 2014, Vol. 34
2014, Vol. 34文章信息
- 陈云峰, 唐政, 李慧, 韩雪梅, 李钰飞, 胡诚
- CHEN Yunfeng, TANG Zheng, LI Hui, HAN Xuemei, LI Yufei, HU Cheng
- 基于土壤食物网的生态系统复杂性-稳定性关系研究进展
- Research progress on ecosystem complexity-stability relationships based on soil food web
- 生态学报, 2014, 34(9): 2173-2186
- Acta Ecologica Sinica, 2014, 34(9): 2173-2186
- http://dx.doi.org/10.5846/stxb201311012644
-
文章历史
- 收稿日期:2013-10-31
- 修订日期:2014-2-13






2. 贺州学院化学与生物工程学院, 贺州 542899;
3. 河南农业大学资源与环境学院, 郑州 450002;
4. 南阳师范学院生命科学与技术学院, 南阳 473061;
5. 中国农业大学资源与环境学院, 北京 100193
2. College of Chemistry and Bioengineering, Hezhou University, Hezhou 542899, China;
3. College of Resources and Environmental Sciences, Henan Agricultural University, Zhengzhou 450002, China;
4. School of Life Science and Technology, Nanyang Normal University, Nanyang 473061, China;
5. College of Resources and Environmental Science, China Agricultural University, Beijing 100193, China
复杂性-稳定性关系是生态学研究热点之一,也是最具争议的观点之一[1, 2]。20世纪50年代,以MacArthur、Elton为代表的生态学家认为复杂性增进稳定性;70年代初,以May为代表的理论生态学家挑战了这一观点,从数学上证明复杂性导致不稳定性[3]。随后几十年,实验生态学家通过分析自然界的真实食物网,理论生态学家通过人为构建食物网模型对复杂性-稳定性关系进行研究,取得了大量的成果[1, 2, 4]。在这个过程中,土壤食物网发挥了重大作用。与水生食物网和地上食物网相比,土壤食物网更易获得、更易解剖,从而更易反映整个生态系统的信息[5],且到20世纪80—90年代,国际上土壤生物学的发展已经为土壤食物网数据整合奠定了基础。因此,以Moore、de Ruiter、Neutel为代表的生态学家,采用理论食物网的数学方法,结合土壤食物网提供的数据,分析了相互作用强度格局(Interaction strength pattern,所有相互作用强度的大小和排列方式)和分室(Compartment,某些物种的集合,集合内部相互作用强,集合之间相互作用无或极弱)对稳定性的影响,生产力和稳定性对食物链长度的影响,以及静态食物网与动态食物网等问题,极大地推进了复杂性-稳定性理论的发展[6]。由于国内土壤食物网研究力量较弱[7],将土壤食物网与稳定性理论串联起来的研究较少[8, 9, 10],且国内在复杂性-稳定性关系方面的综述一般直接列出相关结论[11, 12, 13, 14, 15],而对方法鲜有描述。因此,本文总结了Moore、de Ruiter、Neutel等国外学者以土壤食物网为工具研究复杂性-稳定性关系的基本方法、结论,并探讨了利用土壤食物网研究复杂性-稳定性关系的发展方向,以资国内同行借鉴。
研究复杂性-稳定性关系的难点之一在于难以找到一个可以精确描述复杂性的真实食物网,而土壤生态学的发展恰好为此做好了准备,这些具体工作包括:(1)20世纪80—90年代,欧美建立起了一批可以精确描述食性关系、功能群生物量的土壤食物网,这批土壤食物网数据为土壤食物网模型、理论提供了坚实的物质基础(表 1);(2)与此同时,DeAnglis等[16]在基于初级生产者的Lotka-Volterra(L-V)模型上,建立起了基于碎屑的L-V模型,为土壤食物网L-V模型打下了基础;(3)1987年,Hunt 等[17]建立起土壤食物网面向过程模型,用来描述土壤食物网的能流,为功能群相互作用强度的计算奠定基础。Moore,de Ruiter,Neutel将这3个有关真实食物网的研究数据与May[3, 18]建立起的局域稳定性(Local stability)数学分析方法结合起来,即可判断复杂性-稳定性关系的一些基本问题。具体方法步骤为:(1)利用土壤食物网面向过程模型描述能流,与土壤食物网L-V模型联合计算相互作用强度,进而结合连通食物网描述复杂性;(2)构建群落矩阵(Community matrix)。利用土壤食物网L-V模型描述功能群动态,在平衡点处,采用泰勒展开式将二维L-V模型降为一维,构建雅各比矩阵(Jacobian matrix,A),即群落矩阵;(3)分析群落矩阵特征,探索复杂性-稳定性关系(图 1)。
|
土壤食物网
Soil food webs (SFWs) | 对复杂性-稳定性关系的贡献
Contribute to complexity-stability debate |
|
美国矮草草原土壤食物网
The SFW of North American Shortgrass Steppe,USA | 建立土壤食物网面向过程模型[17] |
|
美国矮草草原土壤食物网
The SFW of North American Shortgrass Steppe,USA 荷兰常规农业与集成农业土壤食物网 The SFW under integrated and conventional agriculture,Netherlands 美国常耕和免耕土壤食物网 The SFW under no-tillage and conventional tillage,USA 瑞典不施氮肥和施氮肥土壤食物网 The SFW with and without nitrogen fertilizer,Sweden. | 从概念和数学证明分室的存在[19, 20, 21],并证明分室可以提高群落稳定性[22, 23]
相互作用强度计算、格局、环、格局与稳定性关系[24, 25] 能量与稳定性共同影响食物链长度[26] |
|
荷兰两个沙丘4个演替阶段的8个土壤食物网
Eight SFWs derived from four successive stages at two dunes,Netherlands | 静态食物网与动态食物网关系[6, 27] |

|
| 图1 利用土壤食物网分析复杂性-稳定性关系示意图 Fig. 1 Schematic illustration of analysis of complexity-stability relationships based on soil food web |
根据食物网基位种的性质,L-V模型可以分为 基于初级生产者(有生命特征)和基于碎屑(无生命 特征)两类[6]。土壤食物网属于基于碎屑的食物网,碎屑的增长为捐赠控制模式[22],其增长依靠外部枯枝落叶投入、土壤生物的自然死亡或未被捕食后的死亡,而不是像初级生产者一样能自我生长和繁殖(图 2)。由于传统食物网理论主要建立在基于初级生产者的模型上,因此,本文对这两种模型均加以描述(图 2)。生产者和碎屑动态可以用方程(1)和(2)描述,但这两类模型的消费者可用同一方程描述,见方程(3)[6, 26, 28]。

生产者动态:
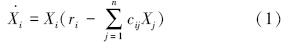
式中,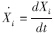 物种i(生产者)增长率(g C · m-2 · a-1);Xi为物种i种群密度 (g C · m-2);
Xj为物种j(捕食者)种群密度(g C · m-2);
ri为物种i特定生长率或称内禀生长率,即出生率减去死亡率 (a-1);
cij为物种j(捕食者)对物种i(生产者)消费系数,单位为(g C · m-2)-1 · a-1;
riXi为物种i(生产者)自然增长量(g C · m-2 · a-1);
物种i(生产者)增长率(g C · m-2 · a-1);Xi为物种i种群密度 (g C · m-2);
Xj为物种j(捕食者)种群密度(g C · m-2);
ri为物种i特定生长率或称内禀生长率,即出生率减去死亡率 (a-1);
cij为物种j(捕食者)对物种i(生产者)消费系数,单位为(g C · m-2)-1 · a-1;
riXi为物种i(生产者)自然增长量(g C · m-2 · a-1);
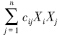 为物种i(生产者)被物种j(消费者)取食的生物量(g C · m-2 · a-1)。若i = j,表明初级生产者i种群密度下降由种内竞争引起。
为物种i(生产者)被物种j(消费者)取食的生物量(g C · m-2 · a-1)。若i = j,表明初级生产者i种群密度下降由种内竞争引起。
碎屑动态:
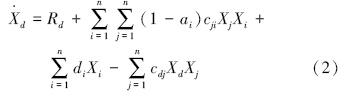
式中,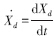 碎屑增长率(g C · m-2 · a-1);
Rd为落叶和根系分泌物进入土壤的速率(g C · m-2 · a-1);
ai为功能群i的同化效率(无量纲);
cji为功能群i对功能群j的消费系数 (g C · m-2)-1 · a-1;
Xi、Xj为功能群i、功能群j的种群密度(g C/m2);
di为功能群i特定死亡率(非捕食死亡率,包括内禀死亡率和种内竞争死亡率)(a-1);
cdj为功能群j对碎屑的消费系数(g C · m-2 · a-1);
碎屑增长率(g C · m-2 · a-1);
Rd为落叶和根系分泌物进入土壤的速率(g C · m-2 · a-1);
ai为功能群i的同化效率(无量纲);
cji为功能群i对功能群j的消费系数 (g C · m-2)-1 · a-1;
Xi、Xj为功能群i、功能群j的种群密度(g C/m2);
di为功能群i特定死亡率(非捕食死亡率,包括内禀死亡率和种内竞争死亡率)(a-1);
cdj为功能群j对碎屑的消费系数(g C · m-2 · a-1);
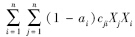 为各功能群未被同化的部分(g C · m-2 · a-1);
为各功能群未被同化的部分(g C · m-2 · a-1);
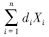 :各功能群自然死亡(非捕食)的有机残体(g C · m-2 · a-1);
:各功能群自然死亡(非捕食)的有机残体(g C · m-2 · a-1);
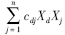 为被食碎屑者取食的生物量(g C · m-2 · a-1)。
为被食碎屑者取食的生物量(g C · m-2 · a-1)。
消费者动态:
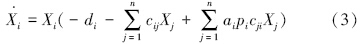
式中,Xi、Xj为功能群i、功能群j的种群密度(g C/m2);
di为物种i特定死亡率(非捕食死亡率,包括内禀死亡率和种内竞争死亡率)(a-1);
cij为物种j(此处为捕食者)对物种i(被捕食者)的取食系数(g C · m-2 · a-1);
ai为物种i的同化效率(无量纲);
pi为物种i生产效率(无量纲);
cji为物种i(此处为捕食者)对物种j(被食者)的取食系数(g C · m-2 · a-1);
diXi为物种i特定死亡量(g C · m-2 · a-1);
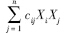 为物种i(此处为被食者)被物种j(捕食者)捕食的量(g C · m-2 · a-1;
为物种i(此处为被食者)被物种j(捕食者)捕食的量(g C · m-2 · a-1;
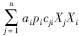 为物种i(此处为捕食者)取食物种j(被食者)后的增长量(g C · m-2 · a-1);
方程(1)—(3)中各系数均取正值。方程(1)和(3)可以用通用L-V模型统一起来,即 :
为物种i(此处为捕食者)取食物种j(被食者)后的增长量(g C · m-2 · a-1);
方程(1)—(3)中各系数均取正值。方程(1)和(3)可以用通用L-V模型统一起来,即 :
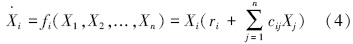
这里ri可正可负,分别代表内禀增长率也可以代表特定死亡率。cij也可正可负,可以代表捕食者对被食者的影响、被食者对捕食者的影响及种内相互作用。
1.1.2 土壤食物网面向过程模型土壤食物网面向过程模型定量描述了捕食者取食被食者后生物量的流转过程(图 3)。捕食者j取食被食者i (取食率,Fj)后,以一定比率(同化效率,aj)将此生物量同化(ajFj),同化的生物量又以一定比率(生产效率,pj)转化为捕食者的生物量(ajpjFj),用于营养生长和生殖生长。在这个转化过程中,能量利用效率(ej)等于ajpj。未同化的部分((1-aj)Fj)进入碎屑中,见方程(2);同化后,未用于生长的部分以矿化的形式(以碳化碳cminij或矿化氮Nij来表示)进入环境,即cminij=(1-pj)ajFj 或Nij=aj(1/CNi-pj/CNj)Fij,式中CNi、CNj分别为被食者和捕食者的碳氮比。

在上述能量转化过程中,最终要计算的为取食率。在面向过程模型中,假定各功能群处在平衡态,即各功能群生物量保持恒定。此时,功能群增长的生物量等于死亡量,其中死亡量包括自然死亡部分(djBj,Bj为功能群j生物量)和被其他功能群捕食的部分(Mj),即Fjajpj=Mj+djBj,式中,dj、aj、pj为常量[17, 29],Bj为田间实测数据。Mj计算时需从顶级功能群开始,因为顶级功能群只取食其他生物而不被其他生物捕食,此时Mj=0。次级功能群Mj则为顶级功能群的Fj。若考虑杂食性,食率计算公式修改为: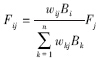 ,式中Fj代表捕食者j对其所有猎物的取食率,Fij代表捕食者j对其某一猎物i的取食率,wij为捕食者j对猎物i的相对取食偏好,Bk为猎物k的生物量。
,式中Fj代表捕食者j对其所有猎物的取食率,Fij代表捕食者j对其某一猎物i的取食率,wij为捕食者j对猎物i的相对取食偏好,Bk为猎物k的生物量。
比较土壤食物网L-V模型和面向过程模型可以看出,L-V模型描述了群落中每个功能群密度变化,面向过程模型则描述了某个功能群的能量收支状况。两者虽然侧重点不一样,但又有一定的关系。以方程(3)为例,功能群i被功能群j取食的量,在L-V模型中以cijXiXj表示,在能量模型中以Fij表示,显然Fij=cijXiXj,此外功能群i(此处为捕食者)取食物种j(被食者)后的增长量aipicjiXjXi=aipiFij。由于功能群相互作用系数不能直接测出,而Fij能从面向过程模型中得出,Xi、Xj能田间测出,因此两模型的连接,可以计算cij或cji[27]。
1.2 生态系统复杂性概念和计算 1.2.1 复杂性与多样性复杂性是一个难以精确定义的概念,不能用一个统一的数学公式来描述,许多文献里也称为多样性,但这两者还是有一定差别的[31, 32, 33]。Naeem[31]将复杂性细分成群落复杂性、进化复杂性和功能复杂性。群落复杂性主要描述了群落的结构和各物种间的相互作用;进化复杂性描述了各物种之间进化距离的长短,即基因差异的大小;功能复杂性描述了生态系统的物流和能流情况。在土壤食物网中,复杂性一般指群落复杂性和功能复杂性。群落复杂性用物种个数(n)、连通度(c)、相互作用强度(α)来表示[3],Montoya等[34]进一步将格局、杂食性、功能群、物种之间的距离(即两个物种之间链接长度)等特征也归为复杂性。功能多样性用功能群数目及能流组织形式(生物量分布及各能流途径的能流大小)[30]来表示。多样性用来描述群落的丰富度和均匀度,而不涉及相互作用强度,尽管两者类似且正相关,但多样性高的群落若相互作用强度很低,其复杂性不一定高,反之亦然[31, 32]。因此,多样性仅仅指描述了复杂性的一个方面,即群落复杂性的部分内容[31]。但在应用研究中,多样性比复杂性研究的更多,一方面因为测量多样性容易,一方面因为多样性与当前生物多样性的丧失这个研究热点比较契合[2]。
1.2.2 相互作用强度
食物网模型3个参数(物种数、连通度和作用强度)中,物种数和连通度一般容易测量,相互作用强度不仅测量难,而且在定义上也比较复杂。理论生态学家和实验生态学家所描述的相互作用强度不一样,甚至在理论生态学家内部或实验生态学家内部所用的相互作用强度概念也不一样[35, 36]。Berlow等[36]综述了目前相互作用强度的概念和度量,将其归为两大类,一类为一物种对另一物种动态的影响,一类为去除某一物种后对整个食物网的影响,前者是对个体的影响,后者是对整体的影响。在土壤食物网中,采取的是第一类相互作用强度。功能群j对功能群i的作用强度(αij)为平衡态( )时,功能群j密度轻微变化后,功能群i密度的瞬时变化率[27, 36, 37, 38]。这个定义在数学上可用偏导数描述,即
)时,功能群j密度轻微变化后,功能群i密度的瞬时变化率[27, 36, 37, 38]。这个定义在数学上可用偏导数描述,即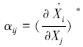 ,式中
,式中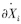 代表功能群i密度的瞬时变化率,即
代表功能群i密度的瞬时变化率,即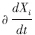 ,*代表平衡态。
,*代表平衡态。
土壤食物网相互作用强度计算步骤为:(1)采用基于碎屑的L-V模型描述功能群动态,构建群落矩阵;(2)根据土壤食物网面向过程模型与L-V模型之间的关系,将田间观测数据代入群落矩阵中,计算出相互作用强度。许多学者从各个侧面描述过具体计算过程[3, 6, 24, 25, 27, 30, 36, 39, 40, 41],为了更有代表性,此处先介绍通用L-V模型相互作用强度的过程,然后将描述土壤食物网的方程代入,计算土壤食物网功能群相互作用强度。
通用L-V模型见方程(4),平衡态时物种i种群密度记作X*i。在平衡点处,方程(4)的泰勒展开式为:
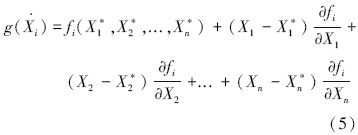
根据平衡态的定义,fi(X*1,X*2,...,X*n)=0,令xi=Xi-X*i,则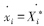 ,方程(5)可以用写成矩阵形式,即:
,方程(5)可以用写成矩阵形式,即:

其中,各偏导数组成的矩阵即为雅各比矩阵,也即群落矩阵,将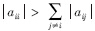 展开,则:
展开,则:

该矩阵以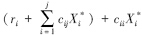 为对角线,将矩阵分为上三角和下三角两部分。上三角元素CijX*i代表了物种j(捕食者)对物种i(被食者)的相互作用强度,即物种i被物种j取食的死亡率,为负值;下三角元素CjiX*j代表了物种i物种j的相互作用强度,即物种j取食物种i后的增长率,为正值。对角线元素代表了种间相互作用强度,为负值。因平衡态时,
为对角线,将矩阵分为上三角和下三角两部分。上三角元素CijX*i代表了物种j(捕食者)对物种i(被食者)的相互作用强度,即物种i被物种j取食的死亡率,为负值;下三角元素CjiX*j代表了物种i物种j的相互作用强度,即物种j取食物种i后的增长率,为正值。对角线元素代表了种间相互作用强度,为负值。因平衡态时,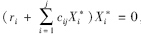 ,所以对角线元素又可以写成ciiX*i。
,所以对角线元素又可以写成ciiX*i。
将方程(2)和(3)带入方程(6)中,土壤食物网的群落矩阵(As)可以写成:
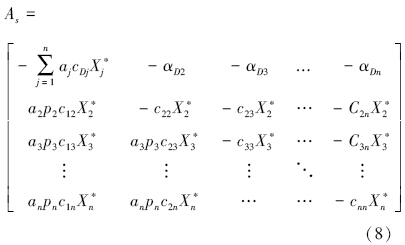
其中,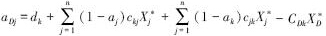 ,式中及矩阵As各系数为正值,表示了食碎屑者对碎屑的作用强度,若CDkX*D=0,则表示为非食碎屑者对碎屑的作用强度。
,式中及矩阵As各系数为正值,表示了食碎屑者对碎屑的作用强度,若CDkX*D=0,则表示为非食碎屑者对碎屑的作用强度。
矩阵As中元素可以分成5类:(1)下三角元素,表示被食者对捕食者的影响,;(2)上三角第一排元素,表示捕食者对碎屑的相互作用强度;(3)上三角除去第一排的元素,表示捕食者对被捕食者(除去碎屑)的相互作用强度;(4)对角线第一个元素,表示碎屑种内相互作用强度;(5)除去第一个元素的其他对角线元素,表示除碎屑外其他功能群的种内相互作用强度。上三角元素(除去第一排)中,X*i及其系数均为未知数。在现实中,功能群的生物量往往多年多次取样,其平均值(Bi)可以近似代替平衡态时的生物量,即X*i=Bi。此外,由于Fij=cijXiXj,aipicjiXjXi=aipiFij,因此,
捕食者j对被捕食者i的相互作用强度为:
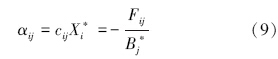
被食者i对被捕食者j的相互作用强度为:
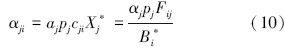
类似的,各功能群对碎屑的作用强度为:

碎屑种内相互作用强度为:
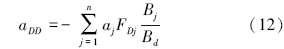
除碎屑外,其他功能群种内相互作用强度,从现有田间试验数据和理论上都难以估算。实际上,在定义特定死亡率dj时(“特定”意为功能群死亡率随其生物量的变化而变化),即包含内禀死亡率和因种内相互作用而造成的死亡率,因此,种内相互作用强度为特定死亡率dj的一部分,即:

与相互作用强度相比,稳定性的定义及度量更复杂[1, 2, 42]。稳定性概念可分为内涵和外延两个部分,内涵包括抵抗力、恢复力、持久性、变异性等,外延包括局域稳定性和全局稳定性等[11]。在利用土壤食物网研究复杂性-稳定性关系中,稳定性采用的是局域稳定性,也叫邻近稳定性[4]、动态稳定性[26],即系统经受小的干扰后回到平衡态的能力,若系统经受大的干扰后依然能返回平衡态,此时的系统具有全局稳定性。
通用L-V模型(方程(4))用可以用矩阵的方式表示(方程(6)),由于矩阵代表了一种线性变化,矩阵特征值的正负代表了线性变换的方向,因此若矩阵A所有的特征值(实数或复数实部)均小于0,则表明L-V模型中所有物种数量都朝着平衡态方向的发展,即系统趋于稳定。可见,分析群落矩阵的特征值即可以判断群落的稳定性。对于一个具体的矩阵,也可以不用计算特征值,只需根据矩阵结构,采用Routh-hurwitz 判别法[6]即可来判断所有特征值是否都小于0或有负实部(虚数实部小于0),进而定性判断群落稳定性。但Routh-hurwitz 判别法是建立在各相互作用强度都能测定出来的基础之上。对于某些相互作用强度不能完全测定的食物网,如土壤食物网中种内相互作用强度群落不能测定,还可以采用McKenzie定理来判断:若群落矩阵中每一行的非对角元的模之和都小于这一行的对角元的模,即 ,则该群落稳定[6],满足这种结构的矩阵称为对角线优势矩阵。由于αii=-sidi(0≤si≤1),因此,在比较多个群落的稳定性,若群落A满足Mckenzie定理所需的si要小于群落B所需要的si,则认为群落A比群落B稳定[25]。
理论上,复杂性-稳定性关系最终转变成对群落矩阵特征值性质的讨论,为了使读者对群落矩阵、复杂性、稳定性之间关系有个清晰的认识,将其对应关系归纳在表 2。
| 复杂性和稳定性Complexity and stability | 群落矩阵Community matrix |
|
物种数或功能群数目
The numbers of species or functional groups | 维度
Dimensionality |
| 联通度connectedness | 非零元素占总元素的比例
Proportion of elements that were non-zero |
| 相互作用强度Interaction strength | |
|
种间相互作用强度大小和类型
Extraspecific interaction strength | 以对角线为对称轴的成对元素的正负符号和绝对值大小
Sign and absolute value of paired off-diagonal elements |
|
种内相互作用强度大小和类型
Interspecific interaction strength | 对角线元素正负符号及绝对值大小
Sign and absolute value of diagonal elements |
| 格局Pattern | 元素的排列形式 Arrangement of elements |
| 计算过程Process of calculation | 求解L-V模型的雅各比矩阵
Solute the Jacobian matrix of L-V mode |
| 局域稳定性Local stability | 所有特征值或虚数实部小于0
All eigenvalues or the real parts of the eigenvalues are negative |
| 稳定性的判断Criterion of stability | Routh-hurwitz 判别法 Routh-hurwitz criteria
McKenzie定理 McKenzie′s theorem 最大环重 Maximum loop weight |
在建立起利用土壤食物网研究复杂性-稳定性关系的基本方法后,Moore、de Ruiter、Neutel等对复杂性-稳定性关系进行了一系列的探索,取得了大量的成果,主要表现在3个方面,即相互作用强度格局、分室与生态系统稳定性之间的关系,及生产力与稳定性共同对食物链长度的调节。
2.1 相互作用强度格局对生态系统稳定性的影响格局为食物网的结构特征之一[43],土壤食物网中的最主要的格局为相互作用强度格局和分室[30]。其中相互作用强度格局呈现出两个特征:(1)成对出现的相互作用强度是不对称的。捕食者j对被食者i的相互作用强度(aij,负值)绝对值高于被食者对捕食者的相互作用强度(aji,正值)1至2个数量级,表现在群落矩阵中,即上三角元素绝对值远高于下三角的对应元素;(2)aij绝对值在低营养位时较高,随着营养位的升高而显著降低;aij则在低营养位时较低,随着营养位的升高而显著升高[24, 40, 44]。
de Ruiter等[24, 40, 44]进一步以随机构建的食物网为参照物,利用群落矩阵探讨了相互作用强度格局对稳定性的影响。这种影响可以从某对相互作用强度和所有相互作用强度的排列两个方面进行描述。由于群落矩阵中以对角线为轴的成对元素,即aij和aji代表了相互作用的两个方面,因此,随机改变某对aij和aji的值,观察群落矩阵的稳定性,即可判断出某对相互作用对稳定性的影响。具体做法为:将群落矩阵中其他元素固定,在[2aij,0]或 [0,2aji]内随机取样100次后,其中稳定矩阵所占的比例,即代表了某对相互作用强度对稳定性的影响。结果表明,相互作用强度大小对稳定性影响无关。所有相互作用强度的排列对稳定性的影响主要通过比较“真实矩阵”和“随机矩阵”来判断。真实矩阵也叫逼真矩阵,矩阵中各元素位置不变,但大小随机变动。随机矩阵中,矩阵的联通度不变、多样性不变、平均相互作用强度不变,但各成对元素与其他成对元素进行随机对换,并确保每对元素都与其他成对元素至少对换一遍,这种对换实际上破坏了相互作用强度的排列方式。因此,该矩阵也称为干扰矩阵。此外,为了与前人[45]研究结果相比较,构建了理论矩阵和检测矩阵。理论矩阵中捕食者对被食者的作用强度在[-10, 0],被食者对捕食者的作用强度在[0,0.1]范围内,检测矩阵中用来计算相互作用强度的参数,如同化效率等不是田间试验结果而来,而是随机取的。各矩阵均采用Monte Carol方法,以随机改变矩阵中元素1000次时,稳定群落矩阵的百分率来代表各矩阵的稳定性。结果表明,逼真矩阵稳定性远高于其他几个矩阵,这表明相互作用强度以一定格局存在提高了系统的稳定性。
Neutel等[24]引进了“环”的概念,进而证明了为什么相互作用强度的这种分布模式能提高系统的稳定性。环一般包括数个强相互作用(捕食者对被食者的相互作用强度)和弱相互作用(被食者对捕食者的相互作用强度),环中物种的个数称为环长,各相互作用强度的几何平均值称为环的权重,简称环重。Neutel等[25]证明了最大环重为满足群落稳定的最小si值的近似,因此,最大环重指示了土壤食物网的稳定性。真实食物网中,相互作用强度分布模式为较长的环中弱相互作用强度较多,这种分布模式决定了最大环重较小,更接近si值,从而使群落更稳定。由于生物量金字塔的形状(即金字塔斜面,从低到高各营养级生物量递减规律)决定了食物网的环长和环重,因此本质上是食物网的能量特征决定了食物网的稳定性[37, 46]。在研究荷兰两个沙丘4个演替阶段的8个土壤食物网,Neutel等[27, 37]进一步证明了环长为3或3以上的环能指示稳定性,且细菌食物链中的金字塔斜面越低(各营养级生物量差异越大),食物网越稳定,它们是决定稳定性的关键环和关键金字塔。
2.2 分室与食物网稳定性关系Moore及其同事采用多元统计的方法分析了美国科罗拉多州中部矮草草原土壤食物网等7个食物网,从数学上证实了分室的存在[20, 21, 47],并从功能群的生理特征、生境、生活史特征等方面描述了分室的生态学意义和边界[19, 48],利用Post等[49]创造的方法分析了分室对生态系统稳定性的影响[22, 23],Rooney等[50]进一步证明除土壤食物网外,分室在许多群落中均存在。土壤食物网分室由能流和营养关系决定,因此又可称为能量通道。根据能量来源途径,可以将土壤食物网分成3个分室,即3个能量通道,分别为细菌通道、真菌通道和根系通道。细菌通道中的土壤生物主要包括细菌、原生动物、食细菌线虫等,它们生活在充满水的孔隙和水膜中,与真菌通道的生物相比,世代更替、物质和能量流动均较快,因此通常又称为“快”通道;真菌通道中的土壤生物主要包括真菌、食真菌线虫和微节肢动物,它们的生境为土壤中充满空气的孔隙和水膜中,世代更替、物质和能量流动均较慢,因此又称为“慢”通道;根系通道主要由根系和植物寄生线虫组成。实际上,分室是某些占据相同生态位的生物的集合,其适应性特征(生境、生活史、生理参数)决定分室的边界。如取食同一真菌的各种微节肢动物,其生境相同,取食时间类似,活动空间也类似,它们之间相互作用的机会较多,因此,真菌、微节肢动物及微节肢动物的捕食者将形成一个分室[47]。而分别取食细菌和真菌的微节肢动物,其取食时间、活动空间不一致,生境也不同,因此,它们不在一个分室。本质上,分室是环境异质性造成的,且在低营养级更容易形成分室[51]。
分室对稳定性的影响即判断细菌通道和真菌通道相对大小对稳定性的影响。所采用的分室模型见图 4,具体步骤为[22, 23]:(1)采用霍林Ⅱ型L-V模型描述分室模型中细菌、食细菌生物、真菌、食真菌生物和捕食者的动态;(2)将碎屑的量(N)固定,变换易利用碎屑占总碎屑量的比例(p),求取群落矩阵;(3)求取群落矩阵特征值,若所有特征值小于0或有负实部,则群落稳定。结果表明,在0.2 < p < 0.6时群落最稳定,这表明分室有助于提高群落的稳定性。
2.3 生产力和稳定性共同影响食物网链长度
自然界中食物链长度到底受什么因子限制,是生态学中一个基本的问题,由此问题可以引出食物网内部的调控机制问题,如营养级联概念的提出[52]。目前的研究认为限制食物链长度的因子有很多,如能量(生产力)、动态稳定性、干扰、生态系统大小、群落组织、捕食相互作用强度大小等[53],但主流观点仍然是初级生产力和动态稳定性共同限制了食物链的长度。Moore 等[6, 26, 28, 41, 52, 54]利用土壤食物网数据、基于初级生产者或碎屑的食物链L-V模型,将Hutchinson[55]能量沿着食物链递减从而限制了食物链长度及Pimm和 Lawton[56]稳定性限制了食物链长度的观点结合起来,证明了能量和动态稳定性两者共同影响了食物链的长度,而不是某一方面单独作用。
Moore等采用图 2中的6条食物链来探索能量、稳定性和食物链长度的关系。基于初级生产者的食物链,生产力为基位种通过生长和繁殖的生物量增长率,即b1X*1,X*1为基位种平衡态时的生物量,通过改变b1的大小可以模拟生产力的变化。基于碎屑的食物链,其生产力为增加的碎屑生物量,可通过改变Rd的大小来模拟生产力的大小。该模型需满足生物学上的现实性,即各物种生物量均大于0,若延伸到某一级食物链上该物种生物量小于或等于0(在数学上可以存在),该食物链断裂,不再延长。这种性质,在数学上用食物链可行性来判断,即在生产力、死亡率、同化效率、增长效率、消费系数等参数范围内随机取样,构建一定数量的一定链长的食物链,其中可行性食物链(所有物种大于0)的比例即为可行性。以链长为3的食物链为例,在某一特定的生产力条件下,所有参数(生产力,死亡率、同化效率、增长效率、消费系数)在(0,1)内均匀分布,随机取样10000次,计算出的3个物种均大于0的次数占10000的比例,即为链长为3的食物链的可行性。可行性食物链的稳定性采用恢复力来(Resilience)表示,恢复力用返回时间(Return time,RT)来度量,RT=-1/Real (λmax),λmax为最大特征值,Real代表实部,即群落矩阵最大特征值负实部的倒数。返回时间越小,表明食物链能更快的返回到平衡态,稳定性更高。
在构建完食物链模型、建立起度量生产力对食物链长度和稳定性的影响公式后,可以通过改变生产力、方程(1)—(3)参数(死亡率、同化效率、增长效率、消费系数)的大小和分布来模拟能量和稳定性是如何限制食物链长度。Moore等从3个方面证明了生产力和稳定性是如何限制食物链长度的:
(1)从纯理论的角度证明生产力和稳定性共同影响食物链长度
纯理论上,所有参数(生产力,死亡率、同化效率、增长效率、消费系数)在(0,1)内均匀分布。模拟结果表明,食物链可行性随着食物链的延长而降低,但在同等食物链长度下,基于碎屑的食物链可行性较高。食物链的返回时间随着食物链的延长而增加,这表明较长的食物链稳定性较差。
(2)模拟自然界中生产力,证明生产力和稳定性共同影响食物链长度
根据土壤食物网中死亡率、同化效率、增长效率、消费系数的范围和自然界中各类型生态系统的生产力,可将生产力调至10-2—105 7个数量级,其中1 = 36或4 g C m-2 a-1。计算可行性发现,D2食物链(图 2)在整个生产力梯度内可行性为100%,其余5条食物链可行性随着生产力提高而提高,在某一生产力水平下,随着食物网链长度的增高可行性下降。计算返回时间发现,随着生产力提高食物链稳定性提高,在低生产力水平下(≤ 10),较长的食物链稳定性低;在高生产力水平下(> 10),链长为4的食物链比链长为3的食物链稳定,但又不如链长为2的食物链稳定。
(3)从能量效率角度证明生产力和稳定性共同影响食物链长度
自然界中各生物的能量效率差异很大,如变温动物和恒温动物。可行性和返回时间均对能量效率很敏感,但在上述分析中,能量效率在(0,1)均匀分布,这与现实世界差异较大。通过将能量效率分布调整至Beta分布,可以模拟生产力和稳定性对具有不同能量效率的食物链长度的影响。以Beta(9,2)代表高能量效率生物,Beta(2,9)代表低能量效率生物,生产力范围10-2—105,死亡率、消费系数(0,1)内均匀分布,分别计算可行性和返回时间,结果表明:在整个生产力范围内,两种能量效率下的D2食物链可行性为100%,其余5条食物链中,在低生产力范围内(≤ 10),高能量效率的食物链可行性更高,当生产力大于10时,高能量效率的食物链可行性均达到100%;链长为3、4时,高能量效率的食物链稳定性更高,链长为2时,在低生产力范围内,高能量效率的食物链稳定性更高。
以上利用简单食物链模型模拟的结果可以进一步推广到复杂的模型中[6]。这些结果说明生产力必须达到一定程度才能维持食物链的延长,同时,食物链的长度又必须确保食物链能抵抗外界的干扰,从而维持一种动态的平衡。
3 静态食物网和动态土壤食物网尽管Moore、de Ruiter、Neutel等以土壤食物网为工具研究复杂性-稳定性关系取得了很多的成果,但连他们自己也承认其研究存在不足之处,即采用静态食物网的分析方法对待动态土壤食物网[6, 28, 37]。面向过程模型中,输入数据为各功能群年际平均生物量,由此而决定了利用土壤食物网研究复杂性-稳定性关系时,系统时间尺度为1a,年内的各功能群时空动态被忽略[57]。在这个时间尺度内,土壤食物网的连通性(功能群个数、相互作用强度类型)固定、形状(食物网链长度、分室等)固定、参数(特定死亡率、同化效率、生长效率等)固定[6]。除忽略了土壤食物网的动态外,静态食物网分析方法本身也存在一些缺陷,如:(1)平衡态假设中,利用各功能群多年生物量平均值作为平衡态时功能群的生物量,但到底多少次采样的平均值才能代表平衡态,并没有一个准确的数字[27, 58, 59]。从逻辑上讲当然采用次数越多,越接近平均值,但底线是多少,并没有一个标准,而这在指导实验生态学家上有重要意义,即如何利用最少的工作量取得最多的信息;(2)土壤食物网中的营养关系很多也是模糊的,与同位素方法研究的营养关系差距较大[60];(3)L-V模型中,以霍林Ⅰ型功能来反应功能群间的相互作用过于简单,在现实世界中,霍林Ⅰ型功能很少存在[28, 37];(4)忽略了功能群内的各生物取食偏好、对环境压力的响应不同[57];(5)在面向过程模型中,仅考虑捕食作用,忽略了一些其他作用,如种内竞争等(L-V模型包含此种相互作用)、互利等[56]。可见,利用土壤食物网研究复杂性-稳定性关系中,忽略了很多特征,但这也为新一代土壤食物网模型的发展留下了空间。
尽管忽略食物网动态在研究复杂性-稳定性关系中存在着不足,但Moore、de Ruiter、Neutel等也证实了土壤食物网中,静态分析可以作为动态分析的一个近似。这是因为:(1)平衡态假设是可行的。首先,土壤食物网中功能群的年际动态是相似的,其差异远低于年内之间的变化率[59];其次,以平衡态假设为基础的面向过程模型,其模拟的碳氮矿化与实测结果相似[61];再次,逼真矩阵和随机矩阵(干扰矩阵、理论矩阵、检测矩阵)的相互作用强度、稳定性的计算都基于平衡态假设,但逼真矩阵稳定性远高于随机矩阵,这证明稳定性取决于现实观测的数据而不是取决于平衡态假设[6, 24];(2)忽略时空动态是可行的。在最初提出取食率公式Fj= djBj+Mj+ΔBj ajpj 时,已经考虑到了动态,因此增加了ΔBj一项,但Neutel 等[27]在研究土壤食物网的演替过程中发现ΔBj与(djBj+Mj)相比量很低,可以忽略。
下一代土壤食物网模型的发展方向无疑是动态土壤食物网[6, 57]。目前,动态食物网的发展主要停留在理论上,相关的模型、概念和理论较多(表 3),甚至将其扩展到生态网络、社会网络和信息网络等[62]。但在现实世界中,仍然难找到一个描述的很好的动态食物网,土壤食物网也不例外。Dekker 等[57]描述了动态土壤食物网应该包括的3个内容:间接作用(如对枯枝落叶的粉碎、搬运、混合等)、时间动态和空间动态,但其在指明动态土壤食物网这个发展方向的同时,并没有明确指出如何构建动态土壤食物网。
| 静态食物网
Static food web | 动态食物网
Dynamic food web | |
|
平衡态假设
Equilibrium assumption | 有 | 无 |
|
个体特征
Individual properties | 个体大小、生长和行为不变[6] | 个体大小、生长和行为发生变化,如在各个生命阶段的生理机能(出生率、死亡率、扩散率、捕食能力等)差别比较显著[6, 63] |
|
种群特征
Population properties | 种群数量在平衡点上下波动,生活史策略不变[6, 51] | 数量、生活史策略参数发生变化[51] |
|
群落特征
Community properties | 群落物种无迁进、迁出、时空变化等,群落组成、相互作用类型和强度不变。体现在雅各比矩阵上即矩阵维度、各元素符号、大小不变[4, 6] | 群落存在迁进、迁出、时空变化等,群落组成、相互作用类型和强度发生变化。体现在雅各比矩阵上即矩阵维度、各元素符号、大小均可以变。相互作用强度非线性,即霍林Ⅱ或Ⅲ型功能反应[4, 6] |
|
群落结构比喻
Metaphor of community structure | 石头拱桥。石头代表物种,石头的粘结代表相互作用强度,关键种在稳定拱桥结构中起着最关键作用的石头[6] | 抽积木游戏[6, 51] |
|
拓补结构
Topology structure | 采用静态模型描述基位种、中间种及顶位种的比例、连通度、链接密度等[64] | 采用随机模型描述食物网动态结构,包括级联模型(cascade model)、生态位模型(Niche model)、嵌套等级模型(nested-hierarchy model)[65] |
|
食物网动态模拟
Modeling Dynamic of food web | 霍林Ⅰ型功能反应L-V模型
| 霍林Ⅱ或Ⅲ型功能反应L-V模型;
生物能量动态模型(Bioenergetic dynamics model);异速生物能量动态模型(Allmetric bioenergetic dynamics)[66] |
|
相互作用类型
Interaction types | 捕食关系[57] | 除捕食外,还包括互利、寄生等类型[67],或这些相互作用混合在一起[68] |
| 稳定性Stability | 局域稳定性 | 持久性(Permanence)[69] |
| 时间尺度Time scales | 渐进行为(Asymptotic behavior)[6, 68] | 瞬时动态(Transient dynamics)[6, 70] |
复杂性-稳定性关系是生态学中永恒的主题之一,随着人们对群落、生态系统认识的不断深入而深入。Moore、de Ruiter、Neutel等人以土壤食物网为模式食物网,对复杂性-稳定性关系中的一些问题进行了探讨,大大推进了复杂性-稳定性关系研究。此外,以土壤食物网研究复杂性-稳定性的不足之处,实际上也为下一代土壤食物网模型指明了研究方向,驱动了利用土壤食物网研究复杂性-稳定性关系的理论发展。
展望未来,从土壤食物网稳定性理论和应用两个方向加以概括,具体包括:
(1)结合长期定位试验,加强土壤生物基础研究,为构建动态土壤食物网提供数据支持
从利用土壤食物网研究复杂性-稳定性的发展史来看,土壤生物生理、生态以及分类的发展奠定了土壤食物网模型的基础,为理论分析提供了素材。土壤食物网面向过程模型中的输入为真实的观测数据,但L-V模型中并没有融入观测数据,仅仅只是借用模型来推导相互作用强度及建立稳定性判别方法。以目前的观测数据,尚不能建立起土壤食物网L-V模型。但随着土壤生物检测技术的提高和参数(死亡率、同化效率、生长效率)的进一步精确,利用观测数据建立起土壤食物网L-V模型是可能的,即使不能实时观测整个食物网数据的话,也可以观测部分土壤食物网如微食物网等[71]。目前,世界和中国已建有大量有影响力的长期土壤观测试验站[72],如果能利用这些试验站长期观测各功能群的动态,从而明确土壤食物网的时空动态,将能抛弃平衡态假设,建立起真正的动态土壤食物网。
(2)利用食物网稳定性理论指导农田土壤食物网管理
稳定性之所以引起生态学家的重视,正是因为自然界中各类干扰太多,进而对人类生存环境造成伤害。对于农田土壤食物网来说,各种农业管理措施(如耕作、施肥、灌溉、病虫害防治)和气象灾害均是一种干扰,这些干扰通过改变食物网能量组织形式来改变土壤食物网稳定性[30]。此外,这种干扰可以看成是一种较小的干扰,因为无论是农业管理措施还是极端气候都很少使农田土壤食物网崩溃,因此,农田土壤食物网稳定性可以看成是一种局域稳定性。维护农田土壤食物网的稳定性,即可维持土壤生态系统的功能。目前农业管理措施对农田土壤食物网稳定性的影响研究较少,de Ruiter等[40]通过比较土壤食物网的逼真矩阵,发现各农业管理措施下土壤食物网稳定性为:常规耕作>免耕、不施氮肥>施氮肥、集成管理>常规管理。食物网分室理论证明0.2 < p < 0.6(p为易利用的碎屑占总碎屑比例)时,土壤食物网更稳定[22],这暗示了可以通过调节土壤食物网的基质来调节食物网的稳定性,这与实际土壤食物网管理措施不谋而合[73]。今后,应加大这方面的研究,加大土壤食物网理论对实践的指导作用。
| [1] | McCann K S. The diversity-stability debate. Nature, 2000, 405(6783): 228-233. |
| [2] | Ives A R, Carpenter S R. Stability and diversity of ecosystems. Science, 2007, 317(5834): 58-62. |
| [3] | May R M. Will a large complex system be stable?. Nature, 1972, 238(5364): 413-414. |
| [4] | Dunne J A, Brose U, Williams R J, Martinez N D. Modeling food-web dynamics: complexity-stability implications // Belgrano A, Scharler U M, Dunne J A, Ulanowicz R E eds. Aquatic Food Webs: an Ecosystem Approach. Oxford: Oxford University Press, 2005:117-129. |
| [5] | Cao Z P. Future orientation of ecological agriculture. Chinese Journal of Eco-Agriculture, 2013, 21(1): 29-38. |
| [6] | Moore J C, de Ruiter P C. Energetic Food Webs: an Analysis of Real and Model ecosystems. Oxford: Oxford University Press, 2012. |
| [7] | Fu S L, Zou X M. Coleman D. Highlights and perspectives of soil biology and ecology research in China. Soil Biology and Biochemistry, 2009, 41(5): 868-876. |
| [8] | Chen X L, Tu Y. The effect of food web on the relationship of biodiversity and ecosystem stability. Science and Technology of West China, 2007, 13(10): 56-58. |
| [9] | Chen Y F, Cao Z P. The soil food web: structure, energy flux and stability. Acta Ecologica Sinca, 2008, 28(10): 5055-5064. |
| [10] | He J Z, Li J, Zheng Y M. Thoughts on the microbial diversity-stability relationship in soil ecosystems. Biodiversity Science, 2013, 21 (4): 1-10. |
| [11] | Huang J H. Impact of species diversity within ecosystem on stability // Biodiversity Committee of Chinese Academy of Sciences ed. Principles and Methodologies of Biodiversity Studies. Beijing: China Science and Technology Press,1994:178-191. |
| [12] | Huang J H, Han X G. Biodiversity and ecosystem stability. Biodiversity Science, 1995, 3(1):31-37. |
| [13] | Wang G H. Further thoughts on diversity and stability in ecosystems. Biodiversity Science, 2002, 10(1):126 -134. |
| [14] | Gao D, He H X. Research advances on biodiversity and ecosystem stability. Chinese Journal of Ecology, 2010, 29(12): 2507- 2513. |
| [15] | Wang S K, Sheng Q, Chu T J, Li B, Chen J K, Wu J H. Impact of invasive plants on food webs and pathways. Biodiversity Science, 2013, 21 (3): 249-259. |
| [16] | DeAngelis D L, Bartell S M, Brenkert A L. Effects of nutrient recycling and food-chain length on resilience. American Naturalist, 1989, 134(5): 778-805. |
| [17] | Hunt H W, Coleman D C, Ingham E R, Ingham R E, Elliott E T, Moore J C, Rose S L, Reid C P P, Morley C R. The detrital food web in a shortgrass prairie. Biology and Fertility of Soils, 1987, 3(1):57-68. |
| [18] | May R M. Stability and Complexity in Model Ecosystems. Princeton: Princeton University Press, 1973. |
| [19] | Moore J C. Hunt H W. Resource compartmentation and the stability of real ecosystems. Nature, 1988, 333 (6170): 261-263 |
| [20] | Moore J C, Zwetsloot H J C, de Ruiter P C. Staistical analysis and simulation modelling of the belowground food webs of two winter wheat management practices. Netherlands Journal of Agricultural Science,1990, 38(3): 303-316. |
| [21] | Moore J C, de Ruiter P C. Temporal and spatial heterogeneity of trophic interactions within below-ground food webs. Agriculture, Ecosystems and Environment, 1991, 34(1): 371-397. |
| [22] | Moore J C, Berlow E L, Coleman D C, de Ruiter P C. Dong Q, Hastings A, Johnson N C, McCann K S, Melville K, Morin P J, Nadelhoffer K, Rosemond A D, Post D M, Sabo J L, Scow K M, Vanni M J, Wall D H. Detritus, trophic dynamics, and biodiversity. Ecology Letters, 2004, 7(7): 584-600. |
| [23] | Moore J C, McCann K, de Ruiter P C. Modeling trophic pathways, nutrient cycling, and dynamic stability in soils. Pedobiologia, 2005, 49(6): 499-510. |
| [24] | de Ruiter P C, Neutel A M. Moore J C. Energetics, patterns of interaction strengths, and stability in real ecosystems. Science, 1995, 269(5228): 1257-1260. |
| [25] | Neutel A M, Heesterbeek J A P, de Ruiter P C. Stability in real food webs: weak links in long loops. Science, 2002, 296(5570): 1120-1123. |
| [26] | Moore J C, de Ruiter P C, Hunt H W. Influence of productivity on the stability of real and model ecosystems. Science, 1993, 261(5123): 906-908. |
| [27] | Neutel A M, Heesterbeek J A P, Van de Koppel J, Hoenderboom G, Vos A, Kaldeway C, Berendse F, de Ruiter P C. Reconciling complexity with stability in naturally assembling food webs. Nature, 2007, 449(7162): 599-602. |
| [28] | Moore J C, de Ruiter P C, Hunt H W, Coleman D C, Freckman D W. Microcosms and soil ecology: critical linkages between fields studies and modelling food webs. Ecology,1996, 77(3): 694-705. |
| [29] | de Ruiter P C, Veen J A, Moore J C, Brussaard L, Hunt H W. Calculation of nitrogen mineralization in soil food webs. Plant and Soil, 1993, 157(2): 263-273. |
| [30] | Moore J C, de Ruiter P C. Soil food webs in agricultural soils // Cheeke T, Coleman D C, Wall D H, eds. Microbial Ecology in Sustainable Agroecosystems. Boca Raton: the Chemical Rubber Company (CRC) Press, 2012: 63-88. |
| [31] | Naeem S. Complexity versus diversity // Levin S A ed. Encyclopedia of Biodiversity. Volume 1. Burlington: Academic Press, 2001:831-843. |
| [32] | Dell A I, Kokkoris G D, Banasek-Richter C, Bersier L F, Dunne J A, Kondoh M, Romanuk T N, Martinez ND. How do complex food webs persist in nature? // de Ruiter P C, Wolters V, Moore J C, eds. Dynamic Food Webs: Multispecies Assemblages, Ecosystem Development and Environmental Change. Burlington: Academic Press, 2005:425-436. |
| [33] | Justus J. Complexity, diversity and stability // Sarkar S, Plutynski A, eds. A companion to the philosophy of biology. Oxford: Blackwell, 2008: 321-351. |
| [34] | Montoya J M, Pimm S L. SoléR V. Ecological networks and their fragility. Nature, 2006, 442(7100): 259-264. |
| [35] | Laska M S, Wootton J T.Theoretical concepts and empirical approaches to measuring interaction strength. Ecology, 1998; 79(2):461-476. |
| [36] | Berlow E L, Neutel A M, Cohen J E, de Ruiter P C, Ebenman B, Emmerson M, Fox J W, Jansen V A A, Jones J I, Kokkoris G D, Logofet D O, McKane A J, Montoya J M, Petchey O. Interaction strengths in food webs: issues and opportunities. Journal of Animal Ecology, 2004, 73(3):585-598. |
| [37] | Neutel A M. Stability of Complex Food Webs: Pyramids of Biomass, Interactions Strengths, and Weight of Trophic Loops . Utrecht: Utrecht University, 2001. |
| [38] | Rooney N, McCann K S. Integrating food web diversity, structure and stability. Trends in Ecology and Evolution, 2011, 27(1): 40-46. |
| [39] | Pimm S L. Food webs. New York: Chapman and Hall, 1982. |
| [40] | de ruiter P C, Neutel A M, Moore J C. Biodiversity in soil ecosystems: the role of energy flow and community stability. Applied Soil Ecology, 1998, 10(3): 217-228. |
| [41] | Moore,J C, de Ruiter P C. A food web approach to disturbance and ecosystem stability // Van straalen N M, Lkke H, eds. Ecological risk assessment of contaminants in soil. New York: Chapman and Hall, 1997:157-171 |
| [42] | Pimm S L. The complexity and stability of ecosystems. Nature, 1984, 307(5949):321-326 |
| [43] | Pimm S L, Lawton J H, Cohen J E. Food web patters and their consequence. Nature, 1991, 350(6320):669-674 |
| [44] | de Ruiter P C, Neutel A M, Moore J C. Energetics and stability in belowground food webs // Polis G A, Winemiller K O, eds. Food Webs: Integration of Patterns and Dynamics. New York: Chapman Hall, 1996:201-210. |
| [45] | Yodzis P. The stability of real ecosystems. Nature, 1981, 289(5799):674-676. |
| [46] | Raffaelli D. From Elton to mathematics and back again. Science, 2002, 296(5570): 1135-1136. |
| [47] | Moore J C, de Ruiter PC. Compartmentalization of resource utilization within soil ecosystems // Gange A, Brown V, eds. Multitrophic Interactions in Terrestrial Systems. Oxford: Blackwell Science, 1997:375-393. |
| [48] | Moore J C. Walter D E. Hunt H W. Arthropod regulation of micro- and mesobiota in below-ground detrital food webs. Annual Review of Entomology,1988, 33(1):419-439. |
| [49] | Post D M, Conners M E, Goldberg D S. Prey preference by a top predator and the stability of linked food chains. Ecology, 2000, 81(1):8-14. |
| [50] | Rooney N, McCann K, Gellner G. Moore J C. Structural asymmetry and the stability of diverse food webs. Nature, 2006, 442(7100):265-269. |
| [51] | de Ruiter P C, Wolters V, Moore J C. Winemiller K O. Food web ecology: playing Jenga and beyond. Science, 2005, 309(5731): 68-70. |
| [52] | Moore J C, de Ruiter P C. Invertebrates in detrital food webs along gradients of productivity // Coleman D C, Hendrix P F, eds, Invertebrates as Webmasters in Ecosystems. Oxford: CABI publishing, 2000:161-184. |
| [53] | Post D M. The long and short of food-chain length. Trends in Ecology and Evolution, 2011,17(6): 269-277. |
| [54] | de ruiter P C, Neutel A M, Moore J C. The balance between productivity and food web structure in soil ecosystems // Bardgett R D, Usher M B, Hopkins D W, eds. Biological Diversity and Function in Soils. Cambridge: Cambridge University Press, 2005:139-153. |
| [55] | Hutchinson G E. Homage to Santa Rosalia, or why are there so many kinds of animals?. American Naturalist, 1959, 93(870):145-159. |
| [56] | Pimm S L, Lawton J H.Number of trophic levels in ecological communities. Nature, 1977, 268(5618):329-331. |
| [57] | Dekker S C, Scheu S, Schrter D,Setla, H, Szanser, Traas T P. Towards a new gereration of dynamics soil decomposer food webs models // de Ruiter P C, Wolters V, Moore J C, eds. Dynamic Food Webs: Multispecies Assemblages, Ecosystem development, and environmental change. Burlington: Academic Press, 2005:258-266. |
| [58] | Brussaard L, Bouwman L A, Geurs M, Hassink J, Zwart K B. Biomass, composition and temporal dynamics of soil organisms of a silt loam sil under cnventional and integrated management. Netherlands Journal of Agricultural Science, 1990, 38(3):283-302. |
| [59] | Bengtsson J, Berg P. Variability in soil food web structure across time and space // de Ruiter P C, Wolters V, Moore J C, eds. Dynamic Food Webs: Multispecies Assemblages, Ecosystem development, and environmental change. Burlington: Academic Press, 2005:201-210. |
| [60] | Irvine L, Kleczkowski A, Lane A M J, Pitchford J W, Caffrey D, Chamberlain P M. An integrated data resource for modelling the soil ecosystem. Applied Soil Ecology, 2006, 33(2):208-219. |
| [61] | de Ruiter P C, Bloem J, Bouwman L A, Didden W A M, Hoenderboom G H J, Lebbink G, Marinissen J C Y, de Vos J A, Vreeken-Buijs M J, Zwart K B, Brussaard L. Simulation of dynamics in nitrogen mineralisation in the belowground food webs of two arable farming systems. Agriculture, Ecosystems and Environment.1994, 51(1/2):199-208. |
| [62] | Montoya J M, Yvon-Durocher G. Ecological networks: information theory meets Darwin's entangled bank. Current Biology, 2007, 17(4):128-130. |
| [63] | Gao S J. Chen L S. Permanence and global stability for single-species model with three life stages and time delay. Acta Mathematica Scientia, 2006, 26(4):527-533. |
| [64] | Palamara G M, Zlatic V, Scala A. Caldarelli G. Population dynamics on complex food webs. Advances in Complex Systems, 2011, 14(4): 635-647. |
| [65] | Brose U, Williams R J, Martinez N D. Allometric scaling enhances stability in complex food webs. Ecology Letters, 2006, 9(11): 1228-1236. |
| [66] | Brouse U, Dunne J A. Modelling the dynamics of complex food webs // Verhoef H A, Morin P J, eds. Community Ecology: Processes, Models, and Applications. Oxford: Oxford University Press, 2009:37-44. |
| [67] | Ings T C, Montoya J M, Bascompte J, Blüthgen N, Brown L E, Dormann C F, Edwards F K, Figueroa D, Jacob U, Jones J I, Lauridsen R B, Ledger M E, Lewis H M, Olesen J M, Frank van Veen F J, Warren P H, Woodward G. Ecological networks-beyond food webs. Journal Animal Ecology, 2009, 78(1): 253-269. |
| [68] | Mougi A. Kondoh M. Diversity of interaction types and ecological community stability. Science, 2012, 337(6092):349-351 |
| [69] | Chen X, Cohen J E. Global stability, local stability and permanence in model food webs. Journal of Theoretical Biology, 2001, 212(2): 223-235. |
| [70] | Hastings A. Timescales, dynamics, and ecological understanding. Ecology, 2010, 91(12): 3471-3480. |
| [71] | Niwa S, Mariani L, Kaneko N, Okada H, Sakamoto K. Early-stage impacts of sika deer on structure and function of the soil microbial food webs in a temperate forest: a large-scale experiment. Forest Ecology and Management, 2011, 261(3): 391-399. |
| [72] | Xu M G, Zhang H M, Zhang F D. Evolution of soil fertility of China. Beijing: China agriculture press, 2006. |
| [73] | Chen Y F, Hu C, Li S L, Qiao Y. Managing farmland soil food web: principles and methods. Acta Ecologica Sinca, 2011, 31(1): 286-292. |
| [5] | 曹志平.生态农业未来的发展方向.中国生态农业学报,2013, 21(1): 29-38. |
| [8] | 陈秀莉,图雅.食物网与物种多样性和系统稳定性研究.中国西部科技, 2007, 13(10): 56-58. |
| [9] | 陈云峰,曹志平.土壤食物网:结构,功能及稳定性.生态学报, 2008, 28(10): 5055-5064. |
| [10] | 贺纪正,李晶,郑袁明.土壤生态系统微生物多样性——稳定性关系的思考.生物多样性, 2013, 21 (4): 1-10. |
| [11] | 黄建辉.生态系统内的物种多样性对稳定性的影响//中国科学院生物多样性委员会.生物多样性研究的原理与方法.北京:中国科学技术出版社,1994:178-191 |
| [12] | 黄建辉,韩兴国.生物多样性和生态系统稳定性.生物多样性,1995,3(1):31-37. |
| [13] | 王国宏.再论生物多样性与生态系统的稳定性.生物多样性,2002,10(1): 126 -134. |
| [14] | 高东,何霞红.生物多样性与生态系统稳定性研究进展. 生态学杂志, 2010, 29(12): 2507- 2513. |
| [15] | 王思凯,盛强,储忝江,李博,陈家宽,吴纪华.植物入侵对食物网的影响及其途径. 生物多样性,2013, 21 (3): 249-259. |
| [63] | 高淑京,陈兰荪.具有三个成长阶段的单种群时滞模型的永久持续生存和全局稳定性.数学物理学报,2006, 26(4):527-533. |
| [72] | 徐明岗,梁国庆,张夫道.主编.中国土壤肥力演变.北京:中国农业出版社,2006. |
| [73] | 陈云峰,胡诚,李双来,乔艳.农田土壤食物网管理的原理与方法. 生态学报, 2011,31(1): 286-292. |