 2014, Vol. 34
2014, Vol. 34文章信息
- 陈云, 王海亮, 韩军旺, 韦博良, 贾宏汝, 叶永忠, 袁志良
- CHEN Yun, WANG Hailiang, HAN Junwang, WEI Boliang, JIA Hongru, YE Yongzhong, YUAN Zhiliang
- 小秦岭森林群落数量分类、排序及多样性垂直格局
- Numerical classification, ordination and species diversity along elevation gradients of the forest community in Xiaoqinling
- 生态学报, 2014, 34(8): 2068-2075
- Acta Ecologica Sinica, 2014, 34(8): 2068-2075
- http://dx.doi.org/10.5846/stxb201308162093
-
文章历史
- 收稿日期:2013-8-16
- 修订日期:2014-3-20






2. 河南小秦岭国家级自然保护区管理局, 灵宝 472500;
3. 郑州大学 离子束生物工程省重点实验室, 郑州 450052
2. Administration Bureau of Xiaoqinling National Nature Reserve of Henan, Lingbao 472500, China;
3. Henan Provincial Key Laboratory of Ion Beam Bio-engineering, Zhengzhou University, Zhengzhou 450052, China
植被群落分类与排序一直是植被生态学研究中最基本、最困难的问题[1]。数量分析则可以深刻地揭示植物种、植物群落与环境因子间的生态关系,是研究群落分类与排序的重要途径[2]。目前,常用的植被数量分类方法有很多,如排序、聚类、双向指示种分析(TWINSPAN)等,但这些方法均需要人为判别。多元回归树是一种较新的数量分类方法[3],无需人为确定分类结果,较以往的数量分类方法更客观,并在近来的相关研究实践中取得了很好的分类结果[4, 5]。
物种多样性海拔梯度格局研究可以更深入反映物种的生物学、生态学特性,分布状况及其对环境的适应性,对揭示生物多样性环境梯度变化规律具有重要意义[6],是了解山区生态系统功能并对其进行合理经营的重要依据[7]。但不同区域及不同生活型的物种由于受不同因子的制约呈现不同的梯度格局[8, 9, 10],因此,对特定山地开展物种多样性海拔格局的案例研究非常必要[11]。
小秦岭自然保护区位于豫、陕两省交界的灵宝市西部,东接崤山丘陵,西连秦岭主脉,南倚莽莽群山,北临涛涛黄河,属暖温带—亚热带过渡区[12]。该区生物资源丰富,区系成分复杂,植被垂直带谱明显,是开展森林群落及物种多样性垂直分布格局研究的理想场所。目前在本区开展的研究多是有关植物资源[12, 13, 14]、珍贵物种[15]、物种空间分布格局[16]等方面,而植物群落分类、排序及物种多样性格局的相关研究还没见报道。
本文以小秦岭自然保护区森林群落为研究对象,采用分层取样的方法,对56个样地的物种进行全面调查,运用MRT和DCA排序法综合分析森林群落之间及森林群落与环境之间的关系,并用GAM模型研究物种多样性沿海拔梯度分布格局,揭示植被及其环境因子空间分布格局,以期为小秦岭自然保护区的保护提供理论基础,同时也为阐明本区山地植物群落分布和多样性垂直分布格局积累基础资料。
1 研究区概况小秦岭国家级自然保护区的地理坐标为北纬34°23′—34°31′,东经110°23′—110°44′,保护区东西长31 km,南北宽12 km,总面积15160 hm2,森林覆盖率81.2%,其主峰老鸦岔垴高2413.8 m,为河南最高峰。本区气候属暖温带大陆性季风型半干旱气候,四季分明,年均温11.2—14.2 ℃,极端低温-17 ℃,极端高温42.7 ℃,无霜期170—215 d,年降水量620 mm左右,土壤类型南坡是棕壤、黄棕壤,北坡是褐土。
2 研究方法 2.1 样方调查2012年8月在研究区内选择具代表性的山体,从山麓1020 m到山顶2413 m沿海拔梯度设置样地,海拔每升高50 m在山体南、北两侧选择具代表性的群落分别设置1个20 m × 20 m的乔木样方,在乔木样方中选择典型群落设置1个5 m × 5 m 的灌木样方,在乔木样方的4个角分别设置1个1 m × 1 m的草本样方,共设置乔、灌、草样方336个。乔木样方采用GPS 定位,记录其海拔、坡度、坡向,及胸径≥1 cm 乔木种类、数量、胸径、树高和冠幅。灌木和草本样方分别记录物种名、多度、盖度和高度。
2.2 数据处理 2.2.1 重要值对每个样地分别计算乔木、灌木和草本的重要值,其公式如下[17]:
乔木、灌木= (相对多度+相对胸高断面积+相对盖度)/3
草本=(相对多度+相对盖度)/2
2.2.2 多样性指数分别计算每个样地内乔木、灌木和草本植物的生物多样性。
(1)Patrick 丰富度指数

(2)Shannon-Wiener多样性指数
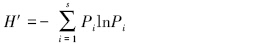
(3)Pielou均匀度指数
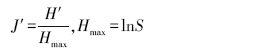
式中,S为样地内的物种数目;Pi为中i的相对重要值,Pi=Ni /N ;Ni为种N的重要值;N为种i所在样地的各个种的重要值之和。
2.3 多元回归树MRT是一种较新的研究物种分布与环境因子之间关系的数量分类方法。它将环境因子梯度作为分类节点,利用递归划分法,将样地划分为尽可能同质的类别,减少样地中群落的非相似性,同时采用交叉验证来确定分类结果,能很好地反映异质环境下群落的结构特征[3]。本研究采用56个样地海拔、坡向和坡度为自变量,405种植物在56个样地内的重要值为因变量进行群落分类。MRT采用R语言mvpart程序包进行运算[18]。
2.4 DCA排序除趋势对应分析(DCA)是以CA(Correspondence analysis,CA)和RA(Redundancy analysis,RA)为基础修改而成的,是植被分析中较为有效的一种方法,特别是同分类结合使用,效果要好于CCA (Canonical correspondence analysis,CCA)[19]。本研究用R语言vegan程序包[20],以405 × 56维重要值矩阵进行DCA数量排序,分析它们与环境因子之间的关系。
2.5 回归分析广义可加模型(GAM)是广义线性模型的进一步推广,是通过拟合非参数函数来估计响应变量与解释变量之间关系的模型,其拟合曲线能最大限度的符合原始数据。其一般形式为:

式中,α、β分别相当于线性回归模型中的截距和回归系数矩阵,XT 表示预测变量矩阵,E ( Y ) 为响应变量Y 的期望,g ( )表示联结函数。GAM分析用R语言gam程序包完成[21]。
采用GAM模型的偏差(D2)评价拟合结果,其计算公式如下:
D2=(无效偏差-剩余偏差)/无效偏差
式中,D2是指模型可解释的偏差,相当于线性回归模型中回归系数R2,无效偏差是指模型在只剩截距时的偏差,剩余偏差指拟合状态下仍不能解释的偏差[22]。
3 结果与分析 3.1 MRT数量分类将56个样地进行MRT分类,经交叉验证并依据植物群落分类和命名原则,本区植物群落可分为5类(图 1)。5种群落类型的总体多样性指数随海拔梯度变化情况如图 2所示。

|
| 图1 56个样地的多元回归树分类树状图 Fig. 1 Dendrogram of the multivariate regression trees classification of 56 samples n=样方个体数 |

|
| 图2 小秦岭自然保护区不同植物群落物种多样性海拔梯度的变化 Fig.2 Changes trend in species diversity of different plant community types along an altitudinal gradient in Xiaoqinling Nature Reserve |
Ⅰ 领春木(Euptelea pleiospermum)-葛萝槭(Acer grosseri)-活血丹(Glechoma longituba) + 白顶早熟禾(Poa acroleuca)群落,包括样地14—41,共28个样地。该群落分布于海拔1345—2048 m,土壤为棕壤或棕色森林土,气候冬冷夏凉、年降水量 1500 mm 左右。本带分布面积广,森林覆盖率高,林木蓄积量大,乔木层的优势种为领春木,主要伴生种为华山松(Pinus armandii)、野漆(Toxicodendron succedaneum)等;灌木层的优势种为葛萝槭,主要伴生种为多腺悬钩子(Rubus phoenicolasius)、连翘(Forsythia suspensa)、菝葜(Smilax china)等;草本层优势种为活血丹和白顶早熟禾,主要伴生种为蛇莓(Duchesnea indica)、宽叶薹草(Carex siderosticta)、细叶薹草(Carex duriusata subsp. stenophylloides)等。
Ⅱ 华山松(Pinus armandii) + 糙皮桦(Betula utilis)-东陵山柳(Salix phylicifolia)-宽叶薹草(Carex siderosticta)群落,包括样地44—56,共13个样地。该群落分布于海拔2100—2413 m,地势陡峻、岩石裸露、山风强烈,周围无高大林木,群落呈矮态。乔木层的优势种为华山松,主要伴生种为秦岭冷杉(Abies chensiensis )、红瑞木(Swida alba)、陕甘花楸(Sorbus koehneana)等;灌木层的优势种为东陵山柳,主要伴生种为多腺悬钩子、花楸树(Sorbus pohuashanensis)等;草本层优势种为宽叶薹草,主要伴生种为六叶葎(Galium asperuloides subsp. hoffmeisteri)、糙苏(Phlomis umbrosa)、紫菀(Aster tataricus)、茜草(Rubia cordifolia)等。
Ⅲ 蒙古栎(Quercus mongolica) -美蔷薇(Rosa bella)- 莎草(Cayerus compressus)群落,包括样地42、43,共2个样地。该群落分布于海拔2048—2100 m,土壤为棕色森林土,气候寒冷,风大,无霜期210 d,年降水量 1500 mm 左右。乔木层的优势种为蒙古栎,主要伴生种为华山松、青杨(Populus cathayana)、青楷槭(Acer tegmentosum)等;灌木层的优势种为美蔷薇,主要伴生种为槲栎(Quercus aliena )、山梅花(Philadelphus incanus)等;草本层的优势种为莎草,主要伴生种为宽叶薹草、盘果菊(Prenanthes tatarinowii)等。
Ⅳ 华山松(Pinus armandii) + 橿子栎(Quercus baronii var. baronii) + 黄栌(Cotinus coggygria)-连翘(Forsythia suspensa)-莎草(Cayerus compressus),包括样地1、2、3、5、6、9、12,共7个样地。该群落分布于1020—1345 m的阴坡上,与南坡水热条件存在差异,植被主要为华北暖温带的植物区系特征。乔木层的优势种为华山松,主要伴生种为油松(Pinus tabulaeformis)、野漆等;灌木层的优势种为连翘,主要伴生种为悬钩子等;草本层的优势种为莎草,主要伴生种为宽叶薹草、荩草(Arthraxon hispidus)、早熟禾、酢浆草(Oxalis corniculata)等。
Ⅴ 华山松(Pinus armandii) + 油松(Pinus tabulaeformis)-葛萝槭(Acer grosseri)-求米草(Oplismenus undulatifolius)群落,包括样地4、7、8、10、11、13,共6个样地。该群落位于海拔1020—1345 m的阳坡上,植被有亚热带特色。乔木层的优势种为华山松,主要伴生种为葛萝槭、野漆、黄橿子等;灌木层的优势种为葛萝槭,主要伴生种为粗榧(Cephalotaxus sinensis)、连翘等;草本层的优势种为求米草,主要伴生种为莎草、细叶薹草、荩草等。
3.2 样方DCA排序分析通过DCA分析得到前4个排序轴的特征值分别为:0.6759、0.6438、0.4918、0.4333。第一排序轴的特征值最大,第二轴次之,前两个排序轴所包含的生态信息量较大,能够体现出重要的生态意义。从56个样地DCA二维排序图可以看出(图 3),第一排序轴(横轴)基本上反映了海拔高度由高到低的变化梯度;第二排序轴(纵轴)反映了植物群落坡向由阴坡到阳坡的变化梯度。

|
| 图3 小秦岭自然保护区56个植物样地的DCA二维排序图 Fig.3 Two-dimensional DCA ordination diagram of 56 quadrats of plant community in Xiaoqinling Nature Reserve |
排序图中56个样地可以划分成A、B、C和D四个生态区,较好地反应了植物群落与环境因子之间的相互关系。A和B区主要包括群落Ⅴ和群落Ⅳ,分布在1020—1345 m之间;C区中主要为群落Ⅰ,分布在1345—2048 m之间,D区主要包括群落Ⅱ,分布在2100—2413 m之间。随着海拔高度的增加,环境中的水热因子等相应发生变化,群落中的植物种类组成也表现出一定的差异[23, 24]。例如群落中乔木的优势种由低海拔的华山松、黄橿子、油松,逐渐变为高海拔的华山松和糙皮桦;灌木低海拔的优势种连翘和葛萝槭,被分布于较高海拔的东陵山柳等取代;草本植物的优势种由莎草和求米草变为宽叶薹草。也就是说,随着海拔高度的增加,温度逐渐降低,群落优势种的耐寒性逐渐增强。
3.3 海拔对物种多样性分布的影响从表 1中可看出海拔与不同生活型物种的各多样性指数相关程度不同,GAM拟合模型中D2反应了模型可解释的偏差。乔木层3个指数的相关程度最大,D2介于13.1%到20.2%,灌木层3个指数的D2介于7.1%到8.6%,草本层3个指数的D2介于1.4%到9.3%。
|
自变量
Independent variable | 响应变量
Predicted variable | 无效偏差
Null deviance | 剩余偏差
Residual deviance | 偏差
D2 |
| 海拔Elevation | 乔木层丰富度指数 D1 | 90.127 | 72.018 | 0.201 |
| 海拔 | 灌木层丰富度指数 D2 | 111.151 | 102.832 | 0.075 |
| 海拔 | 草本层丰富度指数 D3 | 28.509 | 25.862 | 0.093 |
| 海拔 | 乔木层Shannon-Wiener指数H′1 | 6.742 | 5.381 | 0.202 |
| 海拔 | 灌木层Shannon-Wiener指数H′2 | 6.063 | 5.54 | 0.086 |
| 海拔 | 草本层Shannon-Wiener指数H′3 | 2.431 | 2.251 | 0.074 |
| 海拔 | 乔木层均匀度指数 J′1 | 1.236 | 1.074 | 0.131 |
| 海拔 | 灌木层均匀度指数 J′2 | 3.149 | 2.927 | 0.071 |
| 海拔 | 草本层均匀度指数 J′3 | 0.184 | 0.182 | 0.014 |
物种丰富度、Shannon-Wiener指数和均匀度指数随海拔梯度变化如图 4所示,从图中可以看出不同生活型的物种丰富度对环境变化的敏感程度不同。物种丰富度中,乔木层总体上中海拔区域较高,低海拔、高海拔区物种丰富度较低,其中海拔1345—2048 m的样地最高,该区乔木层物种比较丰富,亚优势种和伴生种较多,如华山松、野漆;灌木层物种丰富度在中海拔区域呈下降趋势,其余海拔下物种丰富度呈增长趋势;草本层物种丰富度随着海拔的升高总体呈下降趋势。Shannon-Wiener多样性指数中,乔木层、灌木层和草本层的Shannon-Wiener多样性指数随海拔变化趋势与物种丰富度变化趋势大体相同。物种均匀度中,乔木层、灌木层和草本层均匀度指数随海拔变化趋势较平缓。

|
| 图4 小秦岭自然保护区物种多样性海拔梯度的变化 Fig.4 Varition of species diversity along altitudinal gradient of Xiaoqinling Nature Reserve |
群落物种组成是沿着环境因子梯度连续渐变的,没有明显的界限,根据植物群落本身的特征进行群落划分困难很大。群落分类不仅要求种类组成和结构相对一致,占据的生境也是重要的特征[1]。在生境异质性比较大的地方,地形因子是形成生境的最根本因素之一[25]。因此,采用基于地形因子和物种组成数据的MRT技术将小秦岭自然护区56块样地的405种植物分成了5类,结果显示每个群落类型给都有各自的物种组成和群落特征,并占据着不同的生境。由图 2可看出5类群落丰富度指数、Shannon-Wiener指数和均匀度指数随海拔变化呈现出较为一致的变化趋势,即不对称的单峰分布格局,这是因为中海拔比高海拔地区水热条件更为适宜,相对于低海拔干扰较少有关。
DCA排序结果较好地反映了植物群落之间以及植物群落与环境因子之间的相互关系。MRT划分的5种群落类型与样地在DCA排序图上散布格局吻合也较好,说明同分类方法结合使用DCA效果较好,它较准确地反映出各森林群落之间以及森林群落与环境之间的关系。DCA第一排序轴主要反映了海拔的变化,第二排序轴主要反映了坡向的变化。海拔和坡向等生态因子对群落生境热量和湿度等的影响,导致群落类型分布呈现出一定的梯度变化。
MRT和DCA分析结果均表明海拔是影响小秦岭物种多样性分布格局的重要环境因素,为进一步揭示小秦岭自然保护区海拔因素对物种分布格局和多样性的影响,运用GAM模型拟合海拔与不同生活型物种各多样性指数的相关关系。乔木层多样性与海拔拟合模型的效果相对较好,特别是丰富度指数和Shannon-Wiener指数,D2值分别为20.1%、20.2%,这与假设是相符的,而灌木层、草本层与海拔相关程度较小,这是因为除海拔外,乔木层的熊高断面积也对灌木层和草本层的物种分布产生一定的影响,特别是草本层,F测验显示3个指数与海拔相关程度均不显著,这与很多研究是一致的,即沿环境梯度,不同生活型的物种多样性差异较大,木本植物具有比草本植物更为明确的分布格局[26]。另外拟合模型的偏差均小于1也说明除了除海拔外还有其他的因素影响物种的分布,这些都有待进一步研究。
在不同研究尺度和不同区域,植物多样性沿海拔梯度具有不同分布格局,主要有5种形式,分别是是随海拔升高先降低后升高、先升高后降低、单调升高、单调下降和没有明显格局[27]。小秦岭自然保护区物种多样性沿海拔梯度发生一定的波动,且乔、灌、草3层的物种多样性随海拔变化呈现不同的多样性格局。乔木层物种丰富度呈单峰曲线,即在中海拔物种丰富度最高,这是因为中海拔的水热条件适宜,相对于低海拔干扰较少有关。灌木层的丰富度在中海拔呈下降趋势,这可能是因为中海拔乔木层物种丰富,受郁闭度影响,限制了灌木的发育。草本层物种丰富度沿海拔梯度呈下降趋势,中海拔地段可能是受乔木层、灌木层郁闭度的影响,到高海拔地段,温度降低到只适合部分物种生长,所以物种丰富度也不高。Shannon-Wiener指数与物种丰富度变化趋势大体相同,这是因为两指数间有一定的相关性,还与个体数目或生物量等指标在各个物种中分布的均匀程度有关,这与相关研究一致[28, 29]。3个层次的均匀度指数随海拔的升高分布平缓,变化幅度均不是很大,群落的发展、替代速率较稳定,与干扰、水热条件、小生境的影响等有很大关系。
森林群落组成和物种多样性随海拔梯度垂直变化格局对揭示生物多样性环境梯度变化规律具有重要意义,该结果有助于深入理解保护区森林群落的空间分布格局及各物种的生态适应特性,为今后保护区管理和保护提供相应的理论依据,同时也为阐明山地植物群落分布和多样性的垂直分布格局积累基础资料。
| [1] | Wu Z Y. Vegetation of China. Beijing: Science Press, 1980: 1-1375. |
| [2] | Zhang J T. Quantitative Ecology. Beijing: Science Press, 2004: 1-372. |
| [3] | De'Ath G. Multivariate regression trees: a new technique for modeling species-environment relationships. Ecology, 2002, 83(4): 1105-1117. |
| [4] | Lai J S, Mi X C, Ren H B, Ma K P. Numerical classification of associations in subtropical evergreen broad-leaved forest based on multivariate regression trees―a case study of 24 hm2 Gutianshan forest plot in China. Chinese Journal of Plant Ecology, 2010, 34(7): 761-769. |
| [5] | Zhang R, Liu T. Plant species diversity and community classification in the southern Gurbantunggut Desert. Acta Ecologica Sinica, 2012, 32(19): 6056-6066. |
| [6] | Deng H B, Hao Z Q, Wang Q L. The changes of co-possession of plant species between communities with altitudes on northern slope of Changbai Mountain. Journal of Forestry Research, 2001, 12(2): 89-92. |
| [7] | Gaston K J. Global patterns in biodiversity. Nature, 2000, 405(6783): 220-227. |
| [8] | Rahbek C. The relationship among area, elevation, and regional species richness in neotropical birds. The American Naturalist, 1997, 149(5): 875-902. |
| [9] | Rahbek C. The role of spatial scale and the perception of large-scale species-richness patterns. Ecology Letters, 2005, 8(2): 224-239. |
| [10] | Lomolino M V. Elevation gradients of species-density: historical and prospective views. Global Ecology and Biogeography, 2001, 10(1): 3-13. |
| [11] | Zhu Y, Kang M Y, Jiang Y, Liu Q R. Altitudinal pattern of species diversity in woody plant communities of mountain Helan, northwestern China. Journal of Plant Ecology, 2008, 32(3): 574-581. |
| [12] | Ye Y Z, Yuan Z L, You Y, Li X W. A preliminary study of the bryoflora of Xiaoqinling Nature Reserve in Henan Province. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(8): 1472-1475. |
| [13] | Wang H L, Li D L. Distribution and utilization of euonymus in Henan Xiaoqinling. Hebei Jouranl of Forestry and Orchard Research, 2010, 25(4): 348-351. |
| [14] | Huang D, Wang J, Li D L, Wang H L. Distribution and utilization of Lonicera in Xiaoqinling, Henan. Chinese Wild Plant Resources, 2010, 29(1): 28-31. |
| [15] | Wang H L, Yao G Z, Han J W, Zhang Y, Zhang Y F. Henan Xiaoqinling national nature reserve Abies Chensiensis growth investigates and analyzes. Journal of Henan Forestry Science and Technology, 2006, 26(3): 24-25. |
| [16] | Wang H L, Zhuang J J, Zhang K. Major species spatial distribution pattern of the transit station plots in Henan Xiaoqinling Nature Reserve. Journal of Henan Agricultural University, 2012, 46(4): 385-391. |
| [17] | Zhang X P, Wang M B, She B, Xiao Y. Quantitative classification and ordination of forest communities in Pangquangou National Nature Reserve. Acta Ecologica Sinica, 2006, 26(3): 754-761. |
| [18] | De'ath G. Package‘mvpart’. 2010. |
| [19] | Su R G G, Zhang J T, Zhang B, Cheng J J, Tian S G, Zhang Q D, Liu S J. Numerical classification and ordination of forest communities in the Songshan National Nature Reserve. Acta Ecologica Sinica, 2010, 30(10): 2621-2629. |
| [20] | Oksanen J, Blanchet F G, Kindt R, Legendre P, O'Hara R B, Simpson G L, Solymos P, Stevens M H H, Wagner H. Package‘vegan’. 2010. |
| [21] | Trevor Hastie, Package‘gam’. 2013. |
| [22] | Wang S P, Zhang Z Q, Zhang J J, Zhu J Z, Guo J T, Tang Y. Modeling the effects of topography on the spatial distribution of secondary species in a small watershed in the Gully region on the Loess Plateau, China. Acta Ecologica Sinica, 2010, 30(22): 6102-6112. |
| [23] | Wimberly M C, Spies T A. Influences of environment and disturbance on forest patterns in coastal Oregon watersheds. Ecology, 2001, 82(5): 1443-1459. |
| [24] | Zhao C M, Chen W L, Tian Z Q, Xie Z Q. Altitudinal pattern of plant species diversity in Shennongjia Mountains, Central China. Journal of Integrative Plant Biology, 2005, 47(12): 1431-1449. |
| [25] | Fang J Y, Shen Z H, Cui H T. Ecological characteristics of mountains and research issues of mountain ecology. Biodiversity Science, 2004, 12(1): 10-19. |
| [26] | O'Brien E M. Climatic gradients in woody plant species richness: towards an explanation based on an analysis of Southern Africa's woody flora. Journal of Biogeography, 1993, 20(2): 181-198. |
| [27] | He J S, Chen W L. A review of gradient changes in species diversity of land plant communitles. Acta Ecologica Sinica, 1997, 17(1): 91-99. |
| [28] | Ojeda F, Maranon T, Arroyo J. Plant diversity patterns in the Aljibe Mountains (S. Spain): a comprehensive account. Biodiversity and Conservation, 2000, 9(9): 1323-1343. |
| [29] | Ma K P, Huang J H, Yu S L, Chen L Z. Plant community diversity in Dongling Mountain, Beijing, China Ⅱ.Species richness, evenness and species diversities. Acta Ecologica Sinica, 1995, 15(3): 268-277. |
| [1] | 吴征镒. 中国植被. 科学出版社, 1980: 1-1375. |
| [2] | 张金屯. 数量生态学. 科学出版社, 2004: 1-372. |
| [4] | 赖江山, 米湘成, 任海保, 马克平. 基于多元回归树的常绿阔叶林群丛数量分类——以古田山24公顷森林样地为例. 植物生态学报, 2010, 34(7): 761-769. |
| [5] | 张荣, 刘彤. 古尔班通古特沙漠南部植物多样性及群落分类. 生态学报, 2012, 32(19): 6056-6066. |
| [11] | 朱源, 康慕谊, 江源, 刘全儒. 贺兰山木本植物群落物种多样性的海拔格局. 植物生态学报, 2008, 32(3): 574-581. |
| [12] | 叶永忠, 袁志良, 尤扬, 李孝伟. 小秦岭自然保护区苔藓植物区系分析. 西北植物学报, 2004, 24(8): 1472-1475. |
| [13] | 王海亮, 李冬林. 河南小秦岭卫矛属植物资源的分布与开发利用研究. 河北林果研究, 2010, 25(4): 348-351. |
| [14] | 黄栋, 王谨, 李冬林, 王海亮. 河南小秦岭忍冬属植物的分布及资源利用探讨. 中国野生植物资源, 2010, 29(1): 28-31. |
| [15] | 王海亮, 姚冠忠, 韩军旺, 张莹, 张亚芳. 河南小秦岭国家级自然保护区秦岭冷杉生长调查与分析. 河南林业科技, 2006, 26(3): 24-25. |
| [16] | 王海亮, 庄静静, 张凯. 河南小秦岭自然保护区西曼林区样地主要物种的空间分布格局. 河南农业大学学报, 2012, 46(4): 385-391. |
| [19] | 苏日古嘎, 张金屯, 张斌, 程佳佳, 田世广, 张钦弟, 刘素军. 松山自然保护区森林群落的数量分类和排序. 生态学报, 2010, 30(10): 2621-2629. |
| [22] | 王盛平, 张志强, 张建军, 朱金兆, 郭军庭, 唐寅. 黄土残塬沟壑区流域次生植被物种分布的地形响应. 生态学报, 2010, 30(22): 6102-6112. |
| [25] | 方精云, 沈泽昊, 崔海亭. 试论山地的生态特征及山地生态学的研究内容. 生物多样性, 2004, 12(1): 10-19. |
| [27] | 贺金生, 陈伟烈. 陆地植物群落物种多样性的梯度变化特征. 生态学报, 1997, 17(1): 91-99. |
| [29] | 马克平, 黄建辉, 于顺利, 陈灵芝. 北京东灵山地区植物群落多样性的研究Ⅱ丰富度、均匀度和物种多样性指数. 生态学报, 1995, 15(3): 268-277. |