪虎. 牛皮杜鹃植株解剖结构的研究. 延边大学农学学报, 2008, 30(2): 90-97. |
| [8] | 苗莉云, 王孝安, 王志高. 太白红杉群落交错带物种多样性的研究. 广西植物, 2005, 25(2): 112-116. |
| [9] | Lomolino M V. Elevation gradients of species-density: historical and prospective views. Global Ecology& Biogeography, 2001, 10(1): 3-13. |
| [10] | 朱登强, 王军辉, 张守攻, 张建国, 茹广欣, 罗大庆, 潘刚. 西藏色季拉山西坡急尖长苞冷杉林物种多样性及群落结构的垂直分布格局. 西北林学院学报, 2008, 23(5): 1-6. |
| [11] | 邹春静, 韩士杰, 周玉梅, 王晓春, 陈永亮. 过渡带中岳桦种群生态特征的研究. 应用与环境生物学报, 2001, 7(1): 1-6. |
| [12] | 孟宪玺. 长白山的高山苔原土. 地理科学, 1982, 2(1): 57-64. |
| [13] | 张金屯. 数量生态学. 北京: 科学出版社, 2004. |
| [14] | 沈蕊, 张建利, 何彪, 李峰, 张志明, 周睿, 欧晓昆. 元江流域干热河谷草地植物群落结构特征与相似性分析. 生态环境, 2010, 19(12): 2821-2825. |
| [15] | 沈泽昊, 刘增力, 方精云. 贡嘎山海螺沟冷杉群落物种多样性与群落结构随海拔的变化. 生物多样性, 2004, 12(2): 237-244. |
| [16] | 何艳华, 闫明, 张钦弟, 张直峰, 苗艳明, 毕润成. 五鹿山国家级自然保护区物种多样性海拔格局. 生态学报, 2013, 33(8): 2452-2462. |
| [17] | 赵振勇, 王让会, 尹传华, 张慧芝, 王雷. 天山南麓山前平原植物群落物种多样性及空间分异研究. 西北植物学报, 2007, 27(4): 784-790. |
| [18] | 袁建英, 张金屯, 席跃翔. 山西关帝山亚高山灌丛、草甸物种多样性的研究. 草业学报, 2004, 13(3): 34-39. |
| [19] | 王长庭, 王启基, 龙瑞军, 景增春, 史惠兰. 高寒草甸群落植物多样性和初级生产力沿海拔梯度变化的研究. 植物生态学报, 2004, 28(2): 240-245. |
| [20] | 柳妍妍, 胡玉昆, 王鑫, 公延明. 天山南坡中段高寒草地物种多样性与生物量的垂直分异特征. 生态学杂志, 2013, 32(2): 311-318. |
| [21] | 杨利民, 周广胜, 李建东. 松嫩平原草地群落物种多样性与生产力关系的研究. 植物生态学报, 2002, 26(5): 589-593. |