 2014, Vol. 34
2014, Vol. 34文章信息
- 展茂魁, 杨忠岐, 王小艺, 张彦龙, 克热曼
- ZHAN Maokui, YANG Zhongqi, WANG Xiaoyi, ZHANG Yanlong, Kereman SEMI
- 肿腿蜂类寄生蜂室内控害效能评价——以松脊吉丁肿腿蜂为例
- Evaluation on control efficiency of bethylid parasitoids on pest insects indoor:a case of Sclerodermus sp. (Hymenoptera:Bethylidae)
- 生态学报, 2014, 34(9): 2411-2421
- Acta Ecologica Sinica, 2014, 34(9): 2411-2421
- http://dx.doi.org/10.5846/stxb201308062037
-
文章历史
- 收稿日期:2013-8-6
- 修订日期:2014-2-13






2. 新疆林科院森林生态研究所, 乌鲁木齐 830001
2. Research Institute of Forest Ecology, Xinjiang Forestry Academy, Urumchi 830001, China
天敌昆虫品质和控害效果的定量评价一直以来是生物防治领域的一个难题,生命表分析法[1, 2, 3, 4]虽然是目前比较好的定量评价天敌控害作用的方法,但它更适用于天敌释放后的林间、田间害虫防控效果评价。目前,在天敌资源开发、利用于生物防治中还未见到有关评选最佳天敌种类方法的报道。随着天敌资源的不断发现,将会有越来越多的天敌被发掘出来,而选择出生物防治可利用的优秀天敌种类(或品系)、达到既能有效控制害虫同时又节省天敌资源,成为利用天敌昆虫生物防治领域面临的重要课题。
肿腿蜂这类多寄主型寄生蜂具有较强的学习能力[5],能通过自身学习行为提高对目标害虫和生活环境的适应性[6],达到良好的防治效果,王小艺等[5]、张彦龙等[7]分别研究了白蜡吉丁肿腿蜂(Sclerodermus pupariae Yang et Yao)对栗山天牛(Massicus raddei Blessig)低龄幼虫的寄生作用和松褐天牛肿腿蜂(Sclerodermus alternatusi Yang)[8]对寄主松褐天牛3龄幼虫的功能反应(functional response),为肿腿蜂类寄生蜂室内控害效能评价体系的形成打下了基础。
由于松材线虫[Bursaphelenchus xylophilus (Steiner et Buhrer)]是目前对我国林业危害最为严重的外来入侵有害生物,对其进行防治的有效手段是控制其传播媒介昆虫——松褐天牛(Monochamus alternatus Hope)。为此,开展了生物防治松褐天牛技术研究,在浙江省余姚市梁弄镇调查其天敌时,在松褐天牛和日本松脊吉丁危害的马尾松树干中发现了一种寄生日本松脊吉丁(Chalcophora japonica Gory)(以下简称松脊吉丁)幼虫的肿腿蜂,经杨忠岐教授鉴定为肿腿蜂科硬皮肿腿蜂属(Sclerodermus)的一个新种(待发表)。经研究发现,该种肿腿蜂也对松褐天牛幼虫具有良好的寄生效果。为了开发利用新发现的肿腿蜂新种这个天敌资源防治松褐天牛,依据构建的肿腿蜂类寄生蜂室内控害效能评价体系,采用直接观察法[9]开展了松脊吉丁肿腿蜂(Sclerodermus sp.)对松褐天牛3龄幼虫的控害效能评测研究。该评价体系包括供试虫源标准,寄生蜂生物学与寄生行为学观察,寄生蜂对寄主的功能反应和寻找效应以及寄生蜂的忠岐指数(Y)4个部分。
1 材料与方法 1.1 供试昆虫松褐天牛3龄幼虫,2012年7月采自浙江省余姚市梁弄镇雅贤村(29°57.715′ N,121°03.873′ E,海拔142.10 m)松林中受其危害的马尾松(Pinus massoniana)树干中。幼虫头壳宽度2.5×10-3 m左右,体长2.5×10-2 m左右[10, 11]。
松脊吉丁肿腿蜂(Sclerodermus sp.),2012年5月采自浙江省余姚市梁弄镇沿山(29°57′ N,121°04′ E,海拔127.00 m)受松褐天牛和日本松脊吉丁危害的马尾松树干中,从日本松脊吉丁幼虫尸体旁的肿腿蜂茧中养出,供试肿腿蜂雌蜂为室内繁育的第3代松脊吉丁肿腿蜂种群,为羽化后交配过的雌蜂。
1.2 松脊吉丁肿腿蜂生物学特性和寄生行为学观测将直径约为5.0×10-2 m马尾松枝锯成5.0×10-1 m的木段,两端用石蜡封住以防失水。用雕刻刀将木段树皮切成“U”字形切口,直至木质部,然后将该“U”形树皮揭起但不割断,再在表皮下的木质部挖长、宽、深为3.0×10-2 m×0.5×10-2 m×0.5×10-2 m的小凹槽,然后将上述供试松褐天牛3龄幼虫放入该凹槽中,最后将揭起的树皮盖回,再用石蜡或用Para film膜封上切口缝以保湿,令天牛幼虫在木段内自行蛀食。最后将接入松褐天牛幼虫的马尾松树木段放入对应编号的500目尼龙网纱做成的袋子内,待松褐天牛蛀出虫粪和锯末后接入松脊吉丁肿腿蜂雌蜂,每天观察松脊吉丁肿腿蜂和松褐天牛活动情况,连续观察50d,记录寄生蜂的产卵时间、产卵量、结茧时间、羽化时间、出蜂量、雌雄性比。对照组不接寄生蜂,以观察天牛幼虫的自然死亡率。
试验期间室内温度为(20±5)℃,相对湿度为(70±5)%。
1.3 功能反应试验功能反应是指每个天敌的控制效能随猎物密度变化的一种反应,即天敌对猎物的控制效应[12, 13, 14, 15, 16, 17, 18]。
1.3.1 单头松脊吉丁肿腿蜂对松褐天牛3龄幼虫的功能反应松褐天牛幼虫接入马尾松树木段并装袋接入肿腿蜂的方法同1.2,每根/袋木段上分别接1、2、3、4、5、6头松褐天牛幼虫,每个处理重复5次,每袋接入松脊吉丁肿腿蜂雌蜂1头,对照组不接肿腿蜂,连续观察50d,每天观察记录数据。试验期间室内温湿度同1.2。
1.3.2 松脊吉丁肿腿蜂自身密度对松褐天牛3龄幼虫的功能反应方法同1.2,每根/袋木段接5头松褐天牛幼虫,每袋接入松脊吉丁肿腿蜂雌蜂1、2、3、4、5、6头,每个处理重复5次,对照组不接肿腿蜂,连续观察50d,每天观察记录数据。试验期间室内温湿度同1.2。
1.4 数据分析 1.4.1 功能反应由于松脊吉丁肿腿蜂雌蜂在产卵前需先刺蛰麻痹寄主,然后吸食天牛幼虫血淋巴补充营养,以供其卵巢和卵发育的需要。如果寄主个体较小,肿腿蜂在补充营养过程中将致死一定数量的寄主幼虫,因此,本研究将寄生蜂补充营养与寄生对寄主幼虫致死作用的数据进行分别处理[5, 7]。首先对试验结果进行散点图的初步拟合,观察与哪一类的功能反应模型符合,接近HollingⅠ型方程的用公式(1)进行拟合,接近Holling Ⅱ型方程的用公式(2) 进行拟合。
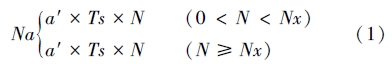
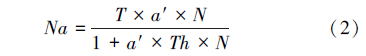
式中,Na为被寄生的寄主数量(头),N为初始寄主密度(头),Nx为被寄生的寄主数量上限(头),a′为瞬时攻击率(头/d),Ts为寻找寄主时间(d),T是试验处理时间(d),Th为寄主处理时间(d/头)[5, 19, 20]。
1.4.2 寄主不同密度下寄生蜂的寻找效应寻找效应是指天敌在寄生或捕食过程中对于猎物攻击的行为效应[21]。
肿腿蜂雌蜂对寄主的寻找效应参考捕食性天敌寻找效应的计算方法,按下面公式(3)计算,寄生作用的寻找效应按公式(4)计算[5]:
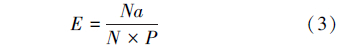
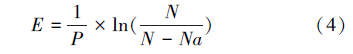
式中,E为寻找效应,Na为被致死或寄生的寄主数量(头),N为寄主密度,P为天敌密度。
1.4.3 寄生蜂自身密度对松褐天牛3龄幼虫的功能反应首先对试验数据进行散点图初步拟合,根据其符合的数学模型用DPS软件进行参数估计和方程的显著性检验。
1.4.4 寄生蜂不同密度下的寻找效应肿腿蜂密度对寻找效应的影响根据Hassell模型[22, 23]进行拟合[19]:
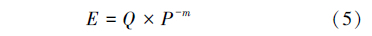
式中:E为寻找效应,Q为寻找参数,m为相互干扰参数,P为寄生蜂的密度。
1.4.5 寄生蜂的忠岐指数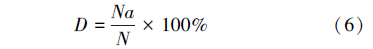
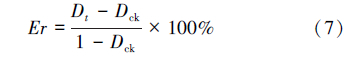
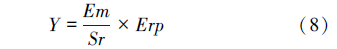
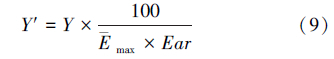
式中,D为虫口减退率,Na为被致死的寄主数量;N为试验初寄主的数量;Er为相对防效;Dt为处理组虫口减退率;Dck为对照组虫口减退率;忠岐指数 Y为寄生蜂最佳释放比例时每头雌蜂产生的下一代子代蜂寄生寄主的平均数量(小写的 y表示其他释放比时的忠岐指数,以示区别);Em为寄生蜂在1头寄主上羽化出的平均雄蜂量;Sr为寄生蜂最佳释放比时子代的雄雌性比;Erp为寄生蜂最佳释放比时的相对寄生率Er;Y′为忠岐指数的标准值,作为寄生蜂控害效能分级量化依据; max为该寄生蜂的平均最大产卵量;Ear为平均羽化率。
肿腿蜂类寄生蜂室内控害效能评价标准的量化指标采用黄金分割法划分成:优秀 (Y′≥32.6228,图 1中蓝色区域)、良好(32.6228>Y′≥23.6076,图 1中绿色区域)、中等(23.6076>Y′≥12.4684,图 1中灰色区域)、较差(12.4684>Y′≥9.0228,图 1中黄色区域)、淘汰(Y′<9.0228,图 1中红色区域)五级。参与控害效能评价的寄生蜂的Y值与忠岐指数标尺 Y′值的换算关系为公式(9)。

|
| 图 1 忠岐指数标尺 Fig. 1 Scale of Zhongqi Index |
所有数据统计均采用DPS(Version 7.05)软件[24]。试验结果经方差分析后进行Duncan′s多重比较检验各处理间的差异显著性。
2 结果与分析 2.1 松脊吉丁肿腿蜂的生物学特性和寄生行为学观测松脊吉丁肿腿蜂(图 2)性二型现象明显,雄蜂绝大多数有翅,雌蜂无翅,属抑性寄生性。雌蜂先爬行寻找在树皮下危害、活动的寄主,在定位寄主后,用上颚在树皮上咬一圆形入侵孔(图 3),进入寄主坑道,然后爬附到寄主幼虫体上,先利用产卵器刺蜇寄主并将分泌的毒液注入松褐天牛幼虫体内,使其永久性麻痹,然后才在该不食不动的寄主体上产卵。产卵前,雌蜂需先吸取寄主幼虫血淋巴补充营养,以完成卵巢和卵的发育。如果寄主个体较小,则雌蜂在这个产卵前的补充营养阶段吸食时会致死一定数量的寄主幼虫。此过程中雌蜂腹部随着卵巢和卵的发育逐渐增大、伸长(图 2-4),卵发育成熟后被产在寄主幼虫的体表。产卵后,雌蜂一直守候在寄主体上及时清理其后代卵、幼虫和寄主体表的木屑等杂物,帮助后代幼虫调整取食部位使其得以健全地发育,在子代蜂即将羽化时雌蜂还会帮助其咬破茧壳实现成功羽化[25] ,表现出良好的抚幼习性(brood guarding behavior)[26]。观察发现,松脊吉丁肿腿蜂的羽化率 (Ear) 为(99.55±0.42)%,其中有1个刻槽中滋生螨虫出蜂率仍达66.67%,平均出蜂量为(93.38±18.43)头,雌雄比为42.16 ∶ 1,单头寄主最大出蜂量为134头。从1头寄主上育出的子代蜂绝大部分为雌性,而雄蜂通常为2—3头,至少为1头,多者可达7头。如此高的雌蜂比例和良好的种群保持能力表明该种肿腿蜂为优秀的寄生蜂。

|
| 图 2 松脊吉丁肿腿蜂 1: 有翅雄蜂;2: 无翅雄蜂;3: 补充营养前的雌蜂;4: 补充营养后卵和卵巢充分发育的雌蜂 Fig. 2 Sclerodermus sp. adults |

|
| 图 3 肿腿蜂咬啮入侵孔进入松褐天牛幼虫坑道寄生寄主 1: 肿腿蜂雌蜂正在咬啮入侵孔;2: 肿腿蜂雌蜂咬啮入侵孔蛀出的木屑;3: 肿腿蜂啮咬出的入侵孔 Fig. 3 Bethylid female wasp bites entrance hole to enter host′s gallery to parasitize her host |
松脊吉丁肿腿蜂寄生过程中,从释放寄生蜂起至在寄主体上产第1粒卵的时间记为补充营养时间;从第1粒卵出现到寄生在寄主体上的肿腿蜂幼虫完成幼虫发育结第1个茧开始化蛹的时间记为实际寄生时间;从第1粒卵产出到第1头子代蜂成功羽化出来的时间记为发育1代之历期;从释放肿腿蜂开始到第1头子代蜂成功羽化的时间记为1个寄生周期,测定结果见表 1。
2.2 松脊吉丁肿腿蜂对松褐天牛3龄幼虫的功能反应 2.2.1 单头松脊吉丁肿腿蜂对松褐天牛3龄幼虫的致死作用的功能反应松脊吉丁肿腿蜂在数量为1头的情况下,对松褐天牛3龄幼虫的致死作用随着寄主密度的增加而显著提高(df=5,29,F=11.45,P=0.0001)。
|
补充营养时间
The supplementary nutrition time of the bethylid wasp/d | 寄生时间
Parasitizing time/d | 发育一代历期
The developing period for a generation/d | 寄生周期
A parasitism cycle/d |
| 试验期间温度为(20±5)℃,相对湿度为(70±5)% | |||
| 4.63±1.06 | 13.43±2.02 | 32.54±2.38 | 37.11±2.66 |

|
| 图 4 松脊吉丁肿腿蜂对松褐天牛3龄幼虫致死作用的功能反应 Fig. 4 Functional response for killing action of Sclerodermus sp. on the 3rd instar larvae of M. alternatus |
经模型拟合(图 4),松脊吉丁肿腿蜂对松褐天牛3龄幼虫致死作用功能反应符合Holling Ⅱ型方程:
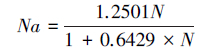
df=1,5,F=55.8621,P=0.0017,r=0.9660
其中,瞬时攻击率a′为0.2778头/d,寄主处理时间Th为2.3144d/头,理论上21d内,1头松脊吉丁肿腿蜂对松褐天牛3龄幼虫的最大致死量为9.07头,平均每日最大致死量为0.43头。
2.2.2 单头松脊吉丁肿腿蜂对松褐天牛3龄幼虫寄生作用的功能反应如图 5所示,单头松脊吉丁肿腿蜂对松褐天牛3龄幼虫的平均寄生数量随寄主密度增加而升高,最大平均寄生量为1头,经模型拟合,松脊吉丁肿腿蜂对松褐天牛3龄幼虫的寄生作用功能反应符合HollingⅠ型方程,这可能是由于肿腿蜂特有的抚幼习性所致。

|
| 图 5 松脊吉丁肿腿蜂对松褐天牛3龄幼虫寄生作用的功能反应 Fig. 5 Functional response of Sclerodennus sp. parasitizing the 3rd instar larvae of M. alternatus |

|
| 图 6 松脊吉丁肿腿蜂寄生过程中的3种虫态 1:在寄主体上产卵,箭头所指为肿腿蜂雌蜂在松褐天牛幼虫体上产的卵粒;2:寄生在松褐天牛幼虫体上的肿腿蜂幼虫及抚幼母蜂;3:新羽化的子代肿腿蜂成虫 Fig. 6 Three stages of Sclerodennus sp. during the parasitism progress |
当0<N<3时,Na=0.2N(df=1,1,F=+∞,P<0.0001,r=1);当N=3时,Na=0.27N;当N≥4时,Na=1。本研究中,松脊吉丁肿腿蜂对寄主的致死密度阈限Nx为4头,即释放到松树木段上1头松脊吉丁肿腿蜂最多可致死4头松褐天牛3龄幼虫,其中有1头是被寄生所致死的,另3头为雌蜂补充营养时的取食所致死的。当寄生作用的寻找时间Ts即实际补充营养时间为4.50d时,该蜂寄生作用的瞬时攻击率a′为0.05556头/d。

|
| 图 7 不同寄主密度下松脊吉丁肿腿蜂对松褐天牛3龄幼虫致死作用的寻找效应 Fig. 7 Searching efficiency in killing action of Sclerodennus sp. on the 3rd instar larvae of M. alternatus |
松脊吉丁肿腿蜂雌成虫对寄主致死作用的寻找效应参考捕食性天敌的寻找效应计算方法,按数据分析中公式(3)进行计算。结果如图 7所示,随着寄主密度的增加,单头松脊吉丁肿腿蜂的致死作用寻找效率直线下降(y=-0.0860x+1.1160,df=1,5,F=49.2129,P=0.0022,r=- 0.9617)。
2.2.4 寄主松褐天牛3龄幼虫密度对单头松脊吉丁肿腿蜂寄生作用的寻找效应

|
| 图 8 不同寄主密度下松脊吉丁肿腿蜂对松褐天牛3龄幼虫寄生作用的寻找效应 Fig. 8 Searching efficiency in parasitism of Sclerodermus sp. on the 3rd instar larvae of M. alternatus |
松脊吉丁肿腿蜂寄生作用的寻找效应按数据分析中公式(4)计算。如图 8所示,单头寄生蜂寄生作用寻找效率随着寄主密度的增加先上升后下降(y=-0.0145x2+0.0948x+0.1292,df=2,3,F=4.5656,P=0.1230,r= 0.8676)。这说明当寄主密度较低时无法满足寄生蜂的营养需要,其寄生作用的寻找效应相应增加;而随着寄主密度的增加,肿腿蜂雌蜂充分补充营养而达到自身极限,肿腿蜂雌蜂卵巢和卵发育成熟又不得不产卵,再加上肿腿蜂良好的抚幼习性,雌蜂产卵后和子代羽化前不再寻找其他寄主寄生,寄生数量不再增加;当寄主密度继续增加时,寻找效应公式(4)的基数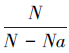 增加,而肿腿蜂的密度为1头不变,其寻找效应必然下降。
增加,而肿腿蜂的密度为1头不变,其寻找效应必然下降。
当y值最大时,x为3.27,这表明该寄生蜂具有较强的搜索扩散能力,并再次印证了1头该肿腿蜂能够防治3—4头松褐天牛3龄幼虫,与2.2.3的结果相同。
2.2.5 寄主松褐天牛3龄幼虫密度对单头松脊吉丁肿腿蜂产卵量的影响不同寄主密度下松脊吉丁肿腿蜂的产卵量见表 2。方差分析结果显示(表 2),0<N<3时,各处理的寄生蜂产卵量显著低于N≥3时的产卵量(P =0.0101),而N≥3时,各处理的寄生蜂产卵量之间差异不显著(P =0.1004)。由此可知,1头松脊吉丁肿腿蜂雌成虫需要3—4头松褐天牛3龄幼虫才能得到充分营养补充并产出一定数量的卵,这说明1头雌蜂成虫能够防治3—4头松褐天牛3龄幼虫。
|
寄主密度
Host density /(头/袋) | 平均产卵量
Mean egg number laid by a female of Sclerodermus sp. bethylid/(粒/袋) |
| 表 中同一列数据(平均值±SE)后不同小写字母表示各处理间经多重比较(Duncan′s新复极差法)在0.05水平上差异显著 | |
| 1 | 15.20±15.20b |
| 2 | 30.00±18.54b |
| 3 | 70.60±17.84a |
| 4 | 94.20±2.48a |
| 5 | 105.20±4.94a |
| 6 | 105.20±9.18a |
当寄主密度固定为5头,随着松脊吉丁肿腿蜂雌成虫密度增加到3头时,寄主的致死率显著上升(df=2,14,F=7.4000,P=0.0081)至100%,二者呈明显的线性关系(y=0.14x+0.5733,df=1,2,F=147.0000,P=0.02768,r= 0.9966)。当雌蜂密度继续上升时,其对寄主的致死率保持100%。
松脊吉丁肿腿蜂不同密度下对松褐天牛3龄幼虫致死率经二次多项式回归分析得方程:y=-0.02212x2+0.2087x+0.5320(df=2,3,F=35.1342,P=0.0083,r= 0.9793)。从图 8得知,当y值最大时,x为4.72,即当肿腿蜂的释放比(肿腿蜂雌蜂数量:松褐天牛3龄幼虫数量)接近1 ∶ 1(4.72 ∶ 5)时防治效果最佳,这表明林间防治松褐天牛时,松脊吉丁肿腿蜂释放比为1 ∶ 1的性价比最高。

|
| 图 9 不同密度的松脊吉丁肿腿蜂补充营养取食对松褐天牛3龄幼虫的致死率 Fig. 9 Killing ratio of Sclerodermus sp. female at different densities on the 3rd instar larvae of M. alternatus by supplementary nutrition of female bethylid |
当寄主密度固定为5头时,随着肿腿蜂密度的增加,被寄生的寄主数量上升,如图 10所示,二者呈明显的抛物线关系:y=-0.0643x2+0.7986x+0.2800(df=2,3,F=176.0733,P=0.0008,r= 0.9958)。

|
| 图 10 不同密度的松脊吉丁肿腿蜂对松褐天牛3龄幼虫寄生作用的功能反应 Fig. 10 Functional response for parasitatism of Sclerodermus sp. at different densities on the 3rd instar larvae of M. alternates |
不同密度下肿腿蜂对松褐天牛3龄幼虫致死作用的寻找效应根据公式(3)得出相应的寻找效应E值,然后按Hassell[21]模型拟合得方程:E=0.741×P-0.80。
随着松脊吉丁肿腿蜂密度的增加,其对松褐天牛3龄幼虫致死作用的寻找效应显著下降(df=1,5,F=21.8834,P=0.0095,r=-0.9955)(图 11)。

|
| 图 11 不同密度的松脊吉丁肿腿蜂对松褐天牛3龄幼虫致死作用的寻找效应 Fig. 11 Searching efficiency for killing action of Sclerodermus sp. at different densities on the 3rd instar larvae of M. alternates by supplementary nutrition of female bethylid |
不同密度下肿腿蜂对松褐天牛3龄幼虫寄生作用的寻找效应根据公式(4)得出相应的寻找效应E值,然后按Hassell[21]模型拟合,拟合方程为:E=0.232×P-0.26 (df=1,5,F=95.8070,P=0.0006,r=-0.9469)。如图 12所示,随着松脊吉丁肿腿蜂密度的增加,其对松褐天牛3龄幼虫寄生作用的寻找效应显著下降。

|
| 图 12 不同密度的松脊吉丁肿腿蜂对松褐天牛3龄幼虫寄生作用的寻找效应 Fig. 12 Searching efficiency for parasitism of Sclerodermus sp. at different densities on the 3rd instar larvae of M. alternatus |
当寄主密度为5头不变时,肿腿蜂不同密度下的平均产卵量差异不显著(df=5,61,F=2.2650,P=0.0603),经模型拟合得方程:y=-4.4885x+111.8864(df=1,5,F=68.9031,P=0.0012,r=-0.9722),这表明二者呈明显的线性关系(图 13)。y值最大为111.8864,即松脊吉丁肿腿蜂的平均最大产卵量( max)为111.89粒,表明该肿腿蜂具有较高的生殖能力。

|
| 图 13 不同密度的松脊吉丁肿腿蜂的产卵量 Fig. 13 Egg number laid by female bethylid at different densities |
依据2.4.1得出的结果,松脊吉丁肿腿蜂防治松褐天牛3龄幼虫的最佳释放比为1 ∶ 1。将试验中寄生蜂释放比为1 ∶ 1的处理组(1.3.1中的蜂虫比为1 ∶ 1的处理组和1.3.2中蜂虫比为5 ∶ 5的处理组)联合统计,按照数据分析中公式(6)、(7)、(8)和(9)计算该松脊吉丁肿腿蜂对松褐天牛3龄幼虫的相对防效、相对寄生率、忠岐指数及其标准值。1 ∶ 1比例释放该肿腿蜂时,对寄主的相对寄生率为44.45%,平均每头寄主的出蜂量为(91.29±19.12)头,本研究中该蜂的雌雄性比为41.60 ∶ 1,松脊吉丁肿腿蜂防治松褐天牛幼龄幼虫的忠岐指数Y为39.63,其Y′为35.5788,对应忠岐指数标尺属优秀级,这说明该蜂对松褐天牛3龄幼虫控害效能优秀。
3 结论与讨论单头松脊吉丁肿腿蜂对松褐天牛3龄幼虫寄生作用功能反应的测定结果与对寄生作用寻找效应的测定结果以及寄主密度对单头寄生蜂产卵量影响的测定结果一致(单头松脊吉丁肿腿蜂能够防治3—4头松褐天牛3龄幼虫),再次表明该蜂具有较强的控害潜力,该评价体系是稳定可靠的。
松脊吉丁肿腿蜂自身密度对松褐天牛3龄幼虫致死作用功能反应影响的测定结果反映防治松褐天牛时,松脊吉丁肿腿蜂的释放比为1 ∶ 1(4.72 ∶ 5)时的控害效能最大,这主要是肿腿蜂类寄生蜂雌蜂良好的抚幼习性所致,与寄生蜂生物学特性、寄生行为学观察结果和王小艺等[5]以及张彦龙等[7]研究结果相同。通过对王小艺等[5]研究结果分析,得白蜡吉丁肿腿蜂控害效能标准值Y′值为31.4258,属对栗山天牛低龄幼虫控害效能良好的寄生蜂;与之相比,松脊吉丁肿腿蜂则是对松褐天牛低龄幼虫控害效能优秀的寄生蜂。
虽然本研究是在室内试验条件下进行的,其结果和林间实验结果可能会存在一定的差异,但是该方法是准确地定量评价寄生蜂控害效能的可行方法,该评价体系可用于筛选寄生蜂最佳种类以应用于生物防治,包括4个部分。
(1) 供试虫源标准
寄生蜂:选取实验室内人工繁育的第3代雌蜂进行试验,确保天敌资源的自然品质,以减少传代引起的退化[27, 28, 29]造成的影响。寄主:即将进入木质部的害虫的幼龄幼虫,如松褐天牛的3龄幼虫,刘柱东等[30]报道了寄主个体大小影响寄生蜂的致死率和寄生率,管氏肿腿蜂更喜欢一定范围内较大的个体,魏可等[31]研究了白蜡吉丁肿腿蜂对白蜡窄吉丁个体大小的偏好是呈抛物线变化。3龄松褐天牛幼虫属中龄幼虫,不仅营养充足,而且个体大小为肿腿蜂适宜处理的阈值内,更重要的是进入木质部的松褐天牛幼虫绝大多数为4龄以上的老熟幼虫,肿腿蜂很难穿透木质层到达松褐天牛蛹室,因此3龄幼虫为最佳试验虫态。
确立此虫态作为标准试虫是本评价体系的基础,是确保天敌品质和开展生物防治应用最重要之处,这关系到天敌应用效能和生物防治的成功与否。如果利用活力小的或劣次的天敌昆虫等生物防治物终将不会发挥应有的效能,这也是当前生物防治应用尚未超越化学防治的重要原因,因此供试虫源标准应当恪守,不宜随意变更。
(2) 寄生蜂生物学特性和寄生行为学观测
观察寄生蜂生物学特性和寄生行为学是了解天敌习性的根本方法和必不可少的内容,是开发天敌资源的基础,需认真作好。
(3) 寄生蜂对寄主的功能反应和寻找效应
通过测定单头寄生蜂对不同密度寄主以及不同密度的寄生蜂对寄主致死作用的功能反应、寄生作用的功能反应,致死作用的寻找效应、寄生作用的寻找效应以及对寄生蜂产卵量的影响,评价不同释放密度时寄生蜂对寄主的最大致死潜力、最大寄生量、致死作用的搜索能力、寄生作用的搜索能力和发挥防治作用并保证后代发育时的防治效价,从而得出寄生蜂最佳释放比,保证寄生蜂应用的性价比最大化。
其中,寄生蜂密度、寄主密度和二者之间比例的设定应均匀并且连续,各处理组需设置多个重复。如本研究中不同密度的寄生蜂为1、2、3、4、5、6头/袋,不同密度的寄主为1、2、3、4、5、6头/木段,每个处理5个重复,以实现观测值无限地接近寄生蜂品质的真实值。至于设定比例的限定,寄生类天敌昆虫有别于捕食类天敌昆虫,寄生性天敌昆虫产的卵只需在1个寄主个体上即可完成整个发育过程,试验时寄生蜂对寄主的最大比例值大于1即可,如本研究中寄生蜂自身密度对寄主功能反应的试验(1.3.2)中,释放的松脊吉丁肿腿蜂与松褐天牛3龄幼虫数量最大比例达6 ∶ 5,超过了1 ∶ 1的比例。而整个试验的观察时限须涵盖寄生蜂的整个寄生周期,本研究中自释放肿腿蜂开始每天观测,直至子代肿腿蜂全部羽化。
(4) 忠岐指数
忠岐指数是个综合指标,是寄生蜂品质评价体系中多个指标的集成,适用于不同种(生物型)之间比较的各自最佳释放比时每头雌蜂产生的下一代子代蜂寄生寄主的平均数量,该指数是整个评价体系的核心和依托,其计算公式为: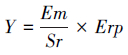 ,应用中依公式(9)转换后将 Y′ 值对照其标尺(图 1)即可得出寄生蜂的控害效能所属的分级。该指数的应用可使寄生蜂控害效能评价更加科学合理,可操作性更强,也增强了评价指标的可比性。
,应用中依公式(9)转换后将 Y′ 值对照其标尺(图 1)即可得出寄生蜂的控害效能所属的分级。该指数的应用可使寄生蜂控害效能评价更加科学合理,可操作性更强,也增强了评价指标的可比性。
公式中选定雄雌性比(Sr)而非雌雄性比的原因:其一,雌蜂数量决定着寄生蜂类天敌昆虫的种群发展趋势,将雌蜂数量放在公式分母的分母位置,转换成单分母式时雌蜂数量即可变成分子。之所以将雄雌性比放在分母位置上的另一个原因是保证雌蜂、雄蜂二者中的任何一个都不可为0,其中之一为0,种群必将于本代灭亡,自身不具有种群保持能力的寄生蜂没有开发利用价值,如此可保持公式的实际应用性也使该评价体系更符合实际。
| [1] | Kuhar T P, Wright M G, Hoffmann M P. Environmental Entomology, 2002, 31: 482-489. |
| [2] | Pang X F, Liang G W. Population system control of Pest. Guangdong Science & Technology Press, Guangzhou, 1995: 1-212. |
| [3] | Zhao X F. Biocontrol of pest insects (3rd eds.). Beijing: Chinese agriculture press, 1999: 1- 275. |
| [4] | Bellows T S Jr, van Driesche R G, Elkinton J S. Principles and Applications of Biological Control//Bellows T S, Fisher T W. eds. Handbook of Biological Control. New York: Academic Press, 1999: 199-223. |
| [5] | Wang X Y, Yang Z Q, Tang Y L, Jiang J, Gao C, Liu Y C, Zhang X W. Parasitism of Sclerodermus pupariae (Hymenoptera: Bethylidae) on the young larvae of Massicus raddei ( Coleoptera:Cerambycidae). Acta Entomogica Sinica, 2010, 53(6): 675-682. |
| [6] | Wei K, Tang Y L, Wang X Y, Yang Z Q, Cao L M, Lu J F, Liu E S, Liu G J. Effects of learning experience on behavior of the generalist parasitoid Sclerodermus pupariaeto novel hosts. Journal of Applied Entomology, 2012, 137(6): 1-7. |
| [7] | Zhang Y L, Yang Z Q, Wang X Y, Zhang Y N, Wu C J, Ma F S, Lu Z G. Functional response of the parasitoid Sclerodermus sp. (Hymenoptera: Bethylidae) to the third instar larvae of host Monochamus alternates (Coleoptera: Cerambycidae). Acta Entomologica Sinica, 2012, 55 (4): 426- 434. |
| [8] | Yang Z Q, Wang X Y, Duan Z Y, Zhang Y N, Li Y H, Cao L M, Jiang Y. A new species of Sclerodermus (Hymenoptera: Bethylidae) parasitizing larva of Monochamus alternatus (Coleoptera: Cerambycidae) from China. Annals of the Entomological Society of America, 2014: 107 (Submitted). |
| [9] | Liu S S. Methods for evaluation of predators and parasitoids in the control of Insect pests and strategy for their practical application. Chinese Journal of Biological Control, 2004, 20(1): 1-7. |
| [10] | Wang L P. Study on the biological characteristic of Monochamus alternatus Hope. Journal of Fujian Forestry Science and technology, 2004, 31(3): 23-26. |
| [11] | Zhao Y X, Dong Y, Xu Z H. Bionomics and geographical distribution of Monochamus alternatus Hope (Coleoptera: Cerambycidae) in Yunnan Province. Forest Pest and Disease, 2004, 23(5): 13-16. |
| [12] | Ge F. Modern ecology (2nd eds.). Beijing: Science Press, 2008: 117-149. |
| [13] | Price P W. Insect Ecology. New York: John Wiley & Sons Press, 1984: 73-110. |
| [14] | Holling C S. Some characteristics of simple type of predation and parasitism. Canadian Entomologist, 1959, 91(7): 385-398. |
| [15] | Ding Y Q. Mathematical model and its estimation of the main parameters in natural enemy-pest system: predator-prey model in the predator-prey systems. Entomological Knowledge, 1983, 20(4): 187-190, 186. |
| [16] | Wu W J, Liang G W. Review of methods fitting Holling disk equation. Entomological Knowledge, 1989, 11(2): 96-100. |
| [17] | Wu K J, Sheng C F, Gong P Y. Equation of predator functional response and estimation of the parameters in it. Entomological Knowledge, 2004, 41(3): 267-269. |
| [18] | Abrams P A. Functional responses of optimal foragers. The American Naturalist, 1982, 120(3): 382-390. |
| [19] | Ding Y Q. Insect mathematical ecology. Beijing: Science Press, 1994: 252-317. |
| [20] | Ding Y Q. Principles and application of insect population mathematical ecology. Beijing: Science Press, 1980: 197-213. |
| [21] | Zhang S Z, Hua B Z, Xu X L. The predatory functional responses and searching efficiency of Propylaea japonica on Rhopalosiphum maidis. Journal of Northwest Sci-Tech University of Agriculture and Forestry(Natural Science Edition), 2005, 33(5): 85- 87, 94. [22] Hassell M P, Varley G C. New inductive population model for insect parasites and its bearing on biological control. Nature, 1969, 223: 1133-1137. |
| [23] | Hassell M P, Roger D J. Insect parasite responses in the development of population models. Journal of Animal Ecology, 1972, 41(3): 661-676. |
| [24] | Tang Q Y, Feng M G. DPS Data Processing System: Experimental Design, Statistical Analysis and Data Mining (2nd eds.). Beijing: Science Press, 2007: 75-128, 656-665. |
| [25] | Zhou Z J, Yang W, Zeng C H, Yang D M, Ye W J. A Preliminary Study on the bionomics of Scleroderma sichuanensis (Hymenoptera: Bethylidae). Scientia Silvae Sinicae, 1997, 33(5): 475-480. |
| [26] | Ian C W. Brood guarding in a bethylid wasp. Ecological Entomology, 1991, 16: 55-62. |
| [27] | Dai P L, Xu Z Q. Population rejuvenation of mass-rearing Scleroderma guani. Chinese Bulletin of Entomology, 2007, 44(3): 402-405. |
| [28] | Yang Z Q. Wei J R. Wang X Y. Mass rearing and augmentative releases of the native parasitoid Chouioia cunea for biological control of the introduced fall webworm Hyphantria cunea in China. BioControl, 2006, 51(4): 401-418. |
| [29] | Xu Y X, Sun X G, Han R D, He Z. Parasitoids of Dendrolimus punctatus in China. Chinese Bulletin of Entomology, 2006, 43(6): 767-773. |
| [30] | Liu Z D, Xu B B, Li L, Sun J H. Host-size mediated trade-off in a parasitoid Sclerodermus harmandi. PLoS One, 2011, 6(8): e23260. |
| [31] | Wei K, Tang Y L, Wang X Y, Cao L M, Yang Z Q. The developmental strategies and related profitability of an idiobiont ectoparasitoid Sclerodermus pupariae vary with host size. Ecological Entomology, 2014, 39(1): 101-108. |
| [2] | 庞雄飞, 梁广文. 害虫种群系统的控制. 广州: 广东科技出版社, 1995: 1-212. |
| [3] | 赵修复. 害虫生物防治(第三版). 北京: 中国农业出版社, 1999: 1- 275. |
| [5] | 王小艺, 杨忠岐, 唐艳龙, 姜静, 高纯, 刘云程, 张显文. 白蜡吉丁肿腿蜂对栗山天牛低龄幼虫的寄生作用. 昆虫学报, 2010, 53 (6): 675-682. |
| [7] | 张彦龙, 杨忠岐, 王小艺, 张翌楠, 吴成进, 马生福, 陆之贵. 松褐天牛肿腿蜂对寄主松褐天牛三龄幼虫的功能反应. 昆虫学报, 2012, 55 (4) : 426-434. |
| [9] | 刘树生. 天敌动物对害虫控制作用的评价方法及其应用策略. 中国生物防治, 2004, 20(1): 1-7. |
| [10] | 王玲萍. 松墨天牛生物学特性的研究. 福建林业科技, 2004, 31(3): 23-26. |
| [11] | 赵宇翔, 董燕, 徐正会. 云南省松墨天牛生物学特性和地理分布研究. 中国森林病虫, 2004, 23(5): 13-16. |
| [12] | 戈峰. 现代生态学(第二版). 北京:科学出版社, 2008: 117-149. |
| [15] | 丁岩钦. 天敌-害虫作用系统中的数学模型及其主要参数估计:捕食者-猎物系统中的捕食作用模型. 昆虫知识, 1983, 20(4): 187-190, 186. |
| [16] | 吴伟坚, 梁广文. Holling圆盘方程拟合方法概述. 昆虫天敌. 1989, 11(2): 96-100. |
| [17] | 吴坤君, 盛承发, 龚佩瑜. 捕食性昆虫的功能反应方程及其参数的估算. 昆虫知识, 2004, 41(3): 267-269. |
| [19] | 丁岩钦. 昆虫数学生态学. 北京: 科学出版社, 1994: 252-317. |
| [20] | 丁岩钦. 昆虫种群数学生态学原理与应用. 北京: 科学出版社, 1980: 197-213. |
| [21] | 张世泽, 花保祯, 许向利. 龟纹瓢虫捕食玉米蚜的功能反应与寻找效应研究. 西北农林科技大学学报:自然科学版, 2005, 33(5): 85-87, 94. |
| [24] | 唐启义, 冯明光. DPS 数据处理系统: 实验设计、统计分析及数据挖掘, 第2版. 北京: 科学出版社, 2007: 75-128, 656-665. |
| [25] | 周祖基, 杨伟, 曾垂惠, 杨德敏, 叶伟军. 川硬皮肿腿蜂生物学特性的研究(膜翅目: 肿腿蜂科). 林业科学, 1997, 33(5): 475-479. |
| [27] | 代平礼, 徐志强. 人工扩繁管氏肿腿蜂的蜂种复壮研究. 昆虫知识, 2007, 44(3):402-405. |
| [29] | 徐延熙, 孙绪艮, 韩瑞东, 何忠. 我国马尾松毛虫寄生性天敌的研究进展. 昆虫知识, 2006, 43(6): 767-773. |