 2014, Vol. 34
2014, Vol. 34文章信息
- 葛晓改, 肖文发, 曾立雄, 黄志霖, 周本智
- GE Xiaogai, XIAO Wenfa, ZENG Lixiong, HUANG Zhilin, ZHOU Benzhi
- 三峡库区马尾松林土壤-凋落物层酶活性对凋落物分解的影响
- Effect of soil-litter layer enzyme activities on litter decomposition in Pinus massoniana plantation in Three Gorges Reservoir Area
- 生态学报, 2014, 34(9): 2228-2237
- Acta Ecologica Sinica, 2014, 34(9): 2228-2237
- http://dx.doi.org/10.5846/stxb201305261185
-
文章历史
- 收稿日期:2013-5-26
- 修订日期:2013-11-14






2. 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091
2. Key Laboratory of Forest Ecology and Environment, the State Forestry Administration; Research Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China
森林凋落物在促进森林生态系统正常的物质循环、养分平衡及维持生态系统功能中具有重要作用;凋落物分解是陆地生态系统土壤有机质、有机养分矿化和碳平衡形成的重要环节[1],在促进森林生态系统正常物质循环和养分平衡[2]、维持土壤肥力、保证植物再生长养分的可利用性中起着重要作用[3]。凋落物分解过程中胞外酶破坏了凋落物的结构组成并还原了有机C、N、P,因而凋落物分解过程是一个受制于土壤和凋落物中酶系统的复杂生物化学过程[4];凋落物的彻底降解是在凋落物和土壤中酶系统的综合作用下完成,酶活性的提高有利于凋落物和土壤有机物质的分解、养分释放,对于维持森林生态系统的物质循环和能量流动及提高森林土壤肥力具有重要意义[5]。
森林凋落物的分解过程是基于分解者细胞水平的酶解过程,有关凋落物分解过程中土壤和凋落物的酶活性动态已开展了研究,如张瑞清等[6]在西双版纳对凋落物叶分解过程中酶活性动态研究表明,凋落物分解速率与蔗糖酶活性呈负相关,与多酚氧化酶活性呈正相关;张鹏等[7]研究表明凋落物分解与土壤酶活性直接相关,土壤酶活性与温度之间都有一定的相关性,最高值均出现在温度较高的7月份;郑洪元等[8]研究表明水解酶系(脲酶、蔗糖酶)主要在分解第一阶段起作用,土壤中可溶性物质(氨态氮、酮酸等)有所增加,随着分解的进行,土壤水解酶系和氧化还原酶系(多酚氧化酶)共同完成,土壤中可溶性物质明显下降,形成的腐殖质开始积累。这些研究均表明凋落物-土壤酶活性在森林凋落物中有着重要作用,森林生态系统凋落物分解与酶系统的相互作用机制将成为新的发展趋势[9],对森林生态系统中物质循环速度和能量流动具有重要作用。目前,森林凋落物分解过程中酶活性的研究多集中于酶活性的测定方法及其影响因素方面[9],然而有关土壤和凋落物层酶对分解的影响研究多数是基于相关分析,凋落物分解过程中凋落物层酶活性和土壤酶活性数量关系及对分解的影响研究报道较少。
马尾松(Pinus massoniana)是我国亚热带地区主要的针叶树种,自然分布面积达到220万hm2,主要分布在南方15个省(区)[10],因其适应性强、耐干旱与瘠薄,广泛用于荒山绿化,是亚热带地区主要的造林树种,也是三峡库区典型造林树种之一。本文以三峡库区30年生马尾松人工林为研究对象,对其凋落物分解动态、凋落物层酶活性、土壤酶活性及其关系进行研究,为凋落物分解过程中土壤肥力状况提供基础资料,为森林生态系统C、N、P等养分的转化提供一定的指示作用,为马尾松人工林土壤质量评价和亚热带地区森林可持续经营提供参考依据,对深入揭示森林生态系统养分循环研究具有重要意义。
1 研究区域概况和研究方法 1.1 研究区概况研究区位于湖北省秭归县(30°38′14″—31°11′31″N,110°00′14″—111°18′41″E),海拔40—2057 m,该地区属于亚热带大陆性季风气候,年均气温16.9 ℃,极端最高气温44 ℃,极端最低气温-2.5 ℃,年降水量1000—1250 mm,降水集中在4—9月,空气湿度达60%—80%,无霜期300—340 d。 秭归县海拔500 m以下以茶园、板栗、柑橘和农田为主,海拔500 m以上主要为灌木、马尾松、杉木(Cunninghamia lanceolata)和松栎混交林等。
本研究以30年生马尾松林(面积为1 hm2)为研究对象,林下灌木多为油茶(Camellia oleifera)、檵木(Loropetalum chinense)和白背叶(Mallotus apeltus)等,草本多为稗草(Echinochloa crusgalli)、腹水草(Veronicastrum axillare)、菝葜(Smilax ocreata)、蜈蚣草(Eremochloa ciliaris)等,样地基本概况见表 1。
|
海拔
Elevation /m | 胸径
DBH /cm | 坡度
Slope /(°) | 坡向
Exposure | 郁闭度
Canopy density | 土壤厚度
Soil depth /cm | 凋落物现存量
Litter standing crop / (t/hm2) | 凋落物层厚度
Litter layer depth /cm |
| Ss: 半阴坡Semi-shady | |||||||
| 357 | 25.3 | 25° | Ss | 0.95 | 60—80 | 9.26±2.97 | 5.77±0.85 |
凋落物分解采用分解袋法。在30年生马尾松林中设置3个20 m×30 m的样方。于2010年5—6 月在林地样方外收集新近自然凋落、上层未分解的凋落物叶,将收集的凋落物叶充分混合,风干后称取凋落物叶15 g,直接装入尼龙分解袋中。尼龙网袋大小为20 cm×20 cm,孔径1 mm。于2010年7月底将凋落物袋随机放置在3个样方中,除去地表凋落物层,使凋落物叶分解袋贴近地面,让凋落物自然分解;放置凋落物分解袋后每月底进行新凋落物清理;3个样方中共放置90袋。
自放置日期起,每3个月每样方随机取回5袋凋落物;取回凋落物分解袋后,清除侵入的根系、泥沙,3袋烘至恒重,测干重并计算干重剩余率和分解速率。另外2袋保留新鲜样品,剪碎后过2 mm筛,于4 ℃冰箱内保存,用于凋落物层酶活性测定。同时,取3种林分凋落物叶各15 g,重复3次,在70 ℃下烘干至恒重,算出风干与烘干重转换系数。
1.2.2 土壤采样于2010年11月(分解90 d)、2011年6月(270 d)、9月初(360 d)(与凋落物袋回收同步)分别在30年生马尾松林样方中用直径5 cm的土钻取凋落物分解袋下面的土壤,分3层(0—5 cm,5—10 cm,10—20 cm)约0.5 kg,同一样方的同层土壤充分混合后作为一个样,每土样重复3次。带回实验室低温保存(4 ℃),样品去掉石砾、动植物残体、根系后混匀,过2 mm筛,装入保鲜袋4 ℃保存,并尽快进行土壤酶活性测定。
1.2.3 酶活性测定凋落物层酶活性和土壤酶活性参照关松荫[11]的方法分析。蔗糖酶活性测定采用3,5-二硝基水杨酸比色法,酶活性以24 h后1 g土壤生成的葡萄糖毫克数表示;纤维素酶活性测定采用硝基水杨酸比色法,酶活性以72 h后10 g土壤生成的葡萄糖毫克数表示;脲酶活性测定采用苯酚钠-次氯酸钠比色法,酶活性以24 h后1 g土壤中NH+4-N的毫克数表示;过氧化物酶活性测定采用容量法,酶活性以2 h后1 g土壤消耗0.005 mol/L I2标准液的毫升数表示;多酚氧化酶测定采用比色法,酶活性以2 h后1 g土壤中紫色没食子素的毫克数表示。
1.3 数据处理凋落物叶分解干重剩余率计算公式采用:

式中,R为凋落物叶分解干重剩余率(%),k为凋落物风干样品与烘干样品转换系数,Mt为凋落物t时间烘干样品重量,M0为凋落物起始风干样品重量。
凋落物叶分解速率计算公式采用:
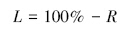
式中,L为凋落物叶分解速率(%),R为凋落物叶分解干重剩余率(%)。
干重剩余率用Olson[12]负指数衰减模型进行拟合:
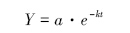
式中,Y为干质量年残留率(%),a为拟合参数,k为年分解系数(kg kg-1 a-1),t为分解时间(a)。凋落物分解半衰期(50%分解)计算式,t0.05=ln0.5/(-k);完全分解时间(95%分解)计算式,t0.95=ln0.05/(-k)。
利用SigmaPlot 11.0和SPSS 16.0软件对数据进行统计分析和作图。凋落物层酶活性和土壤层酶活性显著性检验采用单因子方差分析(ANOVA)和最小显著差异法(LSD),显著性水平设定为α=0.05。图表中数据均用平均值±标准差表示;凋落物分解速率与凋落物层、土壤层酶活性关系用SPSS 16.0 软件的回归分析。
2 结果与分析 2.1 马尾松林凋落物叶分解动态马尾松林凋落物叶经过540 d的分解干重剩余率是59.80%(表 2)。凋落物叶分解速率在0—180 d相对较快,在分解的180—270 d相对较慢(即2011年2—5月反常干旱,几乎没有降雨);凋落物叶分解速率在夏季分解最高(即分解的0—90 d、270—360 d)。马尾松林年凋落物叶分解常数是0.33,除分解的第6—9月外,实测的凋落物叶分解速率比预测的高;凋落物叶分解50%、95%的时间分别是2.11 a、9.15 a。
|
分解动态
Decomposition dynamic | 取样时间 Sampling time | |||||
| 90 d | 180 d | 270 d | 360 d | 450 d | 540 d | |
| 凋落物叶分解样品的90 d、180 d、270 d、360 d、450 d 、540 d对应于2010年11月、2011年3月、2011年6月、2011年9月、2011年11月、2012年1月 | ||||||
| 干重剩余率 Dry mass remaining/% | 91.68 | 82.00 | 83.66 | 73.89 | 63.32 | 59.80 |
| 标准差 Standard deviation/% | 2.60 | 3.35 | 5.01 | 4.43 | 5.44 | 1.60 |
| 变异系数 Coefficient of variation/% | 2.84 | 4.08 | 6.00 | 6.00 | 8.58 | 2.84 |
马尾松林凋落物层酶活性季节动态明显(图 1)。凋落物层氧化还原酶活性变化相似,多酚氧化酶活性和过氧化物酶活性均是11月最低,3月份最高,3月酶活性分别是11月酶活性的4.60倍、6.07倍;多酚氧化酶活性则是11月<6月<9月<3月;过氧化物酶活性则是11月<9月<6月<3月。凋落物层水解酶变化不一致,脲酶活性是9月最高,6月最低,且差异显著(P<0.05);纤维素酶活性则是6月<9月<3月<11月,最高酶活性是最低时的1.52倍,差异显著(P<0.05);蔗糖酶活性则是9月<6月<11月<3月,3月蔗糖酶活性是9月的2.06倍。

|
| 图1 尾松林凋落物层酶活性季节动态 Fig. 1 Seasonal dynamics of litter layer enzymes activities on P. massoniana forest |
马尾松林土壤酶活性季节动态明显(图 2)。30年生马尾松林土壤氧化还原酶均是11月最低,多酚氧化酶活性9月较高,过氧化物酶活性则是6月较高,且最低酶活性与最高酶活性差异显著(P<0.05)。马尾松林土壤水解酶中土壤脲酶活性季节变化不明显(P>0.05),纤维素酶活性和蔗糖酶活性季节变化明显,蔗糖酶活性是9月<6月<11月,纤维素酶活性是11月<9月<6月。

|
| 图2 马尾松林土壤层 (0—20 cm) 酶活性特征 Fig. 2 Enzyme activities characteristic of soil layer (0—20 cm) on P. massoniana forest 不同大写字母表示不同月份同种酶活性间差异显著,小写字母表示不同土层间差异显著(P<0.05) |
马尾松林土壤酶活性在垂直方向上随着土层深度的增加,除过氧化物酶外,其它酶活性均逐渐降低(图 2)。多酚氧化酶和过氧化物酶活性11月和6月垂直方向上差异不显著,9月多酚氧化酶活性随土层深度的增加明显降低,过氧化物酶活性9月随土层深度的增加先增加后降低。马尾松土壤脲酶活性11月和6月在垂直方向上差异显著(P<0.05);纤维素酶活性和蔗糖酶活性各个月份在垂直方向上均差异显著。
2.4 凋落物层酶和土壤层酶活性对分解的影响马尾松林凋落物层酶活性与土壤层酶活性差异较大(图 3)。凋落物层多酚氧化酶活性11月、6月、9月分别是0—5 cm土壤层的1.76倍、1.46倍、2.07倍,过氧化物酶活性分别是0.40倍、0.64倍、0.56倍。水解酶活性差异较氧化还原酶活性差异大,凋落物层脲酶活性、纤维素酶活性和蔗糖酶活性11月、6月、9月分别是0—5 cm土壤层的6.33倍、3.24倍、10.29倍;68.14倍、16.16倍、24.81倍;25.07倍、31.88倍、29.20倍。

|
| 图3 马尾松林凋落物叶分解速率与凋落物层酶活性、土壤层酶活性关系 Fig. 3 The relationships between litter leaf decomposition rate and litter layer enzyme activities,soil layer enzyme activities on P. massoniana forest |
马尾松林凋落物叶分解速率与凋落物层和土壤层酶活性关系紧密(表 3)。马尾松林凋落物分解速率与凋落物层和土壤层中氧化还原酶活性关系一致,均呈极显著的“S”形函数关系。凋落物分解速率与凋落物层水解酶酶活性关系一致,且均呈极显著的二次函数关系(P<0.01),与土壤层水解酶活性呈极显著线性关系(P<0.01)。
|
酶活性
Enzyme activities | 来源
Source | 回归方程
Regression equation | R2 | P | |
| 氧化还原酶活性 | 多酚氧化酶活性 | 凋落物层 Litter layer | y=e(3.362-7.253)/x | 0.68 | 0.000 |
| Oxidoreductase activity | Polyphenoloxidase activity | 土壤层 Soil layer | y=e(3.175-2.358)/x | 0.25 | 0.000 |
| 过氧化物酶活性 | 凋落物层 Litter layer | y=e(3.102-3.887)/x | 0.46 | 0.000 | |
| Peroxidase activity | 土壤层 Soil layer | y=e(3.516-17.32)/x | 0.37 | 0.000 | |
| 水解酶活性 | 脲酶活性Urease activity | 凋落物层 Litter layer | y=30.517-21.337x+5.682x2 | 0.70 | 0.000 |
| Hydrolase activity | 土壤层 Soil layer | y=3.742+44.823x | 0.19 | 0.001 | |
| 纤维素酶活性 | 凋落物层 Litter layer | y=-79.52+5.667x-0.08x2 | 0.43 | 0.003 | |
| Cellulase activity | 土壤层 Soil layer | y=8.752+7.642x | 0.22 | 0.000 | |
| 蔗糖酶活性 | 凋落物层 Litter layer | y=76.18-0.125x+5.975×10-2x2 | 0.36 | 0.010 | |
| Sucrase activity | 土壤层 Soil layer | y=25.924-0.396x | 0.28 | 0.000 |
凋落物分解是森林生态系统养分循环中至关重要的过程,凋落物分解与酶活性直接相关,森林凋落物分解过程中酶活性是一个动态过程,不同凋落物层酶活性季节变化明显[7]。Fioretto等[13]在地中海对灰白岩蔷薇(Cistus incanus)和香桃木(Myrtus communis)凋落物叶分解过程中酶动态研究表明:两种凋落物叶酶活性均呈现明显的季节动态,最低值均出现在夏季。Kshattriya等[14]研究表明纤维素等水解酶具有明显的季节动态,蔗糖酶活性在分解初期较高,而纤维素酶活性等随着分解活性增加。本研究中过氧化物酶活性(多酚氧化酶和过氧化物酶活性)和水解酶活性(纤维素酶和蔗糖酶活性)均是11月份最低,3月份最高,这可能因为生长季初期(3月)植物根系生长促进酶合成[15],微生物开始迅速繁殖,与细菌和真菌的数量有关的纤维素酶活性等水解酶活性明显增加;这与Di Nardo等[16]研究结果一致,即过氧化物酶活性在分解第1年较低,在分解第2年的10—1月增加,酶活性最高在1月份;而邓仁菊等[17]研究则表明凋落物层蔗糖酶活性、脲酶活性、脱氢酶活性均是秋季最高(8—10月),因为8—10月的气温更有利于凋落物中微生物群落的生长。不同地区相同酶活性的季节动态不同,可能因为森林凋落物分解过程中酶活性受气候因素(温度和湿度)、土壤化学性质和呼吸活动等多种因素的影响[9],同时,不同的酶活性的产量受植被类型和凋落物的影响也较大,不同的植被类型林下5 cm土壤温度差异较大[14]。
凋落物层水解酶活性和氧化还原酶活性对分解的影响不同。郑洪元等[8]研究表明水解酶系在分解第一阶段起作用,有机残体分解的第二阶段由水解酶系和氧化还原酶系共同完成。本研究中马尾松林凋落物分解速率与凋落物层水解酶活性均呈极显著的二次函数关系(P<0.01),因为在分解早期阶段凋落物纤维素等可溶性物质先分解[13],水解酶活性通常先增加后降低,尤其是凋落物分解30%—40%时,水解酶活性保持相对较高[13];季晓燕等[18]研究表明在一定范围内,凋落物分解酶活性随凋落物厚度的增加而增加,随分解时间的延长,脲酶、纤维素酶、蔗糖酶活性均有下降趋势。张瑞清等[6]对西双版纳热带雨林凋落叶分解过程中酶活性动态研究表明,蔗糖酶等水解酶活性在凋落物分解初级阶段发挥重要作用,参与易分解成分的转化和分解;多酚氧化酶等氧化还原酶活性在分解后期迅速上升,对凋落叶中、后期木质素的分解起到关键作用,这与本研究中凋落物叶分解速率与凋落物层氧化还原酶活性均呈极显著的“S”形函数关系(P<0.01)一致。Waring[19]研究表明胞外酶是凋落物叶分解的体现者,凋落物层纤维素酶活性、葡萄糖酶活性等与凋落物叶394 d分解速率呈正相关。因此,凋落物层水解酶活性主要在初期有助于凋落物中可溶性物质水解,随着分解时间的进行,氧化还原酶活性参与复杂有机化合物的分解;此外,凋落物分解速率与酶活性关系不同主要由凋落物层酶活性自身结构决定[20]。
3.2 土壤酶活性对凋落物分解的影响土壤酶是土壤生物化学过程的积极参与者,研究林分土壤酶活性的季节动态和差异对评价土壤和维持生态系统养分循环有重要意义[5]。陶宝先等[21]对苏南丘陵地区森林土壤酶活性季节变化研究表明,蔗糖酶活性、脲酶活性、过氧化物酶活性季节变化明显,夏、秋季酶活性相对较高,冬季酶活性均最低,高温因提高土壤酶活性而加速凋落物分解,这与本研究中6月份过氧化物酶活性、纤维素酶活性比11月份高一致,Fioretto等[13]研究也表明夏季0—5 cm土壤温度对酶活性的影响比冬季大,说明过氧化物酶活性、纤维素酶活性对温度较敏感,尤其在水分适宜条件下土壤温度-水分的协同作用促进凋落物分解的温度敏感性[22];支持熊浩仲等[23]在川西亚高山对冷杉林(Betula platyphylla)和白桦林(Abies faxoniana)土壤酶活性季节动态研究结论:即冷杉林表层过氧化氢酶活性7月最高,脲酶、多酚氧化酶和蔗糖酶活性均是10月最高;白桦林表层土壤蔗糖酶活性7月最高,多酚氧化酶10月最高,脲酶和过氧化氢酶活性4月最高;因为温度能提高土壤酶活性,多酚氧化酶明显比其它土壤酶的温度敏感性高,多酚氧化酶活性的Q10明显比纤维素酶、蔗糖酶、脲酶高[24];说明不同土壤酶活性对温度的敏感程度不同,且酶活性的温度敏感性由酶内部构型和结构决定[25]。
土壤层水解酶活性和氧化还原酶活性对分解的影响不同。当某一类酶活性高时,可以反映出某一特定微生物类群具有对凋落物中特定化学成分的高利用性。本研究中马尾松林凋落物叶分解速率与土壤层中氧化还原酶活性和水解酶活性呈不同的函数关系(P<0.01),可能因为土壤水解酶活性与氧化还原酶活性的主导影响因素不同,水解酶类的活性主要受到土壤水热动态的控制,水解酶活性的高低直接影响土壤中养分的供给,通常表现出先增加后降低的趋势;而土壤多酚氧化酶活性在春季最高,加速木质素降解、腐殖化、碳矿化、溶解性有机碳的输出等,多酚氧化酶活性高限制土壤有机碳的积累,酶活性低则会促进土壤有机碳的积累[26];而过氧化物酶活性同水解酶一样,酶活性在夏季最高、春季最小,主要加速土壤有机物的分解和腐殖质的合成。总之,氧化还原酶活性动态变化较为复杂,受土壤pH值、养分含量、土层分布等多种因素影响[4]。
3.3 凋落物层酶和土壤层酶活性的影响因素森林凋落物分解过程中的酶活性动态对指示凋落物分解过程具有重要意义,凋落物-土壤层酶活性动态是森林凋落物分解过程的重要指标[5]。本研究中马尾松林凋落物层酶活性与土壤层酶活性差异较大,且水解酶活性差异较氧化还原酶活性差异大,可能因为凋落物分解过程中凋落物层酶活性比土壤酶活性对凋落物分解质量动态变化敏感[4]。Keiblinger等[27]研究表明纤维素酶活性、过氧化物酶活性和多酚氧化物酶活性与凋落物C/N比呈负相关,与凋落物P含量和凋落物C/P比呈正相关;季晓燕等[18]开展凋落物叶厚度对其分解速率及酶活性的影响研究表明,水解酶活性随凋落物层厚度的增加而增加,因为凋落物叶较厚时,微生物可利用的物质能源越丰富,微生物的数量和种类相对较高;因此,凋落物质量是凋落物分解中酶活性的重要决定因素[28],微生物产生酶的数量由凋落物质量决定,微生物利用其不同的易溶和顽固复合物维持新陈代谢[14]。胡亚林等[29]用不同质量的混合凋落物分解对土壤酶活性影响研究表明,混合凋落物下土壤脲酶、蔗糖酶、脱氢酶活性升高,土壤多酚氧化酶活性下降;其主要原因是凋落物质量的不同导致土壤微生物量、区系组成以及代谢过程改变,致使主要由土壤微生物产生的土壤酶的数量和活性发生变化。
森林凋落物分解过程中酶活性除受凋落物质量的影响外,还受生物因素、非生物因素的影响,微生物群落是影响酶活性的主要生物因素,土壤有机质、温度和湿度等是影响酶活性的重要非生物因素[9]。Fioretto等[30]在地中海对灰白蔷薇(Cistus incanus)凋落物叶分解和酶活性研究表明,凋落物层水分含量明显与纤维素酶和木聚糖酶活性呈正相关;Wallenstein等[22]研究表明北美红栎(Quercus rubra) 和西黄松(Pinus ponderosa Douglas ex Lawson)凋落物叶培养14 d时,25 ℃时的凋落物层酶活性比35 ℃时高2倍。而土壤酶活性的影响因素较复杂,即土壤湿度、温度、pH值、可利用养分有效性均能影响土壤酶活性:徐振锋等[31]研究表明模拟增温增加了土壤蔗糖酶、脲酶、过氧化氢酶和多酚氧化酶活性,土壤酶活性季节动态与土壤温度有着较大相关性,而与土壤水分季节变化关系不明显;Wallenstein等[25]研究表明不同的酶活性对土壤养分的敏感性不同,夏季酶活性可能易受N含量的限制;葛晓改等[32]研究表明土壤养分对土壤酶活性影响显著,土壤主要养分含量高,蔗糖酶活性较高,过氧化物酶活性相对较低;杨万勤和王开运[5]研究表明土壤酶活性与土壤微生物数量、微生物多样性、微生物生物量及土壤动物数量等呈显著相关。凋落物层和土壤层酶活性的差异性不仅与凋落物分解阶段有关,最重要的是对共同影响因素(凋落物质量、温度、水分、土壤养分等)的敏感性不同,同时凋落物质量均能引起土壤酶活性的变化,酶活性的改变反过来又影响凋落物的分解[29];凋落物-土壤层酶相互作用,共同影响森林生态系统的物质循环和养分循环过程。
总之,马尾松林凋落物-土壤层过氧化物酶活性季节变化显著,凋落物分解速率均与土壤和凋落物层氧化还原酶活性呈极显著“S”形曲线,与凋落物层水解酶活性呈二次函数关系,与土壤层水解酶均呈极显著的线性关系;可能因为水解酶在凋落物分解初级阶段发挥重要作用,参与易分解成分的转化和分解;氧化还原酶类在分解后期迅速上升,对凋落叶中、后期木质素的分解起到关键作用。凋落物-土壤层酶活性的差异主要由凋落物分解阶段和共同影响因素的敏感性不同引起。
| [1] | Austin A T, Ballaré C L. Dual role of lignin in plant litter decomposition in terrestrial ecosystems. Proceeding of the National Academy of Sciences of the United States of America, 2010, 107(10): 4618-4622. |
| [2] | Song X Z, Jiang H, Zhang H L, Yu S Q, Zhou G M, Ma Y D, Chang S X. A review on the effects of global environment change on litter decomposition. Acta Ecologica Sinica, 2008, 28(9): 4414-4423. |
| [3] | Pan K W, He J, Wu N. Effect of forest litter on microenvironment conditions of forestland. Chinese Journal of Applied Ecology, 2004, 15(1): 153-158. |
| [4] | Kong A H. The Soil and Litters Enzyme Activity in Pinus tabulaeformis and Quercus variabis Forest at Lower Mountain Area Beijing . Beijing: Beijing Forestry University, 2012: 57-57. |
| [5] | Yang W Q, Wang K Y. Advances in forest soil enzymology. Scientia Silvae Sinicae, 2004, 40(2): 152-159. |
| [6] | Zhang R Q, Sun Z J, Wang C, Yuan T Y. Ecological process of leaf litter decomposition in tropical rainforest in Xishuangbanna, SW China Ⅲ. Enzyme dynamics. Journal of Plant Ecology, 2008, 32(3): 622-631. |
| [7] | Zhang P, Tian X J, He X B, Song F Q, Ren L L. Enzyme activities in litter, fragmentation and humus layers of subtropical forests. Ecology and Environment, 2007, 16(3): 1024-1029. |
| [8] | Zheng H Y, Zhang D S, Zhou W X, Zhang X W. Decomposition on forest organic detritus and soil enzyme activities. Chinese Journal of Soil Science, 1983, 14(1): 36-39. |
| [9] | Zhang D L, Mao Z J, Zhang L, Zhu S Y. Advances of enzyme activities in the process of litter decomposition. Scientia Silvae Sinicae, 2006, 42(1): 105-109. |
| [10] | Zhou Z X. Masson pine in China. Beijing: China Forestry press, 2000. |
| [11] | Guan S Y. Soil Enzyme and Its Research Methods. Beijing: China Agriculture Press, 1986. |
| [12] | Olson J S. Energy storage and the balance of producers and decomposers in ecological systems. Ecology, 1963, 44(2): 322-331. |
| [13] | Fioretto A, Papa S, Curcio E, Sorrentino G, Fuggi A. Enzyme dynamics on decomposing leaf litter of Cistus incanus and Myrtus communis in a Mediterranean ecosystem. Soil Biology and Biochemistry, 2000, 32(13): 1847-1855. |
| [14] | Kshattriya S, Sharma G D, Mishra R R. Enzyme activities related to litter decomposition in forests of different age and altitude in North East India. Soil Biology and Biochemistry, 1992, 24(3): 265-270. |
| [15] | Tierney G L, Fahey T J, Groffman P M, Hardy J P, Fitzhugh R D, Charles C T. Driscoll. Soil freezing alters fine root dynamics in a northern hardwood forest. Biogeochemistry, 2001, 56(2): 175-190. |
| [16] | Di Nardo C, Cinquegrana A, Papa S, Fuggi A, Fioretto A. Laccase and peroxidase isoenzymes during leaf litter decomposition of Quercus ilex in a Mediterranean ecosystem. Soil Biology and Biochemistry, 2004, 36(10): 1539-1544. |
| [17] | Deng R J, Yang W Q, Wu F Z. Effects of seasonal freeze-thaw on the enzyme activities in Abies faxoniana and Betula platyphylla litters. China Journal of Applied Ecology, 2009, 20(5): 1026-1031. |
| [18] | Ji X Y, Jiang H, Hong J H, Ma Y D. Effects of litter thickness on leaf litter decomposition and enzyme activity of three trees in the subtropical forests. Acta Ecologica Sinica, 2013, 33(6): 1731-1739. |
| [19] | Waring B G. Exploring relationships between enzyme activities and leaf litter decomposition in a wet tropical forest. Soil Biology and Biochemistry, 2013, 64(9): 89-95. |
| [20] | Dilly O, Munch J C. Microbial biomass content, basal respiration and enzyme activities during the course of decomposition of leaf litter in a black alder (Alnus Glutinosa L. Gaertn.) forest. Soil Biology and Biochemistry, 1996, 28(8): 1073-1081. |
| [21] | Tao B X, Zhang J C, Yu Y C, Cong R L. Season variations of forest soil enzyme activities in the hilly region of southern Jiangsu province. Ecology and Environmental Sciences, 2010, 19(10): 2349-2354. |
| [22] | Wallenstein M D, Haddix M L, Lee D D, Conant R T, Paul E A. A litter-slurry technique elucidates the key role of enzyme production and microbial dynamics in temperature sensitivity of organic matter decomposition. Soil Biology and Biochemistry, 2012, 47(1): 18-26. |
| [23] | Xiong H Z, Wang K Y, Yang W Q. Seasonal variations of soil enzyme activities in fir and birch forests in subalpine area of western Sichuan. Chinese Journal Applied and Environmental Biology, 2004, 10(4): 416-420. |
| [24] | Wang C Y, Han G M, Jia Y, Feng X G, Tian X J. Insight into the temperature sensitivity of forest litter decomposition and soil enzymes in subtropical forest in China. Journal of Plant Ecology, 2011, 5(3): 279-286. |
| [25] | Wallenstein M D, McMahon S K, Schimel J P. Seasonal variation in enzyme activities and temperature sensitivities in Arctic tundra soils. Global Change Biology, 2009, 15(7): 1631-1639. |
| [26] | Hu S, Zhang Y, Shi R J, Han S Q, Li H, Xu H. Temporal variations of soil microbial biomass and enzyme activities during the secondary succession of primary broadleaved-Pinuskoraiensis forest in Changbai Mountains of Northeast China. Chinese Journal of Applied Ecology, 2013, 24(2): 366-372. |
| [27] | Keiblinger K M, Schneider T, Roschitzkl B, Schmid E, Eberl L, Hmmerle I, Leitner S, Richter A, Wanek W, Riedel K, Zechmeister-Boltenstern S. Effects of stoichiometry and temperature perturbation on beech litter decomposition, enzyme activities and protein expression. Biogeosciences Discuss, 2011, 8(6): 11827-11861. |
| [28] | Kourtev P S, Ehrenfeld J G, Huang W Z. Enzyme activities during litter decomposition of two exotic and two native plant species in hardwood forests of New Jersey. Soil Biology and Biochemistry, 2002, 34(9): 1207-1218. |
| [29] | Hu Y L, Wang S L, Huang Y, Yu X J. Effects of litter chemistry on soil biological property and enzymatic activity. Acta Ecologica Sinica, 2005, 25(10): 2662-2668. |
| [30] | Fioretto A, Papa S, Sorrentino G, Fuggi A. Decomposition of Cistus incanus leaf litter in a Mediterranean maquis ecosystem: mass loss, microbial enzyme activities and nutrient changes. Soil Biology and Biochemistry, 2001, 33(3): 311-321. |
| [31] | Xu Z F, Tang Z, Wan C, Xiong P, Cao G, Liu Q. Effects of simulated warming on soil enzyme activities in two subalpine coniferous forests in west Sichuan. Chinese Journal of Applied Ecology, 2010, 21(11): 2727-2733. |
| [32] | Ge X G, Xiao W F, Zeng L X, Huang Z L, Huang L L, Tan B W. Relationship between soil nutrient contents and soil enzyme activities in Pinus massoniana stands with different ages in Three Gorges Reservoir Area. Chinese Journal of Applied Ecology, 2012, 23(2): 445-451. |
| [2] | 宋新章, 江洪, 张慧玲, 余树全, 周国模, 马元丹, Chang S X. 全球环境变化对森林凋落物分解的影响. 生态学报, 2008, 28(9): 4414-4423. |
| [3] | 潘开文, 何静, 吴宁. 森林凋落物对林地微生境的影响. 应用生态学报, 2004, 15(1): 153-158. |
| [4] | 孔爱辉. 北京低山区栓皮栎和油松林土壤与凋落物酶活性的研究[D]. 北京: 北京林业大学, 2012: 57-57. |
| [5] | 杨万勤, 王开运. 森林土壤酶的研究进展. 林业科学, 2004, 40(2): 152-159. |
| [6] | 张瑞清, 孙振钧, 王冲, 袁堂玉. 西双版纳热带雨林凋落物叶分解的生态过程 Ⅲ. 酶活性动态. 植物生态学报, 2008, 32(3): 622-631. |
| [7] | 张鹏, 田兴军, 何兴兵, 宋富强, 任利利. 亚热带森林凋落物层土壤酶活性的季节动态. 生态环境, 2007, 16(3): 1024-1029. |
| [8] | 郑洪元, 张德生, 周维新, 张宪武. 森林有机残体分解与土壤酶活性. 土壤通报, 1983, 14(1): 36-39. |
| [9] | 张东来, 毛子军, 张玲, 朱胜英. 森林凋落物分解过程中酶活性研究进展. 林业科学, 2006, 42(1): 105-109. |
| [10] | 周正贤. 中国马尾松. 北京: 中国林业出版社, 2000. |
| [11] | 关松荫. 土壤酶及其研究法. 北京: 中国农业大学出版社, 1986. |
| [17] | 邓仁菊, 杨万勤, 吴福忠. 季节性冻融对岷江冷杉和白桦凋落物酶活性的影响. 应用生态学报, 2009, 20(5): 1026-1031. |
| [18] | 季晓燕, 江洪, 洪江华, 马元丹. 亚热带3种树种凋落叶厚度对其分解速率及酶活性的影响. 生态学报, 2013, 33(6): 1731-1739. |
| [21] | 陶宝先, 张金池, 愈元春, 丛日亮. 苏南丘陵地区森林土壤酶活性季节变化. 生态环境学报, 2010, 19(10): 2349-2354. |
| [23] | 熊浩仲, 王开运, 杨万勤. 川西亚高山冷杉林和白桦林土壤酶活性季节动态. 应用与环境生物学报, 2004, 10(4): 416-420. |
| [26] | 胡嵩, 张颖, 史荣久, 韩斯琴, 李慧, 徐慧. 长白山原始红松林次生演替过程中土壤微生物生物量和酶活性变化. 应用生态学报, 2013, 24(2): 366-372. |
| [29] | 胡亚林, 汪思龙, 黄宇, 于小军. 凋落物化学组成对土壤微生物学性状及土壤酶活性的影响. 生态学报, 2005, 25(10): 2662-2668. |
| [31] | 徐振锋, 唐正, 万川, 熊沛, 曹刚, 刘庆. 模拟增温对川西亚高山两类针叶林土壤酶活性的影响. 应用生态学报, 2010(11): 2727-2733. |
| [32] | 葛晓改, 肖文发, 曾立雄, 黄志霖, 黄玲玲, 谭本旺. 三峡库区不同林龄马尾松土壤养分与酶活性的关系. 应用生态学报, 2012, 23(2): 445-451. |