 2015, Vol. 35
2015, Vol. 35文章信息
- 张悦, 易雪梅, 王远遐, 姬兰柱, 吴培莉
- ZHANG Yue, YI Xuemei, WANG Yuanxia, JI Lanzhu, WU Peili
- 采伐对红松种群结构与动态的影响
- Impact of tree harvesting on the population structure and dynamics of Pinus koraiensis (Pinaceae)
- 生态学报, 2015, 35(1): 38-45
- Acta Ecologica Sinica, 2015, 35(1): 38-45
- http://dx.doi.org/10.5846/stxb201403250544
-
文章历史
- 收稿日期:2014-03-25
- 修订日期:2014-11-14






2. 中国科学院大学, 北京 100049;
3. 吉林省露水河林业局宏伟种子园, 白山 134506
2. University of Chinese Academy of Sciences, Beijing 100049, China;
3. Jilin Province, Lushuihe Forestry Bureau, Hongwei Seed Orchard, Baishan 134506, China
森林作为陆地最大的生态系统,不仅能为人类提供木材,而且在保护环境、涵养水分、防止水土流失等方面都发挥着重要的作用。然而大量的森林采伐使得森林资源锐减、质量下降,关于森林采伐所带来的各种影响,一直受到国内外学者的积极关注[1, 2, 3, 4]。森林采伐迹地植物种群动态变化规律是森林开发利用、植被改造或重建、生物多样性保护的重要依据[5, 6]。植物种群数量动态是植物个体生存能力与外界环境相互作用的结果[7, 8]。研究植物种群的年龄结构、存活曲线和生命表不仅可以反映种群生存现状[8],还可以展现植物种群与环境适应的结果,对阐明群落未来发展趋势也具有重要意义[7, 9, 10, 11, 12, 13]。
红松(Pinus koraiensis)是我国长白山阔叶红松林的主要建群种。近百年来,历经3次过度采伐,大范围的原始阔叶红松林遭到破坏[14],红松已成为我国稀有物种,被列为国家二级保护植物[15]。20世纪90年代,有关学者报道了小兴安岭天然红松林、长白山白桦林、云冷杉阔叶红松林、原始阔叶红松林内的红松种群数量变化的波动性、种群结构、更新格局动态和趋势等[16, 17, 18, 19, 20, 21],以期了解不同群落类型中红松种群的生存现状。近年来,有关采伐干扰对阔叶红松林中植物种群的影响已有少量报道。王惠等[22]分析了采伐干扰下长白山阔叶红松林主要灌木种群在伐后不同年度资源维上的生态位动态特征;齐麟等[23]研究了4种不同强度的采伐对白桦和红松的重要值以及幼树(胸径<10cm)占种群数量的比例的影响;倪瑞强[24]研究了原始阔叶红松林及其次生林在不同演替阶段群落内,红松、紫椴(Tilia amurensis)、色木槭(Acer mono)等林中主要树种的径级结构。然而,不同森林经营方式(如择伐和皆伐)、不同森林经营强度(如轻度择伐、中度择伐)对红松种群影响的研究亟需展开,结合红松种群结构和动态的研究方法,从年龄结构、静态生命表、存活曲线、生存函数等的变化规律,阐述采伐干扰对红松种群的影响,可以为长白山阔叶红松林的经营管理和红松种群的恢复提供理论依据。
1 研究地区与研究方法 1.1 研究地概况研究地在吉林省露水河林业局东升林场(42°24′—42°49′N,127°29′—128°02′E),位于长白山西北麓,平均海拔700m;属于温带季风大陆性山地气候[25],年平均气温为3.6℃,最冷月(1月份)平均气温为-15.6℃,最热月(7月份)平均气温为19.7℃;年平均降水量约700mm,且主要集中在7—9月,在480—500mm之间[26, 27]。土壤主要亚类有白浆暗棕壤、典型暗棕壤和暗色暗棕壤[28]。地带性植被为阔叶红松林,分布于海拔500—1100 m的玄武岩台地上[29]。阔叶红松林群落类型以红松、紫椴、蒙古栎(Quercus mongolica)、水曲柳(Fraxinus mandshurica)和色木槭等为主要建群种,林分结构复杂,多形成复层混交林[25]。
1.2 研究方法 1.2.1 样地设置与样地调查露水河林业局1965年建局,主要从事木材和锯材生产,20世纪80年代以前采伐方式为大面积皆伐,后改为采育兼顾伐和径级伐[30]。根据《露水河林业局森林经营方案》,1979年在露水河东升林场的阔叶红松林采取过小面积皆伐,1982年对红松种群采取过15%和40%择伐强度的采伐作业,而东升林场的21林班作为采伐作业的保留地,保持了原始阔叶红松林的原貌。本研究选取露水河东升林场的四类林区,即原始阔叶红松林(T0)、15%择伐林(T1)、40%择伐林(T2)、皆伐后恢复的天然次生林(T3)。每类林区分别选取2个林班,在每个林班内分别设置3个30m×30m的样方,每个样方内各设置5个5m×5m的小样方。对样方内DBH≥ 2.5cm的乔木树种进行每木检尺,记录乔木种名、胸径(DBH)、树高(H);小样方内DBH<2.5cm的幼树、幼苗,记录株数和树高,样地内的数值由小样方换算而来。记录样地的生境条件、植被状况及土壤条件。
1.2.2 发育阶段的划分方法由于乔木树种的年龄较难确定,所以在实际工作中采用发育阶段代替龄级的方法,具体划分结果如下:
Ⅰ级幼苗H<0.33m;Ⅱ级幼树H≥0.33m,DBH<2.5cm;Ⅲ级小树2.5cm≤DBH<7.5cm;Ⅳ级中树7.5cm≤DBH<12.5cm;以后DBH每隔5cm为1级,DBH≥22.5cm为大树。把树木H和DBH由小到大的顺序看作是时间顺序关系,Ⅰ级对应第1发育阶段,Ⅱ级对应第2发育阶段级,以此类推,统计各龄级株数。
1.2.3 生命表的编制方法静态生命表一般包括如下内容:x是单位时间发育等级的中值;ax是在x发育阶段内现有个体数;lx是在x发育阶段开始时标准化存活个体数;dx是从x到x+1发育阶段间隔期内标准化死亡数;qx是从x到x+1发育阶段间隔期间死亡率;Lx是从x到x+1发育阶段间隔期还存活的个体数;Tx是从x发育阶段到超过x发育阶段的个体总数;ex是进入x发育阶段个体的生命期望或平均期望寿命;Kx为消失率(损失度)。表中各项都是相互关联的,可以通过实测值求得,其关系如下:
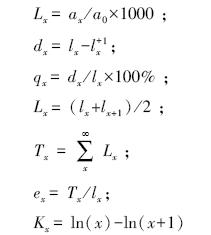
由于静态生命表是反映了多个世代重叠的发育阶段动态历程中的一个特定时间,而不是对这一种群的全部生活是追踪,并且调查中存在系统误差,在生命表中会出现死亡率为负的情况,因此本研究采用匀滑技术[31]进行处理,对ax进行匀滑修正得到a′x,据此编制红松种群静态生命表。
1.2.4 生存分析的4个函数计算方法为更好地分析红松种群的结构形式,阐明其生存规律,引入生存分析中的4个函数项目于生命表中,即生存率函数S(t)、积累死亡率函数F(t)、死亡密度函数f(t)、危险率函数λ(t)[32, 33, 34]:

式中,Pi为存活频率。

式中,hi为区间长度,qi为死亡频率。
2 结果与分析 2.1 红松种群生命表及4个函数生存分析
根据红松种群调查资料,把种群分为18个发育阶段,按照静态生命表的编制方法和生存理论,分别以原始林和3个采伐梯度的天然次生林的5400m2样地的调查数据,编制出红松种群静态生命表(表 1)和4个函数的估算值(表 2)。限于篇幅,本文仅列出了生命表的部分项目。
| 龄级 | x | 原始林(T0) | 15%择伐林(T1) | 40%择伐林(T2) | 皆伐林(T3) | ||||||||||||
| a′x | qx | Kx | ex | a′x | qx | Kx | ex | a′x | qx | Kx | ex | a′x | qx | Kx | ex | ||
| x,ax,qx,Kx,ex分别为径级中值、在x龄级内出现的个体数、死亡率、致死力和平均生命期望 | |||||||||||||||||
| 1 | 0 | 47 | 0.468 | 0.631 | 2.703 | 43 | 0.488 | 0.669 | 3.115 | 62 | 0.435 | 0.571 | 2.436 | 102 | 0.412 | 0.531 | 1.795 |
| 2 | 2 | 25 | 0.600 | 0.915 | 3.641 | 22 | 0.637 | 1.013 | 4.654 | 35 | 0.685 | 0.155 | 2.956 | 60 | 0.299 | 0.356 | 1.702 |
| 3 | 5 | 10 | 0.103 | 0.109 | 7.343 | 8 | 0.000 | 0.000 | 10.935 | 11 | 0.000 | 0.000 | 7.197 | 42 | 0.430 | 0.561 | 1.216 |
| 4 | 10 | 9 | 0.110 | 0.116 | 7.131 | 8 | 0.000 | 0.000 | 9.935 | 11 | 0.090 | 0.094 | 6.197 | 24 | 0.749 | 1.382 | 0.753 |
| 5 | 15 | 8 | 0.124 | 0.132 | 6.947 | 8 | 0.124 | 0.132 | 8.935 | 10 | 0.099 | 0.104 | 5.759 | 6 | 0.169 | 4.078 | 0.508 |
| 6 | 20 | 7 | 0.141 | 0.152 | 6.852 | 7 | 0.000 | 0.000 | 9.123 | 9 | 0.110 | 0.116 | 5.336 | — | — | — | — |
| 7 | 25 | 6 | 0.000 | 0.000 | 6.891 | 7 | 0.000 | 0.000 | 8.123 | 8 | 0.123 | 0.131 | 4.931 | — | — | — | — |
| 8 | 30 | 6 | 0.000 | 0.000 | 5.891 | 7 | 0.141 | 0.152 | 7.123 | 7 | 0.140 | 0.151 | 4.53 | — | — | — | — |
| 9 | 35 | 6 | 0.164 | 0.179 | 4.891 | 6 | 0.000 | 0.000 | 7.207 | 6 | 0.163 | 0.178 | 4.214 | — | — | — | — |
| 10 | 40 | 5 | 0.000 | 0.000 | 4.748 | 6 | 0.000 | 0.000 | 6.207 | 5 | 0.000 | 0.000 | 3.939 | — | — | — | — |
| 11 | 45 | 5 | 0.196 | 0.218 | 3.748 | 6 | 0.164 | 0.179 | 5.207 | 5 | 0.195 | 0.217 | 2.939 | — | — | — | — |
| 12 | 50 | 4 | 0.244 | 0.280 | 3.535 | 5 | 0.000 | 0.000 | 5.128 | 4 | 0.242 | 0.278 | 2.530 | — | — | — | — |
| 13 | 55 | 3 | 0.000 | 0.000 | 3.508 | 5 | 0.000 | 0.000 | 4.128 | 3 | 0.340 | 0.416 | 2.180 | — | — | — | — |
| 14 | 60 | 3 | 0.000 | 0.000 | 2.508 | 5 | 0.197 | 0.219 | 3.128 | 2 | 0.000 | 0.000 | 2.030 | — | — | — | — |
| 15 | 65 | 3 | 0.338 | 0.413 | 1.508 | 4 | 0.000 | 0.000 | 2.766 | 2 | 0.185 | 0.663 | 1.030 | — | — | — | — |
| 16 | 70 | 2 | 0.488 | 0.670 | 1.023 | 4 | 0.245 | 0.281 | 1.766 | 1 | 1.000 | 0.833 | 0.529 | — | — | — | — |
| 17 | 75 | 1 | 1.000 | 3.091 | 0.500 | 3 | 0.338 | 0.413 | 1.169 | — | — | — | — | — | — | — | — |
| 18 | 80 | — | — | — | — | 2 | 1.000 | 3.850 | 0.511 | — | — | — | — | ||||
| 龄级 | x | 原始林(T0) | 15%择伐林(T1) | 40%择伐林(T2) | 皆伐林(T3) | ||||||||||||
| Sx | Fx | fx | λx | Sx | Fx | fx | λx | Sx | Fx | fx | λx | Sx | Fx | fx | λx | ||
| x,S,F,f,λ分别为径级中值、在x龄级内的生存函数、累计死亡率函数、死亡密度函数、危险率函数 | |||||||||||||||||
| 1 | 0 | 0.532 | 0.468 | 0.106 | 0.200 | 0.512 | 0.488 | 0.102 | 0.200 | 0.565 | 0.435 | 0.113 | 0.200 | 0.588 | 0.412 | 0.118 | 0.200 |
| 2 | 2 | 0.213 | 0.787 | 0.064 | 0.300 | 0.186 | 0.814 | 0.065 | 0.351 | 0.178 | 0.822 | 0.077 | 0.435 | 0.412 | 0.588 | 0.135 | 0.085 |
| 3 | 5 | 0.191 | 0.809 | 0.004 | 0.023 | 0.186 | 0.814 | 0.000 | 0.000 | 0.178 | 0.822 | 0.000 | 0.000 | 0.235 | 0.765 | 0.035 | 0.151 |
| 4 | 10 | 0.17 | 0.830 | 0.004 | 0.025 | 0.186 | 0.814 | 0.000 | 0.000 | 0.162 | 0.838 | 0.003 | 0.020 | 0.059 | 0.941 | 0.035 | 0.060 |
| 5 | 15 | 0.149 | 0.851 | 0.004 | 0.028 | 0.163 | 0.837 | 0.005 | 0.028 | 0.146 | 0.854 | 0.003 | 0.022 | 0.000 | 1.000 | 0.012 | — |
| 6 | 20 | 0.128 | 0.872 | 0.000 | 0.033 | 0.163 | 0.837 | 0.000 | 0.000 | 0.130 | 0.870 | 0.003 | 0.025 | — | — | — | — |
| 7 | 25 | 0.128 | 0.872 | 0.000 | 0.000 | 0.163 | 0.837 | 0.000 | 0.000 | 0.114 | 0.886 | 0.003 | 0.028 | — | — | — | — |
| 8 | 30 | 0.128 | 0.872 | 0.004 | 0.000 | 0.140 | 0.860 | 0.005 | 0.033 | 0.098 | 0.902 | 0.003 | 0.033 | — | — | — | — |
| 9 | 35 | 0.107 | 0.893 | 0.000 | 0.039 | 0.140 | 0.860 | 0.000 | 0.000 | 0.082 | 0.918 | 0.003 | 0.039 | — | — | — | — |
| 10 | 40 | 0.107 | 0.893 | 0.004 | 0.000 | 0.140 | 0.860 | 0.000 | 0.000 | 0.082 | 0.918 | 0.000 | 0.000 | — | — | — | — |
| 11 | 45 | 0.086 | 0.914 | 0.004 | 0.049 | 0.117 | 0.883 | 0.005 | 0.039 | 0.066 | 0.934 | 0.003 | 0.048 | — | — | — | — |
| 12 | 50 | 0.065 | 0.935 | 0.000 | 0.065 | 0.117 | 0.883 | 0.000 | 0.000 | 0.050 | 0.950 | 0.003 | 0.064 | — | — | — | — |
| 13 | 55 | 0.065 | 0.935 | 0.000 | 0.000 | 0.117 | 0.883 | 0.000 | 0.000 | 0.003 | 0.967 | 0.003 | 0.103 | — | — | — | — |
| 14 | 60 | 0.065 | 0.935 | 0.004 | 0.000 | 0.094 | 0.906 | 0.005 | 0.049 | 0.003 | 0.967 | 0.000 | 0.000 | — | — | — | — |
| 15 | 65 | 0.043 | 0.957 | 0.004 | 0.102 | 0.094 | 0.906 | 0.000 | 0.000 | 0.017 | 0.983 | 0.003 | 0.188 | — | — | — | — |
| 16 | 70 | 0.022 | 0.978 | 0.004 | 0.191 | 0.071 | 0.929 | 0.005 | 0.065 | 0.000 | 1.000 | 0.003 | — | — | — | — | — |
| 17 | 75 | 0.000 | 1.000 | 0.004 | — | 0.147 | 0.953 | 0.005 | 0.102 | — | — | — | — | — | — | — | — |
| 18 | 80 | — | — | — | — | 0.000 | 1.000 | 0.009 | — | — | — | — | — | ||||
从表 1中可以看出,红松是长寿命物种,样地中调查到的最高龄级为18发育阶段。从种群的死亡率(qx)和致死率(Kx)来看,第1,2发育阶段幼苗的死亡率和致死率最高,第3,4发育阶段相差不大,第5,6发育阶段保持较低水平的死亡率,在7,8,9发育阶段出现波动,死亡率升高,此后高发育阶段保持较高的死亡率,达到生理年限附近,死亡率达到最大。采伐干扰对红松种群的波动周期影响不大。原始林和择伐林(T0、T1、T2)与皆伐后次生林(T3)相比,林下红松幼苗、幼树的数量普遍偏少,匀滑后幼苗、幼树的数量在个别不同的龄级相似,死亡率为零。择伐内红松约占60%以上,种群世代重叠严重,高发育阶段个体死亡率水平偏高,但个别高发育阶段出现死亡率为零的现象;林下幼龄株数稀少,种群更新不良。皆伐后恢复30余年的天然次生林内,红松种群生长到第5发育阶段,幼苗、幼树的死亡率均较高。
平均生命期望( ex ) 反映的是x发育阶段内个体的平均生存能力。从表 1可以看出,皆伐后恢复的天然次生林内红松幼苗数量比原始林和择伐林多,但生命期望值比母树林的幼苗低,原始林和择伐林内(T0、T1、T2)内红松第1—5发育阶段的生命期望为 2.4—10.9,而皆伐林(T3)为 0.5—1.8。从一个世代周期的生命期望来看,各群落内红松种群的生命期望排序:15%择伐林(T1)>原始林(T0)>40%择伐林(T2),说明15%的择伐强度可以提高红松种群的生存期望。
从4个函数生存分析(表 2)结果来看,原始林和择伐林(T0、T1、T2)内红松第二发育阶段的危险率(λx)(0.30—0.43)最高,皆伐林内(T3)第一龄级的危险率(0.2)最高,皆伐干扰对红松种群危险率的影响最大。原始林(T0)、15%择伐林(T1)、40%择伐林(T2)内红松累计死亡率(Fx)分别在第16、18、15发育阶段达到0.98以上,15%的择伐强度的采伐降低了红松高发育阶段个体的死亡速率。
2.2 种群存活曲线分析以红松种群生命表中存活量的对数ln(lx)为纵坐标,以龄级为横坐标,分别绘制红松不同种群存活曲线(图 1)。从图 1可见,母树林(T0、T1、T2)在第1—3发育阶段存活曲线的斜率最大,第3—10发育阶段保持平稳,而皆伐林在第1—5发育阶段存活曲线的斜率较大,皆伐干扰对红松幼苗时期的存活量影响最大。第11发育阶段以后,随着择伐强度的增大,存活曲线的斜率逐渐增大,种群存活量逐渐增大。

|
| 图 1 红松种群在不同采伐强度群落中的存活曲线 Fig. 1 Survival curve of P. koraiensis population |
采用指数方程和幂函数方程来描述存活曲线,检验估算的存活状况是否符合DeeveyⅡ型还是符合DeeveyⅢ型,而DeeveyⅠ型则符合直线分布。对4个存活曲线进行模型拟合,结果表明,原始林(T0)和15%择伐林(T1)内红松存活曲线均为DeeveyⅡ型,除去幼苗阶段死亡率较高外,其它发育阶段死亡率相当,种群处在稳定期。40% 择伐林(T2)内,红松存活曲线介于DeeveyⅡ型和DeeveyⅢ型之间,群落内高发育阶段红松个体偏多,种群正在由稳定期向成熟期过渡。皆伐林(T3)内存活曲线为DeeveyⅠ型,种群处在增长期。随着采伐强度增大,采伐干扰对种群结构影响也增大。
2.3 种群年龄结构及方差分析根据红松的径级组中值和株数百分数构建种群径级结构图(图 2)。从图 2可见,原始林和择伐林(T0、T1、T2)内种群径级结构均呈稳定的倒J型;第10—30发育阶段幼树比相邻发育阶段的株数百分比偏少,该阶段的幼树处于生长更新的停滞阶段,这与野外调查中发现的“只见红松幼苗,不见幼树”相符。与原始阔叶红松林相比,15%择伐林内幼树(DBH<10cm)比例略有减少,而40%择伐林和皆伐迹地,随采伐强度的增加,幼树比例明显增大。

|
| 图 2 红松种群径级分布 Fig. 2 Diamerter class distribution of P. koraiensis population |
对红松不同发育阶段的存活量(lx)及其在不同群落类型中的存活量(lx)进行方差分析(表 3),结果表明,红松种群不同发育阶段组间生存状况存在很大差异(P<0.001),种群在不同发育阶段间的生存方式明显不同。红松种群在4个不同群落类型中的生存过程存在较大差异,不同强度的采伐干扰对红松种群生存过程的影响达到显著水平(P<0.05)。
| 来源 Source | 离差平方和 Sum of squares | 自由度 df | 均方 Mean square | F | P |
| * 表示P<0.001 | |||||
| 4个群落Communities | 18066.606 | 3 | 6022.202 | 3.887 | 0.017 |
| 发育阶段Development stage | 3140846.308 | 16 | 196302.894 | 126.688 | 0.000* |
| 残差Error | 54232.477 | 35 | 1549.499 | — | — |
| 合计Total | 6145266.000 | 55 | — | — | — |
红松种群不同年龄阶段存在波动,这种波动的过程和速度是不连续或不均匀的,表现出一定的阶段性和随时间不断发生变化的特征[35]。红松幼苗时期耐阴性较强,适宜郁闭度为0.6—0.7,幼树的适宜郁闭度为0.3—0.5,随着年龄的增加需光量明显增大[18, 21, 29, 36, 37]。本研究中,不同群落内红松幼苗时期死亡率较高,幼树时期存在生长停滞阶段,高龄级期稳定生长,接近生理寿命时的密集死亡,红松种群的天然更新过程呈周期波浪式发展,采伐干扰对红松种群的波动周期无明显影响。
3.2 不同采伐方式和采伐强度对红松种群结构的影响红松种群的恢复依靠种子传播,红松种子来源常受鸟类、鼠类搬运或残留在地被物下的种子数量为转移[39],种子的更新没有一定规律性。皆伐迹地最先更新的是枫桦(Betula costata)、白桦(Betula platyphylla)等小粒种子,以后是五角槭(Acer mono Maxim)、千金榆(Carpinus cordata)、水曲柳、黄檗(Phellodendron amurense)等阔叶树的中粒种子,而针叶树种不能迅速恢复[40]。本研究中,皆伐迹地恢复30余年,红松种群最高仅生长至第5发育阶段,种群结构恢复缓慢,这与红松种子的动物传播方式和皆伐迹地针叶树种恢复周期有关。
齐麟等[23]研究发现采伐会改变群落中物种的优势地位,红松的重要值排名随采伐强度的增加而降低,采伐30a后,红松幼树(胸径<10 cm)占种群数量的比例随采伐强度的增加而增加。本研究中,与原始林相比,40%择伐林和皆伐迹地红松幼树的数量随采伐强度的增加而明显增加,与前人的研究结论相符。而15%择伐林内红松幼树的比例略有减少,这是由于15%择伐林内,高径级产种母树较多,每到丰收季节大量松塔被非法采摘[41],导致林内松籽数量减少,林下幼苗幼树随之减少。
择伐和皆伐改变了红松种群的径级比例,对种群存活曲线和生活过程也有显著影响。有研究表明不同培育措施(抚育、渐伐和封育措施)影响油松的存活曲线,抚育和渐伐后油松种群属进展型,而经过封育措施的油松种群属稳定型[42]。也有研究表明,不同的干扰强度对栓皮栎、翅果油树也有显著影响,在持续受到砍柴、放牧等强烈干扰的群落中,栓皮栎种群呈现衰退势态,而受人为干扰少、发育较好的群落中,栓皮栎种群呈现持续发展势态[43];而对于翅果油树种群,人为干扰限制了翅果油树的稳定可持续生长[44]。由此可见,不同的干扰方式和干扰强度都会对植物种群的结构产生影响,对目标树种干扰强度增大会导致种群稳定性下降甚至衰退,轻度择伐、抚育和渐伐等可持续性经营管理措施能够维持林木的良好生长。未来林场的经营管理应以轻度择伐为主,同时,对于高径级产种母树较多的林分,应当适当予以补植幼苗、幼树以维持其在种群中的比例,使其能够可持续发展。
| [1] | 安玉泽, 田洪华, 孙亚萍. 不同采伐强度对森林土壤温度影响的探讨. 森林工程, 2006, 22(2): 1-2. |
| [2] | Berry N J, Phillips O L, Ong R C, Hamer K C. Impacts of selective logging on tree diversity across a rainforest landscape: the importance of spatial scale. Landscape Ecology, 2008, 23(8): 915-929. |
| [3] | Behjou F K, Mollabashi O G. Selective logging and damage to unharvested trees in a hyrcanian forest of Iran. Bioresources, 2012, 7(4): 4867-4874. |
| [4] | Imai N, Seino T, Aiba S, Takyu M, Titin J, Kitayama K. Effects of selective logging on tree species diversity and composition of Bornean tropical rain forests at different spatial scales. Plant Ecology, 2012, 213(9): 1413-1424. |
| [5] | 郭泉水, 王德艺, 冯天杰, 李东义, 蔡万坡. 雾灵山落叶阔叶林采伐迹地物种多样性和植物种群动态变化研究. 应用生态学报, 1999, 10(6): 645-649. |
| [6] | Lie M H, Josefsson T, Storaunet K O, Ohlson M. A refined view on the "Green lie": Forest structure and composition succeeding early twentieth century selective logging in SE Norway. Scandinavian Journal of Forest Research, 2012, 27(3): 270-284. |
| [7] | Crawly M J. Plant Ecology. London: Blackwell Scientific Publications, 1986. |
| [8] | 李奎, 郑宝强, 王雁, 卜文圣. 滇牡丹自然种群数量动态. 植物生态学报, 2012, 36(6): 522-529. |
| [9] | Khan D, Shaukat S S. Population structure, intraspecific competition and phasic development of Urochondra setulosa (Trin.)//Hubb C E, ed. A Coastal Halophytic Grass of Pakistan. Pakistan Journal of Botany, 1997, 29(2): 271-288. |
| [10] | Svensson J S, Jeglum J K. Structure and dynamics of an undisturbed old-growth Norway spruce forest on the rising Bothnian coastline. Forest Ecology and Management, 2001, 151(1/3): 67-79. |
| [11] | Fuchsa M A, Krannitzb P G, Harestad A S. Factors affecting emergence and first-year survival of seedlings of Garry oaks (Quercus garryana) in British Columbia, Canada. Forest Ecology and Management, 2000, 137(1/3): 209-219. |
| [12] | Molles M C. Ecology: Concept and Applications. 2nd ed. New York: McGraw-Hill Companies, 2002. |
| [13] | Wu J X, Zhang X M, Deng C Z, Liu G J. Structure and dynamic of Populus euphratica population along Tarim River. Pakistan Journal of Botany, 2012, 44(5): 1651-1656. |
| [14] | Yu D P, Zhou L, Zhou W M, Ding H, Wang Q W, Wang Y, Wu X Q, Dai L M. Forest management in northeast China: History, problems, and challenges. Environmental Management, 2011, 48(6): 1122-1135. |
| [15] | 傅立国. 中国植物红皮书——稀有濒危植物: 第一册. 北京: 科学出版社, 1991: 1-736. |
| [16] | 李俊清, 王业蘧. 天然林内红松种群数量变化的波动性. 生态学杂志, 1986, 5(5): 1-5, 12-12. |
| [17] | 伍业钢, 韩进轩. 阔叶红松林红松种群动态的谱分析. 生态学杂志, 1988, 7(1): 19-23. |
| [18] | 乌弘奇. 长白山白桦林中红松种群动态的研究. 生态学杂志, 1989, 8(5): 16-18. |
| [19] | 葛剑平, 郭海燕, 陈动. 小兴安岭天然红松林种群结构的研究. 东北林业大学学报, 1990, 18(6): 26-32. |
| [20] | 阳含熙, 顾连宏. 云冷杉阔叶红松林演替与天然更新过程及红松种群动态数学模型的研究. 自然资源, 1991, (6): 82-82. |
| [21] | Li J Q, Zhu N. Structure and process of korean pine population in the natural forest. Forest Ecology and Management, 1991, 43(1/2): 125-135. |
| [22] | 王惠, 邵国凡, 代力民, 许东, 王飞. 采伐干扰下长白山阔叶红松林主要灌木种群生态位动态特征. 东北林业大学学报, 2007, 35(11): 27-28, 31-31. |
| [23] | 齐麟, 代力民, 于大炮, 周莉, 钟磊, 苏东凯. 采伐对长白山阔叶红松林乔木群落的影响. 湖南农业科学, 2009, (8): 129-132 |
| [24] | 倪瑞强. 长白山典型针阔混交林群落结构与动态研究[D]. 北京: 北京林业大学, 2014. |
| [25] | 李文华, 邓坤枚, 李摇飞. 长白山主要生态系统生物量和初级生产力研究. 森林生态系统研究, 1981, 2: 34-50. |
| [26] | 许广山, 丁桂芳, 张玉华, 程伯容. 长白山北坡主要森林系统土壤有机质及其特征的研究. 森林生态系统研究, 1980, 1: 215-220. |
| [27] | 程伯容, 许广山, 程伯容, 许广山, 丁桂芳, 张玉华, 王伟. 长白山主要森林生态系统的凋落物量及养分含量. 森林生态系统研究, 1984, 4: 19-24. |
| [28] | 苏东凯, 谢小魁, 刘正纲, 于大炮, 周莉, 代力民. 长白山次生白桦林数量特征. 东北林业大学学报, 2009, 37(10): 1-4. |
| [29] | 代力民, 邵国凡. 森林经营决策: 理论与实践. 沈阳: 辽宁科学技术出版社, 2004. |
| [30] | 苏东凯. 长白山阔叶红松林植被恢复研究[D]. 北京: 中国科学院大学, 2010. |
| [31] | 江 洪. 云杉种群生态学. 北京: 中国林业出版社, 1992: 8-40. |
| [32] | 冯士雍. 生存分析Ⅰ. 数学的实践与认识, 1982, (3): 72-80. |
| [33] | 冯士雍. 生存分析Ⅱ. 数学的实践与认识, 1982, (4): 64-74. |
| [34] | 冯士雍. 生存分析Ⅲ. 数学的实践与认识, 1983, (1): 70-76. |
| [35] | 李俊清, 祝宁. 红松的种群结构与动态过程. 生态学杂志, 1990, 9(4): 6-10. |
| [36] | 周以良, 李景文. 中国东北东部山地主要植被类型的特征及其分布规律. 植物生态学与地植物学丛刊, 1964, 2(2): 190-206. |
| [37] | 刘兴双. 长白山次生林群落动态及其生态恢复[D]. 沈阳: 中国科学院沈阳应用生态研究所, 2001. |
| [38] | 孙一荣, 朱教君, 于立忠, 闫巧玲, 王凯. 不同光环境对红松幼苗光合生理特征的影响. 生态学杂志, 2009, 28(5): 850-857. |
| [39] | 姬兰柱, 刘足根, 郝占庆, 王庆礼, 王淼. 松果采摘对长白山阔叶红松林生态系统健康的影响. 生态学杂志, 2002, 21(3): 39-42. |
| [40] | 李景文, 刘庆良, 陈运大. 紅松林皆伐跡地天然更新的研究. 东北林学院学报, 1964, (3): 15-32. |
| [41] | 刘足根, 姬兰柱, 郝占庆, 朱教君, 康宏樟. 松果采摘对长白山自然保护区红松天然更新的影响. 应用生态学报, 2004, 15(6): 958-962. |
| [42] | 吴涛, 张文辉, 陆元昌, 范少辉. 黄龙山林区不同培育措施对油松种群数量动态及物种多样性的影响. 西北植物学报, 2006, 26(5): 1007-1013. |
| [43] | 张文辉, 卢彦昌, 周建云, 张晓辉, 史小华. 巴山北坡不同干扰条件下栓皮栎种群结构与动态. 林业科学, 2008, 44(7): 11-16. |
| [44] | 卢鹏, 徐燕, 毕润成. 不同干扰程度对翅果油树种群结构与动态的影响. 山西师范大学学报: 自然科学版, 2011, 25(3): 103-108. |