 2015, Vol. 35
2015, Vol. 35文章信息
- 易雪梅, 张悦, 王远遐, 姬兰柱, 吴培莉
- YI Xuemei, ZHANG Yue, WANG Yuanxia, JI Lanzhu, WU Peili
- 长白山水曲柳种群动态
- Population structure and dynamics of Fraxinus mandshurica in Changbai Mountain
- 生态学报, 2015, 35(1): 91-97
- Acta Ecologica Sinica, 2015, 35(1): 91-97
- http://dx.doi.org/10.5846/stxb201403260566
-
文章历史
- 收稿日期:2014-03-26
- 修订日期:2014-11-17






2. 中国科学院大学, 北京 100049;
3. 吉林省露水河林业局宏伟种子园, 白山 134506
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Jilin province, Lushuihe Forestry Bureau, Hongwei seed orchard, Baishan 134506, China
植物种群濒危通常是种群内在特征与外界因素综合作用的结果[1]。一方面是内在的遗传因素、适应力、生活力等方面的缺陷导致生活史中某一环节极为脆弱,使得种群生存受到限制从而导致种群数量减少、分布区面积逐步缩小。另一方面则是由于外部环境条件剧烈变化,植物本身不能适应变化的环境而导致灭绝。近年来,应用保护生物学原理和手段对濒危植物进行研究越来越受到国内外学者的重视。种群是构成群落的基本单位,其结构不仅直接影响群落结构,而且能客观体现群落的发展、演变趋势。种群结构包括径级结构、高度结构以及年龄结构等[2]。物种种群结构的研究,能在对物种的种群现状和未来发展状况整体把握的前提下,对物种的进一步研究提供基础。
水曲柳(Fraxinus mandshurica P)为木犀科梣属(又称白蜡属)的植物。分布于朝鲜、日本、俄罗斯以及中国的陕西、甘肃、湖北、东北、华北等地,与黄菠萝、胡桃楸并称“东北三大硬阔”。水曲柳材质坚韧,纹理美观,可高达30m,是东北、华北地区的珍贵用材树种,可制各种家具、乐器、体育器具、车船、机械及特种建筑材料。同时,对于研究第三纪植物区系及第四纪冰川期气候具有科学意义。
研究水曲柳种群生态学特性,明确种群的结构与特征,对探讨未来环境条件变化对种群结构及动态的影响具有重要意义。目前对水曲柳的研究,主要涉及木材性能[3, 4, 5, 6]、立地造林[7, 8, 9, 10, 11]、种质资源[12, 13, 14]以及生理生态[10, 15, 16]等方面。对水曲柳的自然群落状态、种群结构以及相关濒危机制的研究相对缺乏。本文试图通过对水曲柳种群及种群动态预测的研究,对水曲柳的濒危现状进行评估并探讨其濒危原因,进而为水曲柳的保护提供科学依据。
1 研究地区与研究方法 1.1 研究区概况研究地点位于吉林省的东南部长白山自然保护区内,地跨延边朝鲜族自治州的安图县和浑江地区的抚松县、长白县。东南与朝鲜毗邻。地理位置为127°42′55″E—128°16′48″W,41°41′49″N—42°51′18″S,总面积为196465 hm2,森林覆盖率87.9%,是一个以森林生态系统为主要保护对象的自然综合体自然保护区。研究区气候属于受季风影响的大陆山地气候,气候受地势高低影响大,冬季漫长寒冷,夏季较短,温暖湿润。年平均温度3—7 ℃,年平均降水量700—1400 mm。土壤属于暗棕色森林土,植被以阔叶红松林与白桦次生林为主。阔叶红松林。阔叶红松林以红松、紫椴、蒙古栎、水曲柳和色木槭等为主要建群种,林分结构复杂,多形成复层混交林[17]。长白山白桦次生林是该地区地带性森林阔叶红松林经皆伐和火烧等破坏后恢复形成的主要次生林类型之一,是阔叶红松林次生演替系列中的重要阶段[18]。演替先锋树种白桦和山杨在群落中占有绝对优势地位[19]。
1.1.1 野外调查根据水曲柳在长白山区的分布,分别在阔叶红松林和白桦次生林两种生境对水曲柳自然种群进行调查。每种生境选取20 m×30 m样方36个,调查样方内胸径1 cm以上乔灌木。记录每株的树种、树高、冠幅、层盖度。同时测定样方环境因子:海拔、经纬度、坡度等,记录各样方的群落构成。每个样方内设置5 m×5 m样方,调查灌木层物种组成、胸径和树高,并在其中设置1 m×1 m样方调查草本成的物种组成情况。选择生长发育中等,具有代表性的水曲柳群体,统计其胸径。
1.1.2 种群径级划分和静态生命表编制依据相同环境下相同树种的龄级和径级对环境的反应规律具有一致性的特点[20],采用较为常用的径级代替龄级的方法来分析水曲柳的种群结构。根据调查的数据,结合水曲柳的生物学特性和生长情况,参考其他划分方法,将水曲柳种群按胸径划分为7个径级: 1—2 cm为第Ⅰ径级,2—5 cm为第Ⅱ径级,5—10 cm为第Ⅲ径级,10—20 cm为第Ⅳ径级,20—30 cm为第Ⅴ径级,30—40 cm为第Ⅵ径级。第Ⅰ径级对应第1龄级,依此类推。并以此为依据统计各径级水曲柳的个体数量。
在对所有样方内不同径级存活个体进行统计的基础上,按照静态生命表编制方法[21]编制长白山水曲柳种群生命表,用于种群结构分析。将水曲柳种群按不同生境分为阔叶红松林种群和白桦次生林种群。以径级代替龄级的研究由于具体生长环境的不同在具体结果上会有部分差异,本文着重于水曲柳种群的种群结构,对于不同林型不同生境的比较也是为了水曲柳总体分布的描述。对于水曲柳径级的比较主要集中在幼龄、壮龄和老龄,因而对两种生境选取了同样的径级分类。分别对两种生境种群编制静态生命表和总和静态生命表,并绘制存活曲线和死亡率曲线。以水曲柳径级为横坐标,标准化存活数为纵坐标分别绘制不同种群绘制水曲柳种群存活曲线。
1.1.3 种群生存及数量动态模拟在种群静态生命表的基础上,引入生存分析中的4个函数,即种群生存率函数S、积累死亡率函数F、死亡密度函数f和危险率函数[22],4个函数用以下公式计算:
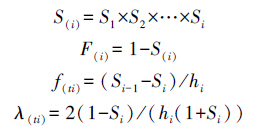
式中,Si为存活率,hi为龄级宽度。根据以上函数绘制生存率曲线、积累死亡率曲线、死亡密度曲线和危险率曲线。
采用时间序列平均推移法对水曲柳种群年龄结构进行预测[23],模型如下:
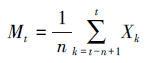
式中,n为预测时间;t为龄级;Mt为未来n年时t龄级的种群存活数;Xk为当前k龄级种群存活数。应用以上模型,以水曲柳种群总和各龄级植株株数为原始数据,按照一次平均推移法计算各龄级水曲柳在未来5、10a和20a后的株数,根据结果绘制年龄株数关系图。
2 结果与分析 2.1 水曲柳自然种群状况植被调查结果显示,阔叶红松林中的水曲柳植株个体较多,径级较大,而白桦次生林中植株个体相对较少,径级也较小。根据水曲柳在长白山的分布将调查到的水曲柳种群按生境不同分为阔叶红松林和白桦次生林两种,对两种生境进行分别统计。
种群年龄结构不仅反映了种群大小个体的组配情况,也有利于解释种群生存状况和繁殖策略[24]。由图 1可知,阔叶红松林中,第Ⅳ径级(胸径: 10—20 cm)的水曲柳植株在种群中所占比例最多,为28.7%。第Ⅰ、Ⅱ径级较少,所占比例都在10%以下。白桦次生林中,第Ⅳ径级(胸径:5—10 cm)的水曲柳植株个体也占据了总体的最多比例,为37%。第Ⅰ、Ⅶ径级个体较少,占种群的10%以下,分别为8.7%和7.2%。第Ⅷ径级,即胸径为30 cm以上的个体数为零。阔叶红松林中,第Ⅳ、Ⅴ、Ⅵ、Ⅶ径级为优势径级,其个体总数占总体的81.3%。白桦次生林中,第Ⅱ、Ⅲ、Ⅳ径级为白桦次生林中水曲柳种群中的主要组成个体,占据了种群的79.7%。

|
| 图 1 水曲柳种群径级结构 Fig.1 Size structure of Fraxinus mandshurica population 第Ⅰ径级: 2 cm以下(包括2cm);第Ⅱ径级: 2—5cm;第Ⅲ径级: 5—10 cm;第Ⅳ径级: 10—20 cm;第Ⅴ径级: 20—30 cm;第Ⅵ径级: 30—40 cm;Ⅶ径级: 40cm以上(包括40cm) |
水曲柳种群整体第Ⅳ径级占总体的比例最多,占32.6%。优势径级为第Ⅲ、Ⅵ、Ⅴ径级,占据了总体的67.7%。第Ⅰ、第Ⅶ径级占据水曲柳种群比例最少,均占总体的6.6%。
2.2 水曲柳静态生命表本研究调查的是长白山水曲柳的天然种群,由“空间推时间”、“横向导纵向”,因此所调查的数据并不完全满足编制特定时间表的3个假设[22]。因此,在生命表中出现死亡率为负的情况,这与数学假设不符,但仍能表明种群并非静止不动,而是在迅速的发展或衰退之中[25]。
由表 1可知,阔叶红松林中第Ⅱ、第Ⅲ径级死亡率(q)为负,白桦次生林中,第Ⅰ、第Ⅱ、第Ⅲ径级死亡率为负,主要原因是由于水曲柳种群中幼苗不足现象导致了这些径级为负值的死亡率。阔叶红松林中的第Ⅰ径级具有最高的死亡率,而在白桦次生林中从第Ⅳ径级依次升高,直到具最高死亡率的第Ⅶ径级。白桦次生林中水曲柳种群中从第Ⅳ径级升高的死亡率以及阔叶红松林中后期死亡率的升高都显示植株进入中龄后,各生境中的水曲柳种群存活率逐渐下降,此种现象可能是由于随着个体对资源环境的需求不断增大,种内和种间竞争导致的种群死亡率不断升高造成。
| 径级
Diameter class | 存活数a
Survival number | 标准化存
活数l Standard living number | 死亡数d
Mortality number | 死亡率q
Mortality rate | 平均存
活数L Average survival number | 总寿命T
Total life | 生命期望e
Mean expectation of life | 存活率s
Alive rate | |
| 阔叶红松林 | Ⅰ | 7 | 162.79 | 69.77 | 0.43 | 127.91 | 3406.98 | 20.93 | 0.57 |
| Korean pine | Ⅱ | 4 | 93.02 | -302.33 | -3.25 | 244.19 | 3279.07 | 35.25 | 4.25 |
| broadleaved | Ⅲ | 17 | 395.35 | -604.66 | -1.53 | 697.67 | 3034.88 | 7.68 | 2.53 |
| forest | Ⅳ | 43 | 1000.00 | 232.56 | 0.23 | 883.72 | 2337.21 | 2.34 | 0.77 |
| Ⅴ | 33 | 767.44 | 139.53 | 0.18 | 697.67 | 1453.49 | 1.89 | 0.82 | |
| Ⅵ | 27 | 627.91 | 186.05 | 0.30 | 534.88 | 755.81 | 1.20 | 0.70 | |
| Ⅶ | 19 | 441.86 | 441.86 | 1 | 220.93 | 220.93 | 0.5 | 1 | |
| 白桦次生林 | Ⅰ | 12 | 235.29 | -215.69 | -0.92 | 343.14 | 2588.23 | 11 | 1.92 |
| Betula | Ⅱ | 23 | 450.98 | -254.90 | -0.57 | 578.43 | 2245.10 | 4.98 | 1.57 |
| platyphylla | Ⅲ | 36 | 705.88 | -294.12 | -0.42 | 852.94 | 1666.67 | 2.36 | 1.42 |
| secondary | Ⅳ | 51 | 1000 | 705.88 | 0.71 | 647.06 | 813.73 | 0.81 | 0.29 |
| forest | Ⅴ | 15 | 294.12 | 274.51 | 0.93 | 156.86 | 166.67 | 0.57 | 0.07 |
| Ⅵ | 1 | 19.61 | 19.61 | 1 | 9.80 | 9.80 | 0.5 | 0 | |
| Ⅶ | 0 | 0 | 0 | 0 | 1 | ||||
| 种群总和 | Ⅰ | 19 | 202.13 | -85.11 | -0.42 | 244.68 | 2962.77 | 14.66 | 1.42 |
| Total | Ⅱ | 27 | 287.23 | -276.60 | -0.96 | 425.53 | 2718.09 | 9.46 | 1.96 |
| population | Ⅲ | 53 | 563.83 | -329.79 | -0.58 | 728.72 | 2292.55 | 4.07 | 1.58 |
| Ⅳ | 84 | 893.62 | 382.98 | 0.43 | 702.13 | 1563.83 | 1.75 | 0.57 | |
| Ⅴ | 48 | 510.64 | 106.38 | 0.21 | 457.45 | 861.70 | 1.69 | 0.79 | |
| Ⅵ | 38 | 404.26 | 202.13 | 0.5 | 303.19 | 404.26 | 1 | 0.5 | |
| Ⅶ | 19 | 202.13 | 202.13 | 1 | 101.06 | 101.06 | 0.5 | 1 |
阔叶红松林中,第Ⅱ径级生命期望最高,而在白桦次生林和种群总和中,第Ⅰ径级具有最高的生命期望值。整个水曲柳种群第Ⅰ、第Ⅱ、第Ⅲ径级死亡率为负,第Ⅱ径级具有最高的存活率。以上数据表明,阔叶红松林和白桦次生林中生命质量最高的分别为第Ⅱ径级和第Ⅰ径级。在整个水曲柳种群中,第Ⅰ径级拥有最高的生命质量。
2.3 水曲柳种群生存力 2.3.1 水曲柳种群存活曲线种群存活曲线描绘群体个体在各龄级的存活情况,反映种群的重要动态特征[22]。从图 2可看出,水曲柳的存活曲线基本呈从图 2可看出,水曲柳的存活曲线基本呈Deevey Ⅲ 型。在阔叶红松林种群、白桦次生林种群和种群总和中,水曲柳种群都呈正态分布,幼龄比例较少,而中老龄占据大部分。存活曲线显示,水曲柳种群个体储备比较不足,更新能力弱,呈衰退趋势。

|
| 图 2 水曲柳种群存活曲线 Fig.2 Survive curve of Fraxinus mandshurica population |
由图 3可知,阔叶红松林中水曲柳第Ⅱ、第Ⅲ径级的死亡率为负数,第Ⅰ径级死亡率最高。白桦次生林中水曲柳种群与种群总和一样,第Ⅵ径级具有最高的死亡率。由水曲柳种群总和的死亡率曲线可知,水曲柳种群尤其是阔叶红松林中水曲柳种群幼株缺乏,死亡率随径级增加而增加,随着中龄植株生存能力的增加,其死亡率逐渐平缓,一直到老龄植株之后死亡率达到最高。

|
| 图 3 水曲柳种群死亡率曲线 Fig.3 Mortality curve of Fraxinus mandshurica population |
由表 2可知,第Ⅱ、第Ⅲ径级水曲柳在阔叶红松林、白桦次生林中都具有相对较高的种群生存率,白桦次生林中第Ⅰ径级水曲柳的种群生存率最高。阔叶红松林中,第Ⅲ、第Ⅳ径级水曲柳植株积累死亡率较高,白桦次生林中第Ⅱ、第Ⅲ径级植株积累死亡率较高,而种群总和中,第Ⅲ径级水曲柳植株具有最高的积累死亡率。阔叶红松林、白桦次生林以及种群总和中水曲柳种群死亡密度最大值均出现在第Ⅰ径级。
|
径级
Diameter class | 阔叶红松林
Korean pine broadleaved forest | 白桦次生林
Betula platyphylla secondary forest | 种群总和
Total population | |||||||||
| S | F | f | λ | S | F | f | λ | S | F | f | λ | |
| S、F、f、λ分别为各龄级的种群生存率 Survival rate、积累死亡率Cumulative death rate、死亡密度Mortality density和危险率Hazard rate | ||||||||||||
| Ⅰ | 0.57 | 0.43 | 0.43 | 0.55 | 1.92 | -0.92 | 1.92 | -0.63 | 1.42 | -0.42 | 1.42 | -0.35 |
| Ⅱ | 2.43 | -1.4 | -1.84 | -0.42 | 3 | -2 | 0.54 | -0.5 | 2.79 | -1.79 | 0.68 | -0.47 |
| Ⅲ | 6.14 | -5.14 | 0.57 | -0.48 | 4.25 | -3.25 | 0.42 | -0.41 | 4.42 | -3.42 | 0.54 | -0.42 |
| Ⅳ | 4.71 | -3.71 | 0.06 | -0.33 | 1.25 | -0.25 | -0.75 | -0.06 | 2.53 | -1.53 | -0.47 | -0.22 |
| Ⅴ | 3.86 | -2.86 | -0.01 | -0.24 | 0.08 | 0.92 | -0.23 | 0.34 | 2 | -1 | -0.11 | -0.13 |
| Ⅵ | 2.71 | -1.71 | 0.02 | -0.15 | 0 | 1 | -0.01 | 0.33 | 1 | 0 | -0.17 | 0 |
| Ⅶ | 0 | 1 | 0.14 | 0.29 | 0 | 1 | 0 | 0.29 | 0 | 1 | -0.2 | 0.29 |
由图 4可知,阔叶红松林中,第Ⅰ径级水曲柳具有最高的危险率,而白桦次生林以及种群总和中的水曲柳种群分别在第Ⅴ和第Ⅶ径级危险率最高。阔叶红松林和白桦次生林中,水曲柳种群危险率最低的径级分别为第Ⅲ和第Ⅰ径级。从第Ⅱ径级开始,水曲柳种群的危险率逐步上升。

|
| 图 4 水曲柳种群危险率曲线 Fig.4 Hazard rate curve of Fraxinus mandshurica population |
由图 5可知,各龄级植株株数随时间推移,幼龄植株减少,老龄植株增多。水曲柳种群幼龄植株逐渐缺失,种群进入老龄化,老龄个体增多,最终导致整个种群的衰退。可以预测,如果不对水曲柳种群的幼苗更新及补充尽快采取有效措施,水曲柳种群的稳定性将很难维持。

|
| 图 5 水曲柳种群数量动态时间预测 Fig.5 Time series prediction of dynamics of Fraxinus mandshurica population |
生态系统种群结构直接影响群落结构,并能体现群落发展演替趋势。种群结构反映了种群个体在空间上的配置、数量动态和发展趋势,同时也体现了种群与环境之间的相互关系及其在群落中的地位和作用[26]。种群的静态生命表和生存函数分析反映种群不同生活史阶段的动态特征及其与环境之间的相互关系,可作为种群保护和经营管理的和总要参考。经营管理者能够针对不同阶段种群的生长发育特点采取适当的经营和保护措施,实现种群的可持续发展。濒危植物的大多数种群年龄结构呈现衰退趋势[1]。如: 银杉、鹅掌楸,绝大多数种群的年龄结构呈现幼龄个体少,老龄个体多的特点,种群扩展缺乏足够的幼龄个体[27, 28]。
长白山水曲柳种群年龄结构、静态生命表和种群数量动态预测表明,水曲柳种群存活曲线为Deevey Ⅰ型,属于典型的衰退型。生命表各参数表明,整个水曲柳种群呈现为老龄个体多,幼龄个体少的衰退型种群结构,种群目前主要靠中壮龄个体维持,种群的持续发展需要相当数量的幼龄个体补充。
3.2 水曲柳的濒危现状种群衰退通常有两个推动因子:一是种群补充的缺乏,二是种群死亡的增加[29]。植物种群的减少,一方面是由于幼苗的供给不足,一方面是已成活个体的死亡。
水曲柳属落叶高大乔木,雌雄异株,果实为翅果,种子主要靠风力传播[30]。阔叶红松林与白桦次生林是水曲柳的主要分布生境,在两种生境中,均有大量幼苗出现,但水曲柳在25 hm2样地内仅有681棵胸径1 cm以上的个体,这可能与这些树种的生活史对策有关,种子和新增幼苗数量并不是水曲柳种群更新的限制因素[31]。由于光照、营养等因素限制造成的高死亡率造成水曲柳幼株补充不足,水曲柳种群幼龄个体较少,种群主要由中龄个体维持,整个种群呈衰退趋势。
3.3 水曲柳的保护水曲柳已被列为国家重点保护树种,但仍然受到采伐带来的威胁。与此同时,森林面积的急剧减少,也是水曲柳种群数量减少的重要因素。人类的过度开发利用以及水曲柳自身的更新限制,造成了水曲柳种群的进一步缩小。
鉴于目前水曲柳种群幼株严重缺少的现状,建议立足现有种群生存的维持保护上,保证并提高森林分布面积。从水曲柳自身更新生物学方面,结合外部环境条件研究水曲柳濒危机制。此外,应减少人为采伐利用,针对生态系统各演替阶段的生物多样性保护对自然保护区采取适当人工措施,如适度干扰,维持森林生态系统的多样性。
| [1] | 张文辉, 祖元刚, 刘国彬. 十种濒危植物的种群生态学特征及致危因素分析. 生态学报, 2002, 22(9): 1512-1520. |
| [2] | Holeksa J, Saniga M, Szwagrzyk J, Dziedzic T, Ferenc S, Wodka M. Altitudinal variability of stand structure and regeneration in the subalpine spruce forests of the Pol'ana biosphere reserve, Central Slovakia. European Journal of Forest Research, 2007, 126(2): 303-313. |
| [3] | 张燕, 佟达, 宋魁彦. 水热-微波处理水曲柳顺纹压缩应力-应变本构关系. 南京林业大学学报: 自然科学版, 2013, 37(4): 105-109. |
| [4] | 刘顺治, 林金国, 林玉明, 柯丽丽, 胡吉萍. 4 种常用装饰装修木材表面特性分析. 福建林业科技, 2013, 40(3): 54-56. |
| [5] | 李涛, 顾炼百. 185℃高温热处理对水曲柳木材力学性能的影响. 林业科学, 2009, 45(2): 92-97. |
| [6] | Sun Y L, Lu Z Y, He L. Studies on stress strength factors at crack tip of Fraxinus mandshurica rupr. Wood. Forestry Studies in China, 2000, 2(2): 76-79. |
| [7] | 吴楚, 王政权, 范志强, 孙海龙. 氮胁迫对水曲柳幼苗养分吸收、利用和生物量分配的影响. 应用生态学报, 2004, 15(11): 2034-2038. |
| [8] | 吴俊民, 礼波宁, 刘广平, 王晓水, 吴保国. 混交林中落叶松挥发性物质对水曲柳生长的影响. 东北林业大学学报, 2000, 28(1): 25-28. |
| [9] | 周亚茹, 何子兵, 费晓光. 东北珍贵绿化树种水曲柳苗木培育技术. 防护林科技, 2012, (1): 124-124, 126-126. |
| [10] | 陈海波, 卫星, 王婧, 王政权. 水曲柳苗木根系形态和解剖结构对不同氮浓度的反应. 林业科学, 2010, 46(2): 61-66. |
| [11] | 刘婷岩, 张彦东, 彭红梅, 甘秋妹. 林分密度对水曲柳人工幼龄林植被碳储量的影响. 东北林业大学学报, 2012, 40(6): 1-4. |
| [12] | 林士杰, 张忠辉, 谢朋, 张大伟, 李利军, 候丽伟, 田宇, 朱红波. 中国水曲柳基因资源的保护与利用. 中国农学通报, 2009, 25(24): 158-162. |
| [13] | 张丽杰. 取材时期对水曲柳合子胚外植体体细胞胚胎发生的影响[D]. 哈尔滨: 东北林业大学, 2006. |
| [14] | 刘立波, 王清君, 张志环, 赵科研, 张庆发. 水曲柳、胡桃楸等天然次生林生长情况调查. 林业科技开发, 2012, 26(1): 50-54. |
| [15] | 柏广新, 张彦东. 水曲柳天然更新及其影响因子. 东北林业大学学报, 2013, 41(1): 7-9, 13. |
| [16] | 战伟, 沙伟, 王淼, 师帅, 问青春. 降水和温度变化对长白山地区水曲柳幼苗生长和光合参数的影响. 应用生态学报, 2012, 23(3): 617-624. |
| [17] | 郝占庆, 郭水良, 曹同. 长白山植物多样性及其格局. 沈阳: 辽宁科学技术出版社, 2002. |
| [18] | 徐化成. 中国红松天然林. 北京: 中国林业出版社, 2001. |
| [19] | 郝占庆, 张健, 李步杭, 叶吉, 王绪高, 姚晓琳. 长白山次生杨桦林样地: 物种组成与群落结构. 植物生态学报, 2008, 32(2): 251-261. |
| [20] | Frost I, Rydin H. Spatial pattern and size distribution of the animal-dispersed tree Quercus robur in two spruce-dominated forests. Ecoscience, 2000, 7(1): 38-44. |
| [21] | 孙儒泳. 普通生态学. 北京: 高等教育出版社, 1993. |
| [22] | 于大炮, 周莉, 董百丽, 代力民, 王庆礼. 长白山北坡岳桦种群结构及动态分析. 生态学杂志, 2004, 23(5): 30-34. |
| [23] | 肖宜安, 何平, 李晓红, 邓洪平. 濒危植物长柄双花木自然种群数量动态. 植物生态学报, 2004, 28(2): 252-257. |
| [24] | 张文辉, 许晓波, 周建云, 谢宗强. 濒危植物秦岭冷杉种群数量动态. 应用生态学报, 2005, 16(10): 1799-1804. |
| [25] | 江洪. 云杉种群生态学. 北京: 中国林业出版社, 1992. |
| [26] | 康华靖, 陈子林, 刘鹏, 郝朝运, 韦福民. 大盘山自然保护区香果树种群结构与分布格局. 生态学报, 2007, 27(1): 389-396. |
| [27] | 谢宗强, 陈伟烈, 路鹏, 胡东. 濒危植物银杉的种群统计与年龄结构. 生态学报, 1999, 19(4): 523-528. |
| [28] | 魏勇, 田如男, 方炎明. 龙王山鹅掌楸种群的静态生命表. 南京林业大学学报: 自然科学版, 2004, 28(6): 61-63. |
| [29] | Yoshida T, Kamitani T. Growth of a shade-intolerant tree species, Phellodendron amurense, as a component of a mixed-species coppice forest of central Japan. Forest Ecology and Management, 1999, 113(1): 57-65. |
| [30] | 刘彤, 胡林林, 郑红, 周志强. 天然东北红豆杉土壤种子库研究. 生态学报, 2009, 29(4): 1869-1876. |
| [31] | 张健, 李步杭, 白雪娇, 原作强, 王绪高, 叶吉, 郝占庆. 长白山阔叶红松林乔木树种幼苗组成及其年际动态. 生物多样性, 2009, 17(4): 385-396. |