 2014, Vol. 34
2014, Vol. 34文章信息
- 金樑, 孙莉, 崔慧君, 赵静静, 刘喜超, 唐鹏杰, 王晓娟
- JIN Liang, SUN Li, CUI Huijun, ZHAO Jingjing, LIU Xichao, TANG Pengjie, WANG Xiaojuan
- 青藏高原东缘高寒草原有毒植物分布与高原鼠兔、高原鼢鼠的相关性
- Correlation between the distribution characteristics of poisonous plants and Ochotona curzoniae, Myospalax baileyi in the East of Tibetan Plateau Alpine meadow ecosystem
- 生态学报, 2014, 34(9): 2208-2215
- Acta Ecologica Sinica, 2014, 34(9): 2208-2215
- http://dx.doi.org/10.5846/stxb201307061849
-
文章历史
- 收稿日期:2013-7-6
- 修订日期:2013-10-29






青藏高原是世界上海拔最高的陆地生态系统,面积约占我国国土总面积的1/4,平均海拔4000—5000 m,素有“世界屋脊”之称,其境内的植被类型主要为高寒草原生态系统,是我国乃至国际上非常独特的陆地生态系统之一,在全球范围内具有典型的特殊性[1]。当前,由于超载过牧、矿产开发、乱采滥挖等人类活动的干扰,加之全球变化,降雨不均等因素,青藏高原高寒草原生态系统退化日趋严重[2],优良牧草数量不断减少,有毒植物大量扩散和蔓延,造成高寒草原生产力下降[1],其中青藏高原东缘地区具有典型的代表性,主要包括甘肃甘南州的玛曲、碌曲、四川的甘孜、阿坝,以及青海的玉树等地区。研究表明,青藏高原东缘高寒草原生态系统退化的典型特点为:以垂穗披碱草(Elymus nutans)、紫花针茅(Stipa purpurea)、早熟禾(Poa trivialis)等为代表的优良牧草种群逐渐衰退,而以黄帚橐吾(Ligularia virgaurea)、披针叶黄华(Thermopsis lanceolata)、狼毒(Stellera chamaejasme)等为代表的有毒植物种群则不断蔓延,局部地区甚至已经形成黄帚橐吾的单优势种群[1, 3],严重威胁到当地畜牧业的发展和生态系统的稳定。
高原鼠兔(Ochotona curzoniae)和高原鼢鼠(Myospalax baileyi)是青藏高原东缘高寒草原生态系统中两种最主要的小型哺乳动物。目前,围绕高原鼠兔和高原鼢鼠的研究主要集中在探讨二者如何适应青藏高原高海拔气候的生理机制,从分子生物学的角度阐明高原鼠兔适应高海拔、低含氧量生境的分子机理[4]。在生理学方面,研究发现高原鼢鼠通过增强肝脏中苹果酸天冬氨酸穿梭系统(malate-spartate shuttle system,MA)的穿梭能力提高了其在持续挖掘土壤活动中所需能量的供应,而高原鼠兔则通过增强肝脏糖异生的作用,为短暂、快速的剧烈运动提供能量来源,有效地将骨骼肌中无氧糖酵解产生的乳酸转化为糖原,从而减少对氧的依赖性[5]。此外,Yu 等通过遗传多样性分析探讨了高原鼠兔在青藏高原上的进化历史及其地理分布特征,提出高原鼠兔在青藏高原的分布现状是由于其辐射状分布和多样化的生境条件共同作用的结果[6],并揭示了其随海拔高度的变化而产生不同生态型之间的亲缘关系[7]。刘伟等分析了高原鼠兔挖掘形成的鼠丘上和鼠丘间植物群落的物种丰富度、物种组成和地上生物量的变化,以及鼠兔挖掘活动对植物群落的影响[8],提出利用生物防控的技术来控制高原鼠兔的危害[9],并采用数学模型对高原鼠兔的种群动态进行模拟,为开展生物防控提供依据[10]。
由于高原鼠兔和高原鼢鼠均为穴生动物,具有挖洞行为,必然产生大量的次生裸地,形成的空生态位可以为有毒植物的生长创造条件,形成草原退化、鼠害入侵、优良牧草减少、有毒植物蔓延的恶性后果。有毒植物的滋生又造成天然草场生产力下降,品质退化,容易导致家畜因误食而中毒,因而探究青藏高原东缘地区高寒草原有毒植物和高原鼠兔、高原鼢鼠之间的相互作用机制具有重要的生态学意义,但目前相关方面的研究尚少有报道。基于此,本研究拟以青藏高原东缘高寒草原生态系统为研究对象,探讨在自然生长的高寒草原生态系统中,有毒植物的种群蔓延与高原鼢鼠、高原鼠兔种群动态的互作关系,分析鼠类活动对草原生态系统的作用机制,进而探究其与有毒植物生长之间的相关性,为我国高寒草原的生态维持和草原生态系统的平衡和稳定提供理论依据。
1 材料与方法 1.1 研究材料高原鼢鼠(Myospalax baileyi),属啮齿目(Rodentia),鼹形鼠科(Spalacidae),鼢鼠属(Myospalax)。主要分布于海拔2800—4200 m、年积温1500 ℃左右的山地、农田、草甸草原等生境类型,为青藏高原特有的高度适应地下生活的鼠种之一[11]。高原鼢鼠主要依靠前肢、门牙或头部挖掘地下洞道系统,取食1年生或多年生非禾本科植物地下根、茎或芽[12]。高原鼢鼠不仅与家畜争夺优良牧草,降低草原载畜量,而且打洞造穴、挖掘草根、推出鼠丘,导致地表土裸露,植被盖度降低,引起草原退化、生产力降低等生态学问题。
高原鼠兔(Ochotona curzoniae),属兔形目(Lagomorpha),鼠兔科(Ochotonidae),鼠兔属(Ochotona)。 喜居于坡位较低,土壤疏松干燥,地形开阔,离水源较近,植被低矮的高山草甸和亚高山草甸,营洞穴生活。多以植物叶、茎、花及根、芽为食,而对粗老纤维化或含水量少的器官和组织采食较少。高原鼠兔喜食禾本科的紫花针茅(Stipa purpurea)、早熟禾(Poa sp.)及豆科的甘肃棘豆(Oxytropis kansuensis)、花苜蓿(Medicago ruthenica)等植物。
1.2 研究地区本研究选择甘肃省甘南藏族自治州玛曲县河曲马场自然生长的高山草原作为试验区(N 33°50′23″,E 102°08′48″,海拔3434 m)。该实验区为一地势平坦、植被分布较为均一、围栏封育2a的典型高寒草原生态系统,面积 >1000 hm2。植被类型以垂穗披碱草、紫花针茅、早熟禾、禾叶嵩草(Kobresia graminifolia)、鹅绒萎陵菜(Potentilla ansrina)、条叶银莲花(Anemone trullifolia)、莓叶萎陵菜(Potentilla fragarioides)等为优势物种。草地植被盖度70%—100%,主要伴生物种包括披针叶黄华、黄帚橐吾、蒲公英(Taraxacum officnala)、丝叶毛茛(Ranunculus tanguticus)、乳浆大戟(Euphorbia esula)、独一味(Ajugaovalifolia)、黄花棘豆(Oxytropis ochrocephala)、小花草玉梅(Anemone rivularis)、钝裂银莲花(Anemone obtusiloba)等。
1.3 有毒植物种类调查因为青藏高原高寒草原的植物生长期仅为每年的6月初至10月底,而6—7月份植物刚刚发芽生长,个体较小,区分植物种类难度较大。同时,到了10月份,高寒草原的植株已经进入果实成熟期,植株开始枯黄。因而8—9月是该地区高寒草原植被特征最为典型的季节,可以较好的代表有毒植物的群落特征。基于此,本研究选择2012年8月20日—8月30日在甘南州玛曲县河曲马场开展相关的野外实验。在实验区自然生长的高寒草原生态系统中采集所有植物种类的植株样品,对其进行鉴定,结合《中国有毒植物》[13]和《中国草地重要有毒植物》[14]等文献资料,统计实验区内所有有毒植物的名称和种类。
1.4 高原鼢鼠与有毒植物的相关性实验高原鼢鼠是一种营地下生活的穴居动物,很难直接统计出其活的鼠群数量,基于此,本研究采用土丘系数法来研究高原鼢鼠的种群数量。研究发现,高原鼢鼠具有独特的生活特性,其在特定空间范围内所挖掘的鼠丘数量和其成活个体数量之间存在显著的相关性[15, 16]。同时,高原鼢鼠具有保持所有生活洞口封闭性的特点,即,当鼠丘通道被打开之后,高原鼢鼠会很快堵住被打开的洞口(堵洞),因而本研究采用铁锨在第1天将样方内所有的鼠丘打开,第2天统计堵洞的数量。如果鼠丘被封堵即代表这些洞内有成活的高原鼢鼠,为有效鼠丘。若未被堵洞,则为无效鼠丘。因此,本研究采用统计样地内所有有效鼠丘数量的方法(即土丘系数法)来代表高原鼢鼠的种群数量。同时,为了避免本研究中任意两个不同样方之间的高原鼢鼠为同一个鼠群,设定任意两个样方之间的距离均>50 m。
试验期间,在高原鼢鼠实验地任意选择不同密度的鼠丘数量作为指标开展数据测定。每个实验样地的面积设定为10 m×10 m,统计样方内所有有效鼠丘的数量。共计测定25个重复。同时,为了将高原鼢鼠的密度和有毒植物的植被特征进行相关性研究,本实验在每个测定高原鼢鼠密度的10 m×10 m样方范围内,随机选择一个1 m×1 m的样方,记录其中所有有毒植物的种类和每个物种的植株数目,共计25个重复。
1.5 高原鼠兔与有毒植物的相关性实验高原鼠兔是一种在地下打洞、地面生活的穴居动物,同样很难直接统计其活的鼠群数量,基于此,本研究采用堵洞盗洞法来研究高原鼠兔的数量。研究发现高原鼠兔在特定空间范围内所挖掘的洞口数量与其种群数量存在显著的相关性[17]。为了有效的逃避天敌和方便出入,高原鼠兔具有保持所有洞口开放的特性。当其洞口被堵塞之后,高原鼠兔会很快移除障碍物,重新打开被堵塞的洞口,因而本研究采用野草在第1天将样方内所有的高原鼠兔洞口堵塞,第2天进行观察。如果洞口被打开即代表这些洞内有生活的高原鼠兔,为有效洞口。若未被打开,则为无效洞口。因而本研究采用统计有效洞口数量的方法(即堵洞盗洞法)代表高原鼠兔的种群数量。同时,为了避免本研究两个不同样方之间的高原鼠兔为同一鼠群,本研究设定任意两个样方之间的距离均 >50 m。每个实验样地的面积设定为10 m×10 m,统计每个样方内有效洞口的数量。共计测定25个重复。
同时,为了开展高原鼠兔密度和有毒植物植被特征之间的相关性研究,本实验在每次测定高原鼠兔密度的10 m×10 m样方范围内,随机选择一个1 m×1 m的样方,记录其中所有有毒植物的种类和每个物种的植株数目,共计25个重复。
1.6 数据统计所有原始数据在进行统计分析之前,均采用Statistica(v6.0)统计软件进行正态性检验,结果表明均符合正态性分布,因而没有进行数据转换。围绕本实验的结果,首先开展有毒植物的生物多样性指数、均匀度指数、丰富度指数的计算,采用马克平等[18]、Margalef指数等[19]的测定方法。
1)生物多样性指数,测定Simpson指数:
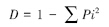
式中,Pi为种i 的个体数占群落中总个体数的比例。
2)均匀度指数,测定Pielou指数:
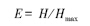
式中,H为实际观察的物种多样性指数,Hmax为最大物种多样性指数,H max = lnS,S为群落中的总物种数。
3)物种丰富度指数,测定Margalef指数:
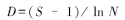
式中,S为群落中的总物种数,N为观察到的个体总数。
在此基础上,采用SPSS(v16.0)进行高原鼢鼠、高原鼠兔的种群数量与有毒植物的物种数、生物多样性指数、均匀度指数、丰富度指数的相关性分析和差异显著性分析。
2 结果 2.1 高寒草原有毒植物的种类实验结果表明(表 1),在青藏高原玛曲县河曲马场自然生长的高寒草原生态系统中,共计采集有毒植物27种,分属于菊科、豆科、毛茛科等11科,以菊科、毛茛科、龙胆科、豆科等为主。
|
科名
Family name | 中文名
Chinese name | 拉丁学名
Latin name | 植物类型
Plant type |
| 菊科 Asteraceae | 黄帚橐吾 | Ligularia virgaurea | 多年生草本植物 |
| 菊科Asteraceae | 长毛风毛菊 | Saussurea hieracioides | 多年生草本植物 |
| 菊科Asteraceae | 乳白香青 | Anaphalis lactea | 多年生草本植物 |
| 豆科Leguminosae | 黄花棘豆 | Oxytropis ochrocephala | 多年生草本植物 |
| 豆科Leguminosae | 披针叶黄华 | Thermopsis lanceolata | 多年生草本植物 |
| 大戟科Euphorbiaceae | 乳浆大戟 | Euphorbia esula | 多年生草本植物 |
| 毛茛科Ranunculaceae | 小花草玉梅 | Anemone rivularis | 多年生草本植物 |
| 毛茛科Ranunculaceae | 钝裂银莲花 | Anemone obtusiloba | 多年生草本植物 |
| 毛茛科Ranunculaceae | 匙叶银莲花 | Anemone trullifolia | 多年生草本植物 |
| 毛茛科Ranunculaceae | 丝叶毛茛 | Ranunculus tanguticus | 多年生草本植物 |
| 毛茛科Ranunculaceae | 高山唐松草 | Thalictrum alpinum | 多年生草本植物 |
| 毛茛科Ranunculaceae | 花葶驴蹄草 | Caltha scaposa | 多年生草本植物 |
| 毛茛科Ranunculaceae | 驴蹄草 | Caltha palustris | 多年生草本植物 |
| 毛茛科Ranunculaceae | 矮金莲花 | Trollius farrer | 多年生草本植物 |
| 毛茛科Ranunculaceae | 金莲花 | Trollius chinensis | 1年生或多年生草本植物 |
| 毛茛科Ranunculaceae | 翠雀 | Delphinium grandiflorum | 多年生草本植物 |
| 毛茛科Ranunculaceae | 水葫芦苗 | Halerpestes cymbalaria | 多年生草本植物 |
| 龙胆科Gentianaceae | 鳞叶龙胆 | Gentiana squarrosa | 1年生草本植物 |
| 龙胆科Gentianaceae | 湿生萹蕾 | Gentianopsis paludosa | 1年生草本植物 |
| 龙胆科Gentianaceae | 麻花艽 | Gentiana straminea | 多年生草本植物 |
| 龙胆科Gentianaceae | 秦艽 | Gentiana macrophylla | 多年生草本植物 |
| 茄科Solanaceae | 山莨菪 | Anisodus tanguticus | 多年生草本植物 |
| 瑞香科Thymelaeaceae | 狼毒 | Stellera chamaejasme | 多年生草本植物 |
| 玄参科Scrophulariaceae | 甘肃马先蒿 | Pedicularis kansuensis | 1年生或2年生草本植物 |
| 唇形科Lamiaceae | 圆叶筋骨草(独一味) | Ajuga ovalifolia | 多年生草本植物 |
| 木贼科Equisetaceae | 问荆 | Equisetum arvense | 多年生草本植物 |
| 鸢尾科Iridaceae | 鸢尾 | Iris tectorum | 多年生草本植物 |
围绕高寒草原有毒植物的特征指标与高原鼠兔、高原鼢鼠的相关性研究表明,无论是在高原鼠兔分布区,还是高原鼢鼠分布区,有毒植物的种类均为8—13种(图 1)。在高原鼢鼠、高原鼠兔与有毒植物物种数量的相关性方面,研究发现,高原鼠兔的有效洞口密度与有毒植物的种类之间存在负相关关系(P<0.05),而高原鼢鼠的数量变化对有毒植物的种类没有显著影响(P>0.05)(图 1)。

|
| 图1 青藏高原东缘高寒草原高原鼠兔、高原鼢鼠与有毒植物物种数量之间的相关性 Fig. 1 Correlation between the numbers of poisonous plants and Ochotona curzoniae,Myospalax baileyi in the East of Tibetan Plateau Alpine steppe ecosystem |
对青藏高原东缘高寒草原有毒植物的生物多样性指数分析表明,高原鼠兔和高原鼢鼠的种群密度和有毒植物的多样性指数之间存在显著差异(图 2)。在高原鼠兔方面,随着高原鼠兔密度的增加,有毒植物的多样性指数呈现出下降的趋势(P<0.05)。而在高原鼢鼠分布区,有毒植物的生物多样性指数不随高原鼢鼠的鼠丘数量变化而改变,表明高原鼢鼠的种群扩散与有毒植物的生物多样性之间没有显著的相关性(P>0.05)。

|
| 图2 青藏高原东缘高寒草原高原鼠兔、高原鼢鼠与有毒植物生物多样性之间的相关性 Fig. 2 Correlation between the biodiversity of poisonous plants and Ochotona curzoniae,Myospalax baileyi in the East of Tibetan Plateau Alpine steppe ecosystem |
在均匀度方面,由实验结果可知青藏高原高寒草原高原鼠兔、高原鼢鼠与有毒植物的均匀度指数之间均没有显著的相关性(P>0.05)(图 3)。随着高原鼠兔和高原鼢鼠密度的增加,有毒植物的均匀度均未发生显著改变(图 3)。

|
| 图3 青藏高原东缘高寒草原高原鼠兔、高原鼢鼠与有毒植物均匀度指数之间的相关性 Fig. 3 Correlation between the evenness index of poisonous plants and Ochotona curzoniae,Myospalax baileyi in the East of Tibetan Plateau Alpine steppe ecosystem |
在有毒植物的物种丰富度方面,本研究发现有毒植物的丰富度指数随着高原鼠兔密度的增长而降低(P<0.05);而高原鼢鼠的密度与有毒植物的丰富度指数之间没有显著性的相关关系(P>0.05)(图 4)。

|
| 图4 青藏高原东缘高寒草原高原鼠兔、高原鼢鼠与有毒植物物种丰富度指数之间的相关性 Fig. 4 Correlation between the Margalef richness index of poisonous plants and Ochotona curzoniae,Myospalax baileyi in the East of Tibetan Plateau Alpine steppe ecosystem |
高原鼠兔和高原鼢鼠作为青藏高原高寒草原中的两种重要小型哺乳动物,在草原生态系统的群落结构稳定性维持中发挥了重要作用,其种群特征必然对该地区植物群落的物种组成、密度、盖度、多样性指数等产生显著影响[20]。
在有毒植物种群特征与高原鼢鼠的相关性实验方面,前人研究表明,高原鼢鼠挖掘鼠丘的行为是草原生态系统土壤更新的一种非常重要的方法。高原鼢鼠在鼠洞挖掘、鼠丘营造的过程中,将深层土壤转移至地表,不仅增加了土壤通透性,加速水分渗透,增加土壤含水量和有机质含量[21, 22],而且加速了速效养分的释放,使土壤有效养分含量上升。与此同时,新土丘的形成也提供了大量的空生态位,由于目前青藏高原高寒草原中优良牧草被家畜过量采食,限制了优良牧草的生长,因而新土丘的形成恰好为有毒植物的建植提供了契机,导致有毒植物的扩散和蔓延[8, 23]。但与此同时,由于高原鼢鼠在地下采食多为1年生或多年生非禾本科植物的地下根、茎或芽,其中包括部分有毒植物的地下根、茎[12],因而高原鼢鼠的采食作用可显著降低直根类及根茎类植物的生活力,使须根类的禾本科植物(优良牧草)获得竞争优势。与此同时,由于洞道挖掘致使禾本科植物的部分根系受损[11]。因而,二者的作用可能相互抵消,导致地上有毒植物的物种多样性与多样性指数保持稳定,而未产生显著变化(图 1,图 2),表明高原鼢鼠对有毒植物的影响受到土壤环境、采食等多种因素的影响。张军等也开展了不同密度条件下高原鼢鼠对植被盖度、高度、地上生物量和植物多样性等指标的研究,发现植被高度和生物量易受到环境中不确定因素的干扰,而盖度和多样性则相对稳定[20],与本研究结果一致。
高原鼠兔是一种在地下打洞、地面生活的穴居动物。本研究结果发现,青藏高原高寒草原有毒植物的蔓延与高原鼠兔的密度之间存在显著的负相关关系。该结果与预期的草原退化、鼠害入侵、优良牧草减少、有毒植物蔓延存在差异,分析其可能的原因如下:(1)对高原鼠兔而言,青藏高原高寒草原植被是其栖息生境中最重要的组成部分之一,不仅是高原鼠兔食物的主要来源,而且也是其生境的重要保护因子[24]。同时,高原鼠兔通过挖掘鼠道、打开鼠洞、掘土等行为对地表植被产生影响。鼠道的挖掘可以把地下深层土壤转移到地表,不仅能够覆盖临近的植物,还可以形成空的生态位,为新生植株提供次生裸地,从而导致包括有毒植物在内的新生植株的生长和繁殖。但在鼠洞挖掘初期,由于次生裸地的形成也覆盖了部分原有的植被群落,导致植被盖度的下降,其中包括有毒植物和优良牧草,因而二者的作用效应可能相互抵消。(2)鼠道、鼠洞的形成也可以改变土壤理化性质和土壤种子库,导致很多新生植株的出现,这些新生植株可能来自于有毒植物的同一物种或者少数几个物种,最后表现为有毒植物植株数量增加,但其物种数和多样性指数却降低,因而高密度的鼠洞对植被的物种组成、草丛高度和生物量等产生不利影响。(3)高原鼠兔挖掘鼠道的行为可以在草原上形成大量的异质性斑块,鼠群密度越高,植被所受到的扰动也就越强烈,本实验结果即表现为随着高原鼠兔密度的增加,有毒植物的物种数、多样性指数均呈显著降低的趋势(图 1,图 2,图 4)。植被性状的变化在很大程度上反映了鼠群密度的变化,二者之间存在着显著的相关关系。研究发现,在高原鼠兔危害程度较轻的区域,原生植被(优良牧草)的重要值较高,杂类草的重要值较低,而危害程度较重的区域正好相反,表明高寒草原植被在受到高原鼠兔危害过程中会产生退化演替效应[25],这也是退化草原有毒植物蔓延的一个重要原因。
综上所述,本研究结果表明,青藏高原东缘高寒草原有毒植物的蔓延与高原鼠兔的密度之间存在显著的负相关关系,而与高原鼢鼠的密度相关性不显著。
| [1] | Jin L, Zhang G Q, Wang X J, Dou C Y, Chen M, Lin S S, Li Y Y. Arbuscular mycorrhiza regulate inter-specific competition between a poisonous plant, Ligularia virgaurea, and a co-existing grazing grass, Elymus nutans, in Tibetan Plateau Alpine meadow ecosystem. Symbiosis, 2011, 55(1): 29-38. |
| [2] | Wang M T, Lu N N, Zhao Z G. Effects of temperature and storage length on seed germination and the effects of light conditions on seedling establishment with respect to seed size in Ligularia virgaurea. Plant Species Biology, 2009, 24(2): 120-126. |
| [3] | Wang M T, Zhao Z G, Du G Z, He Y L. Effects of light on the growth and clonal reproduction of Ligularia virgaurea. Journal of Integrative Plant Biology, 2008, 50(8): 1015-1023. |
| [4] | Wang Z L, Zhang Y M. Predicted structural change in erythropoietin of plateau zokors-Adaptation to high altitude. Gene, 2012, 501(2): 206-212. |
| [5] | Zhu R J, Rao X F, Wei D B, Wang D W, Wei L, Sun S Z. Functional difference of malate-aspartate shuttle system in liver between plateau zokor (Myospalax baileyi) and plateau pika (Ochotona curzoniae). Acta Physiologica Sinica, 2012, 64(2): 177-186. |
| [6] | Yu F H, Li S P, Kilpatrick W C, McGuire P M, He K, Wei W H. Biogeographical study of plateau pikas Ochotona curzoniae (Lagomorpha, Ochotonidae). Zoological Science, 2012, 29(8): 518-526. |
| [7] | Wang Z L, Chen Y, Yang J, Chen W J, Zhang Y M, Zhao X Q. cDNA cloning and expression of erythropoietin in the plateau zokor (Myospalax baileyi) from the Qinghai-Tibet Plateau. Chinese Science Bulletin, 2012, 57(9): 997-1006. |
| [8] | Liu W, Li L, Yan H Y, Sun H Q, Zhang L. Effects of burrowing activity of plateau pika (Ochotona curzoniae) on plant species richness and aboveground biomass. Acta Theriologica Sinica, 2012, 32(3): 216-220. |
| [9] | Du Y, Cao Y F, Jing Z C, He H, Bian J H. Efficacies of coccidian parasites (Protozoa) in control of plateau pika (Ochotona curzoniae) and their effects on embryo development. Acta Theriologica Sinica, 2012, 32(3): 221-227. |
| [10] | Liu H W, Chen Y M, Zhou L, Jin Z. The effects of management on population dynamics of plateau pika. Mathematical and Computer Modelling, 2013, 57(3/4): 525-535. |
| [11] | Zhang Y M, Liu J K. Excavating effects of plateau zokor on the plant biomass and their response patterns in alpine meadow. Acta Theriologica Sinica, 2002, 22(4): 292-298. |
| [12] | Zhang Y M, Liu J K. Effects of plateau zokors (Myospalax fontanierii) on plant community and soil in an alpine meadow. Journal of Mammalogy, 2003, 84(2): 644-651. |
| [13] | Chen J S, Zheng S. The Poisonous Plant in China. Beijing: Science Press, 1987. |
| [14] | Shi Z C. Important Poisonous Plants of China Grassland. Beijing: China Agriculture Press, 1997. |
| [15] | Wang Q Y, Fan N C. Studies on the digging activities and exploration about the method of number estimation of plateau zokor. Acta Theriologica Sinica, 1987, 7(4): 283-290. |
| [16] | Yang S M, Wei W H, Yin B F, Fan N C, Zhou W Y. The predation risks of the plateau pika and plateau zokor and their survival strategies in the Alpine Meadow Ecosystem. Acta Ecologica Sinica, 2007, 27(12): 4972-4978. |
| [17] | Jiang X L. Relationship of population quantities of plateau pika with vegetation homogeneity. Acta Pratacultural Science, 1998, 7(1): 60-64. |
| [18] | Ma K P, Liu Y M. The methods for community biodiversity determination I: α index of biodiversity. Chinese Biodiversity, 1994, 2(4): 231-239. |
| [19] | Sun R Y, Li B, Zhuge Y, Shang Y C. General Ecology. Beijing: Higher Education Press, 1993. |
| [20] | Zhang J, Ge Q Z, Zhang W G, Jiang X L. Correlations between vegetation properties and fitness of plateau zoker habitat. Pratacultural Science, 2011, 28(5): 836-840. |
| [21] | Kerley G I H, Withford W G, Kay F R. Effects of pocket gophers on desert soils and vegetation. Journal of Arid Environments, 2004, 58(2): 155-166. |
| [22] | Li W J, Zhang Y M. Impacts of plateau pikas on soil organic matter and moisture content in alpine meadow. Acta Theriologica Sinica, 2006, 26(4): 331-337. |
| [23] | Karsten W, Karin N, Vroni R. Habitat engineering under dry conditions: The impact of pikas (Ochotona pallasi) on vegetation and site conditions in southern Mongolian steppes. Journal of Vegetation Science, 2007, 18(5): 665-674. |
| [24] | Qu J P, Li W J, Yang M, Ji W H, Zhang Y M. Life history of the plateau pika (Ochotona curzoniae) in alpine meadows of the Tibetan Plateau. Mammalian Biology, 2013, 78(1): 68-72. |
| [25] | Huang C D. Effect of plateau xokoron grassland vegetatiln in cold and high meadow. Pratacultural Science, 2004, 21(10): 58-62. |
| [5] | 朱瑞娟, 饶鑫峰, 魏登邦, 王多伟, 魏莲, 孙生祯. 高原鼢鼠和高原鼠兔肝脏苹果酸天冬氨酸穿梭系统的功能差异. 生理学报, 2012, 64(2): 177-186. |
| [8] | 刘伟, 李里, 严红宇, 孙海群, 张莉. 高原鼠兔挖掘活动对植物种的丰富度和地上生物量的影响. 兽类学报, 2012, 32(3): 216-220. |
| [9] | 杜寅, 曹伊凡, 景增春, 何慧, 边疆晖. 艾美尔球虫防治高原鼠兔实验及对其胚胎发育的影响. 兽类学报, 2012, 32(3): 221-227. |
| [11] | 张堰铭, 刘季科. 高原鼢鼠挖掘对植物生物量的效应及其反应格局. 兽类学报, 2002, 22(4): 292-298. |
| [13] | 陈冀胜, 郑硕. 中国有毒植物. 北京: 科学出版社, 1987. |
| [14] | 史志诚. 中国草地重要有毒植物. 北京: 中国农业出版社, 1997. |
| [15] | 王权业, 樊乃昌. 高原鼢鼠(Myospalax baileyi)的挖掘活动及其种群数量统计方法的探讨. 兽类学报, 1987, 7(4): 283-290. |
| [16] | 杨生妹, 魏万红, 殷宝法, 樊乃昌, 周文扬. 高寒草甸生态系统中高原鼠兔和高原鼢鼠的捕食风险及生存对策. 生态学报, 2007, 27(12): 4972-4978. |
| [17] | 江小雷. 植被均匀度与高原鼠兔种群数量相关性研究. 草业学报, 1998, 7(1): 60-64. |
| [18] | 马克平, 刘玉明. 生物群落多样性的测度方法I α多样性的测度方法(下). 生物多样性. 1994, 2(4): 231-239. |
| [19] | 孙儒泳, 李博, 诸葛阳, 尚玉昌. 普通生态学. 北京: 高等教育出版社, 1993. |
| [20] | 张军, 葛庆征, 张卫国, 江小雷. 植被性状与高原鼢鼠栖息地适合度的关系. 草业科学, 2011, 28(5): 836-840. |
| [22] | 李文靖, 张堰铭. 高原鼠兔对高寒草甸土壤有机质及湿度的作用. 兽类学报, 2006, 26(4): 331-337. |
| [25] | 黄成定. 高原鼢鼠危害对高寒草甸植被的影响. 草业科学, 2004, 21(10): 58-62. |