 2014, Vol. 34
2014, Vol. 34文章信息
- 张淑勇, 夏江宝, 张光灿, 王冰, 赵洪峰, 刘京涛
- ZHANG Shuyong, XIA Jiangbao, ZHANG Guangcan, WANG Bing, ZHAO Hongfeng, LIU Jingtao
- 黄河三角洲贝壳堤岛叶底珠叶片光合作用对CO2浓度及土壤水分的响应
- Photosynthetic responses to changes in CO2 concentration and soil moisture in leaves of Securinega suffruticosa from shell ridge islands in the Yellow River Delta, China
- 生态学报, 2014, 34(8): 1937-1945
- Acta Ecologica Sinica, 2014, 34(8): 1937-1945
- http://dx.doi.org/10.5846/stxb201305060935
-
文章历史
- 收稿日期:2013-5-6
- 修订日期:2013-11-5






2. 山东农业大学林学院、山东省土壤侵蚀与生态修复重点实验室, 泰安 271018;
3. 山东省邹平水务局, 邹平 256200
2. Shandong Provincial Key Laboratory of Soil Erosion and Ecological Restoration, Forestry College, Shandong Agricultural University, Taian 271018, China;
3. Zouping Water Conservancy Department, Zouping 256200, China
The study materials were 2-year-old plants of S. suffruticosa, originally from the shell islands in the Yellow River Delta. These plants were obtained from a nursery. The plants were subjected to seven different levels of water supply using an artificial water control system in a covered shed, and to a series of CO2 concentrations controlled using a LI-COR 6400 portable photosynthesis system. The objective of this study was to investigate photosynthetic responses to CO2 concentrations and determine whether photosynthetic productivity was quantitatively related to soil moisture content. The photosynthetic rate (Pn) and characteristic parameters of photosynthesis showed threshold-value responses to variations in soil relative water content (SRWC). S. suffruticosa plants were able to maintain higher photosynthetic productivity in the range of 50.3% to 83.2% RWC; in this range, the range of suitable CO2 concentrations was 700 to 1100 μmol/mol, and Pn increased with increasing CO2 concentrations. The most suitable SRWC was 70.5%, and Pn peaked at the CO2 concentration of 900 μmol/mol. The characteristic parameters of photosynthesis showed different responses to changes in SRWC. Under normal and double CO2 concentrations, the ranges of SRWC in which S. suffruticosa maintained higher photosynthetic productivity were 45.5%-90.0% and 47.0%-92.6%, respectively, indicating that S. suffruticosa was highly adaptable to changes in soil moisture content. The apparent maximum photosynthesis rate (Pmax,c) and carboxylation efficiency (CE) increased with increasing SRWC, peaking at 70.5% SRWC and then decreasing slowly at higher SRWC. The CO2compensation point showed the opposite trend to that of Pmax,c and CE, and reached a minimum value (approx. 51.3 μmol/mol) at 70.5% SRWC. The photorespiration rate increased slowly with increasing SRWC and peaked at approximately 2.6175 μmol·m-2·s-1 at 50.3% SRWC.
We concluded that photosynthesis in leaves of S. suffruticosa is widely adaptable to changes in CO2 concentrations and soil moisture, and that this species has the typical physiological characteristics of a water-tolerant and drought-tolerant plant. Thus, S. suffruticosa has the potential for cultivation on the shell ridge islands of the Yellow River Delta, China.
植物光合生理活动受多种因素的影响,其中土壤水分、CO2浓度和光合有效辐射是植物生理活动的物质和能量基础,也是影响植物生长、发育、繁殖和分布的重要生态环境因子,因此研究植物光合作用对CO2浓度和土壤水分的响应特征是研究生态系统碳循环和水循环的基础[1]。特别是目前大气CO2浓度逐年升高,导致全球雨热分布的时空格局也发生巨大变化,引起不同地区水资源改变,由此将导致地球上大多数干旱地区更加干旱[2, 3],植物光合作用对CO2浓度和土壤水分的响应特征研究对全球变化条件下食物安全保障、生态环境建设与改善都具有重要现实意义。
贝壳堤是淤泥质或粉质海岸所特有的一种滩脊类型,近年来受自然因素和人类活动的过度干扰,贝壳堤岛脆弱生态系统受到严重破坏,虽然植被恢复措施较为明显,但仍存在树种单一、成活率低和生长缓慢等问题,原因之一是对优良乡土树种认识不足,特别是在黄河三角洲贝壳堤岛滩脊地带淡水资源缺乏的条件下,亟需开展植物光合生理特性与土壤水分作用关系的研究。研究植物如何适应土壤干旱、光因子胁迫及大气CO2浓度升高是揭示其对生存环境生态适应性机制的有效途径。研究表明,CO2浓度升高提高了植物的光合作用,增大了气孔阻力,减少蒸腾作用,并可明显提高水分利用效率[4, 5, 6]。Vurro等[7]的研究表明,较高浓度的CO2对O3造成的伤害有改善作用,也有研究表明大气中CO2倍增对植物盐害具有缓解效应[8],因此,CO2浓度升高有可能增强植物的抗逆性。鉴于黄河三角洲贝壳堤岛滩脊地带植物生长的限制因子主要是水分,因此研究植物对CO2浓度及土壤水分的响应和适应机制十分必要。
国内外学者就光合作用对CO2浓度变化的响应特征进行了大量研究[6, 7, 8, 9, 10]。但大多数研究多为在简单的CO2倍增或自然光照条件下,或仅考虑少数几个水分胁迫和CO2浓度升高的作用,缺乏在系列土壤水分梯度和CO2浓度连续变化条件下,植物光合生理生态特性对土壤水分和CO2浓度响应规律的研究。因此,本文选用黄河三角洲优良的天然次生灌木叶底珠(Securinega suffruticosa)作为研究对象,监测其光合特性对土壤水分和CO2浓度的响应特征,研究其光合作用参数与土壤水分的定量关系,探讨贝壳砂干湿交替生境条件下叶底珠光合的适应特点,为恢复和营造该区的自然植被、保护和改善生态环境、促进水资源的可持续利用提供参考数据。
1 试验地概况研究区位于山东滨州市无棣县中东部滨海低地的汪子岛(N38°02′51″—38°21′06″,E117°46′58″—118°05′43″),属滨州贝壳堤岛与湿地国家级自然保护区。该保护区属于暖温带东亚季风大陆性半湿润气候区,平均降水量为550 mm,主要集中在6—9月份,占全年降水量的71%以上;平均蒸发量为2430.6 mm,蒸降比为4.4。平均气温为12.36 ℃,极端最低气温为-25.3 ℃,极端最高气温为37.5 ℃;多年平均日照2849 h/a,平均无霜期205 d。贝壳堤岛地势平坦,海拔一般在5 m以下,潜水水位浅,矿化度高。贝壳砂平均厚度达1.0—2.5 m,局部达3—4 m,土壤类别主要是贝壳砂土类和滨海盐土类,向海侧和向陆侧以滨海盐渍土为主,成土母质由风积物和钙质贝壳土壤化组成。植被类型以草本、灌木为主,受自然因素和平岛挖砂等人类活动的影响,现有树种较少。
2 材料与方法 2.1 试验材料与水分控制试验材料为2年生叶底珠苗木,平均苗高(75.8±3.1) cm,平均地径(1.15±0.27) cm,苗木定植前,冲洗根系后用吸水纸吸去其表面水分,迅速用电子天平称量鲜重,以计算每株苗木干重。栽培基质为贝壳砂,试验期间栽培基质保持自然肥力,不施肥。试验于2012年3月下旬进行布设,每一处理(水平)重复6次,共42盆,栽培容器高70 cm、口径45 cm,下部有排水通气孔。
苗木成活后,充分浇水待重力水渗出后测定其含水量,在第1、3、5、8、11、15、20天后采用烘干法分别测其土壤含水量(MWC;%),土壤相对含水量(SRWC;%)分别为(94.4±0.2)%、(83.2±0.3)%、(70.5±0.4)%、(62.7±0.3)%、(50.3±0.3)%、(37.9±0.2)%和(23.6±0.4)%。通过自然蒸发和补充灌水使土壤水分含量维持基本不变,不同水分条件处理3个月,7月5日开始对苗木叶片光合生理参数进行测定。各处理生长阶段土壤相对含水量基本不变,为避免因幼树生长对土壤含水量控制水平的影响,每7 d采用标准株法对备用幼树地上部分生物量增量(鲜重)进行调查,以便调整水分控制标准(栽培桶总重量),土壤相对含水量采用称重法控制(ES50K-15型电子天平,感量0.5 g),每2 d称量1次,以便及时调整每日补充灌水量,为保持各处理土壤水分相对稳定,除每天补充灌水外,土壤表面用小砂石覆盖。
用环刀法测得盆栽试验土壤的田间持水量(FC)为(26.1±1.3)%,土壤密度为(1.23±0.11) g/cm3。其中,SRWC=MWC/FC。
2.2 观测指标及方法从叶底珠苗木中部选10片生长健壮的成熟叶片,并做好标记,在不同时间观测时皆用同一叶片。用LI-6400便携式光合测定仪(美国Licor公司生产)对不同供水条件下苗木的气体交换参数以及相应的生态环境要素进行测定。为了尽量减少外界光照波动所造成的影响,选择完全晴朗天气(2012年7月15日—20日)的8:30—11:00时进行测定,每张叶片重复3—5次,取平均值。在苗木气体交换参数的测定过程中,田间气温为(32.4±2.7) ℃,叶室温度为(30.0±0.4) ℃,叶室相对湿度为(36.3±2.7)%,光合测定仪的流速设定为200 μmol/s,光量子通量密度为1200 μmol · m-2 · s-1,参比室CO2浓度设为1400、1200、1000、800、600、400、250、200、150、100、50和20 μmol/mol等12个水平。在每个CO2浓度下,时间控制在5—6 min,数据稳定后记录。仪器自动记录净光合速率(Pn,μmol · m-2 · s-1)、气孔导度(Gs,mol · m-2 · s-1)、胞间CO2浓度( Ci,μmol/mol)等生理参数以及大气温度(Tc,℃)、相对湿度(RH,%)、光合有效辐射(PAR,μmol · m-2 · s-1)等环境因子。
2.3 数据计算与处理应用SPSS软件对试验数据进行方差分析和回归分析。采用Michaelis-Menten方程[11]拟合Pn的CO2响应曲线,计算其表观最大净光合速率,方程为:
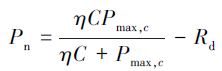
式中,Pmax,c是在一定光强下CO2浓度达到饱和时的表观最大净光合速率,C是CO2浓度,η是表观羧化效率。
根据Farquhar等[12],在Pn-Ci响应曲线的较低CO2浓度下(Ci <200 μmol/mol),Pn主要受Rubisco活性和数量的限制,净光合速率Pn为:
Pn=CE×Ci-Rp
式中,CE为羧化效率,Rp为光下呼吸速率。由于光下暗呼吸很小,可以近似将光下叶片向无CO2的空气中释放CO2的速率看作光呼吸速率[13, 14]。
3 结果与分析在不同土壤水分条件下,Pn对低CO2浓度(Ci<200 μmol/mol)的响应规律基本相似(图 1),即Pn随着CO2浓度的增加呈正比迅速上升,受土壤水分的影响较小,表明此时CO2浓度可能是光合作用的主要限制因子。随着CO2浓度的增加,不同土壤水分条件下Pn对CO2浓度的响应表现出不同的变化规律。首先,在SRWC>70.5%时,Pn随着CO2浓度的增加而上升,当CO2浓度达到1000 μmol/mol左右时,Pn出现饱和现象,然后略为下降,Pn没有显著差异(P>0.05),维持在较高水平。其次,在23.6%<SRWC≤70.5%的水分范围内,Pn随着CO2浓度的增加而上升,当CO2浓度达到饱和后,Pn出现迅速下降的趋势,这可能是由于叶片来不及吸收和利用较高浓度的CO2,同时CO2同化过程的一系列酶促反应速率跟不上而成为了Pn的限制步骤[15];在此水分范围内,Pn下降的幅度各处理有所差异,随着SRWC的增加,下降的幅度逐渐减小,如SRWC分别为50.3%和70.5%时,Pn下降的幅度分别为26.9%和1.5%。在SRWC<37.9%(达到23.6%)时,随着CO2浓度的增加Pn一直维持在较低的水平,说明在CO2充足的条件下,土壤水分是Pn主要限制因素。Pn对SRWC的响应表现为,在SRWC<70.5%时,Pn随着SRWC的增大而上升;此后随着SRWC的增大,Pn反而下降,因此SRWC为70.5%可作为Pn变化的转折点。

|
| 图 1 不同土壤水分条件下净光合速率对细胞间隙CO2浓度的响应 Fig. 1 Responses of net photosynthetic rate (Pn) to intercellular CO2 concentration under different soil moisture condition. Points are the mean of replicate Pn responses for each Ci. Vertical bars indicate ±1 SE of the mean |
以上分析表明,在不同CO2浓度条件下,如果要提高叶底珠的光合生产力,必须有适宜的土壤水分条件进行耦合,维持较高光合生产力的土壤水分条件SRWC为50.3%—83.2%,此水分范围内适宜的CO2浓度为700—1100 μmol/mol,其中Pn最大值出现在SRWC为70.5%左右,对应CO2浓度为900 μmol/mol左右。在土壤水分条件稍充足或者轻度水分胁迫时,可获得中等的光合生产力,但当SRWC<37.9%时,Pn明显降低,叶底珠的正常生长受到明显抑制。
3.2 净光合速率的土壤水分阈值为揭示叶底珠苗木Pn发生显著变化的土壤水分临界点,取正常CO2浓度(350 μmol/mol)和倍增CO2浓度(700 μmol/mol)时对应的Pn 值进行分析,其Pn对SRWC的响应结果符合二次方程(图 2)。由此方程可确定出正常CO2浓度和倍增CO2浓度条件下,叶底珠 最大Pn分别为7.6 μmol · m-2 · s-1和11.2 μmol · m-2 · s-1,维持Pn最高水平的土壤含水量分别在SRWC为67.8%和69.8%; CO2浓度为350 μmol/mol,Pn为零时对应的两个土壤含水量值分别为21.2%和114.3%,CO2浓度为700 μmol/mol,Pn为零时对应的两个土壤含水量值分别为22.5%和117.2%。SRWC为114.3%和117.2%超过100%,所以无实际的生物学意义,需删除,因此,SRWC低于21.2%和22.5%时,叶底珠叶片均不能进行光合作用。根据拟合方程的积分式:
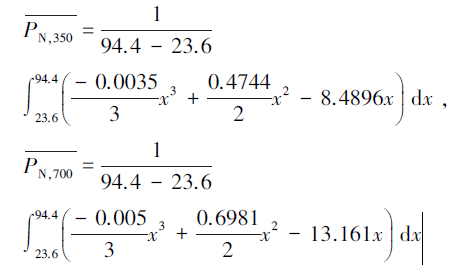

|
| 图 2 正常CO2浓度 (350 μmol/mol) 和倍增CO2 (700 μmol/mol) 浓度下浄光合速率 (Pn) 的土壤水分 (RWC) 阈值 Fig. 2 The threshold value of net photosynthetic rate (Pn) in response to relative soil water content (RWC) under: the normal CO2 concentration (350 μmol/mol) and twice the normal CO2 concentration (700 μmol/mol) |
求出试验土壤含水量范围内(SRWC为23.6%—94.4%),CO2浓度为350 μmol/mol 和700 μmol/mol时Pn的平均值分别为5.9 μmol · m-2 · s-1和8.6 μmol · m-2 · s-1,其对应的SRWC分别为45.5%和90.0%;47.0%和92.6%,因此,正常CO2浓度下维持叶底珠较高光合生产力水平的SRWC范围在45.5%—90.0%,其中最适宜的SRWC为67.8%。随着CO2浓度的增加,Pn平均值明显提高,增加的幅度在51%左右,但土壤水分范围及最适宜的土壤水分增加不明显,仅增加了2%。说明在适宜的土壤水分条件下,CO2浓度是光合作用的主要限制因素。
3.3 光合特征参数对土壤水分的响应叶底珠苗木光合-CO2响应特征参数对SRWC响应规律有所差异(图 3),表观最大净光合速率(Pmax,c)和羧化效率(CE)具有较为相似的变化规律,均呈抛物线趋势,即随着SRWC的增加,Pmax,c和CE呈直线增加的趋势,如SRWC由50.3%到70.5%时,Pmax,c和CE分别增加了49.8%和37.1%;当SRWC增加到一定临界值(约71%)时,两者均达到最大值,分别为20.6 μmol · m-2 · s-1和0.053 μmol · m-2 · s-1(表 1);随着SRWC的持续增加, Pmax,c和CE反而呈现迅速下降的趋势,表明SRWC过高或过低均会导致叶底珠最大光合能力和对CO2的同化能力降低。
CO2补偿点(Г)对SRWC的响应过程(图 3)与Pmax,c和CE呈现相反的规律,呈反抛物线变化。在SRWC为70.5%左右时,Г达到最低值(51.3 μmol/mol)(表 1)。在SRWC为50.3%—83.2%时,Г差异不显著(P>0.05)。随着SRWC的增加或减少,Г均明显增大,在SRWC为23.6%时,Г达到最大值(134.9 μmol/mol)(表 1)。
光呼吸速率(Rp)对SRWC的变化具有阈值响应(图 3)。在SRWC为50.3%,Rp达到最小值(2.6175 μmol · m-2 · s-1)(表 1),随着SRWC的增加,Rp缓慢增加,如SRWC为62.7%时,Rp增加了6%左右;但随着SRWC的下降,Rp增加明显,如SRWC为37.9%时,Rp增加了21.2%。表明适度的水分胁迫迫使其呼吸消耗减小,有利于光合产物的积累。
4 讨论与结论植物的生理过程对大气CO2浓度变化非常敏感,尤其是光合作用、蒸腾速率和羧化效率,会在很短的时间内做出响应[16, 17]。研究表明,短期CO2浓度升高在一定程度上能够促进植物的光合作用、增加气孔阻力、减少水分蒸腾[4, 5, 6, 7],本研究也证明了这一点。随着CO2浓度的增加,叶底珠苗木的Pn逐渐增加,CO2浓度增加对Pn的正效应会减缓干旱对光合作用的不利影响,以增强植物对干旱胁迫的抵御能力。随着土壤水分含量的降低,Pn的CO2饱和点缓慢下降到直线下降。较低水分条件下Pn增加幅度较小,因此在CO2浓度增加的情况下,应该增加水分灌溉,以充分利用CO2浓度升高对Pn的正效应。上 述分析表明,在水分充足或较低水分条件下,大气CO2浓度升高都能明显提高植物的Pn。大量研究表明,植物对土壤水分亏缺有一定的适应性和抵抗性,植物比较活跃的各种生理活动是在适度的水分亏缺范围之内[15],这一范围因植物种类及其生理过程不同而异。本研究表明,在不同的CO2浓度范围内,维持叶底珠苗木Pn较高的SRWC在50.3%—83.2%,其中最适宜光合作用的SRWC为70.5%(图 1)。这一范围既保证了叶底珠苗木维持较高的光合作用水平和正常的生长发育,又防止了蒸腾作用引起的大量耗水,从而达到水分的高效利用。
|
SRWC
/% | 拟合方程
Simulated equations | Pmax,c
/(μmol · m-2 · s-1) | CE
/(μmol · m-2 · s-1) | Rp
/(μmol · m-2 · s-1) | Г
/(μmol/mol) | R2 |
| 不同小写字母表示同一列数据在0.05水平上存在显著性差异; SRWC:土壤相对含水量Relative water content; Pmax,c: 最大净光合速率Apparent maximum photosynthesis rates; CE: 羧化效率 Carboxylation efficiency; Rp: 光呼吸速率 Photorespiration rate; Г: CO2补偿点 CO2 compensation points | ||||||
| 94.4 | Pn = 0.0367×Ci - 3.0311 | 16.8c | 0.0367c | 3.0311b | 82.6c | 0.9847 |
| 83.2 | Pn = 0.0420×Ci - 2.9318 | 19.3b | 0.0420b | 2.9318b | 69.8d | 0.9990 |
| 70.5 | Pn = 0.0525×Ci - 2.6953 | 20.6a | 0.0525a | 2.6953d | 51.3e | 0.9854 |
| 62.7 | Pn= 0.0457×Ci - 2.8161 | 18.3b | 0.0457b | 2.8161c | 61.6d | 0.9960 |
| 50.3 | Pn= 0.0383×Ci - 2.6175 | 13.8d | 0.0383c | 2.6175d | 68.3d | 0.9904 |
| 37.9 | Pn = 0.0311×Ci - 3.1730 | 10.2e | 0.0311d | 3.1730a | 102.0b | 0.9924 |
| 23.6 | Pn = 0.0223×Ci - 3.0074 | 7.6f | 0.0223e | 3.0074b | 134.9a | 0.9733 |

|
| 图 3 表观最大浄光合速率 (Pmax,c) 、羧化效率 (CE) 、CO2补偿点 (Г) 及光呼吸速率 (Rp) 对土壤水分的响应 Fig. 3 The responses of apparent maximum net photosynthesis rate (Pmax,c) ,Carboxylation velocity (Vc) ,CO2 compensation point (Г) ,and photorespiration rate (Rp) to soil relative water content (SRWC) |
CO2浓度饱和时的最大净光合速率在一定程度上反映了净光合速率对短期CO2浓度达到饱和时的潜在光合能力。本研究表明,叶底珠苗木在不同水分条件下的Pmax,c具有明显的阈值响应,在SRWC为70%左右时达到最大值,水分增加或减少都会显著降低Pmax,c。这可能是由于适宜的土壤水分含量有利于碳同化速率的增加,即增加了CO2对Rubisco酶结合位点的竞争从而提高CE,以及通过抑制光呼吸提高净光合速率[17]。植物的光合能力是用来表征不同植物或者作物品种的重要生理指标,在最佳或者最适的环境条件下,叶片的最大净光合速率可反映叶片的最大光合能力[18, 19]。不同植物的最大光合能力有所差异,杨梅(Myrica rubra)[20]、芦苇(Phragmites australis)[21]和胡杨(Populus euphratica oliv)[22]的最大光合能力分别为41.2 μmol · m-2 · s-1、51.8 μmol · m-2 · s-1、26.7 μmol · m-2 · s-1。本研究得出,叶底珠苗木的最大光合能力为20.6 μmol · m-2 · s-1,明显低于以上几种植物的光合能力,可见不同植物对高浓度CO2响应所表现出的光合特性差异较大[23, 24]。不同植物光合能力随着CO2浓度的增加而增加,但不同的植物增加程度不同,尤其是不同光合途径的植物差异很明显。大多数有关C4植物的研究表明,大气CO2浓度升高对C4植物的光合作用及生长没有很大促进[23, 24, 25, 26]。对CAM(景天酸代谢)植物的研究相对较少,其结果也不尽一致[26, 27, 28]。Cure[23]和Kimball[26]等人综合了大量的实验结果计算出CO2浓度倍增后,C4植物净光合速率只增加了4%,而C3植物的净光合速率则可提高66%。通过本研究结果来看,叶底珠苗木在CO2倍增条件下,除重度水分胁迫下,最大净光合速率增幅均超过40%,在SRWC为96.2%时,增幅为75%,与Cure[23]、Kimball[26]等人的研究结果基本一致。
CE反映了植物在给定条件下对CO2的同化能力。本研究结果表明,叶底珠苗木CE对SRWC具有一定的阈值响应,在SRWC为62.7%—70.5%时达到较高值(平均为0.049 μmol · m-2 · s-1),超过此范围,CE均会下降,且CE在高水分条件下降的幅度明显小于在低水分条件,表明低水分条件降低了苗木对CO2同化能力。CO2补偿点(Г)是了解和衡量光合作用和呼吸作用两者关系的一个重要生理指标,在不同时期和不同环境条件下C3植物对CO2响应有明显的变化[29]。在一定的温度范围内,Г随温度升高而升高,随着CO2浓度的降低而下降[29, 30]。本研究结果表明,叶底珠苗木在SRWC为50.3%—83.2%时,Г相差不大,变动幅度在51.3—69.8 μmol/mol之间,随着土壤水分含量的降低,Г明显增加,在严重水分胁迫下达到134.9 μmol/mol。可见叶底珠苗木在不同水分条件下对CO2浓度表现出一定的弹性适应。不同植物对CO2的同化能力以及Г有所差异,这可能与植物本身的特性有关,也可能与植物所处的环境条件有关。光呼吸曾被认为是对光能的消耗,不利于干物质积累[31, 32]。而随着研究的深入,逐渐认识到光呼吸对保证植物进行正常的光合作用是十分重要的[33]。但对于光呼吸的光保护作用一直存在争议[34, 35, 36, 37]。本研究结果表明,在SRWC为50.3%—70.5%时,Rp维持在较低值,以提高植物的生物产量,有利于干物质的积累。随着土壤水分含量的降低,Rp急剧增加,以保证在过剩光能下光合机构的正常运行,这与Kozaki[34]及Wingler等[35]人的研究结果一致,光呼吸很可能是一种耗散过剩光能以保护光合机构免受强光破坏的途径。
| [1] | Mateos-Naranjo E, Redondo-Gómez S, Andrades-Moreno L, Davy A J. Growth and photosynthetic responses of the cordgrass Spartina maritima to CO2 enrichment and salinity. Chemosphere, 2010, 81(6): 725-731. |
| [2] | Lenihan J M, Bachelet D, Neilson R P, Drapek R. Simulated response of conterminous United States ecosystems to climate change at different levels of fire suppression, CO2 emission rate, and growth response to CO2. Global and Planetary Change, 2008, 64(1/2): 16-25. |
| [3] | Xiao G J, Zhang Q, Wang J. Impact of global climate change on agro-ecosystem: A review. Chinese Journal of Applied Ecology, 2007, 18(8): 1877-1885. |
| [4] | Allen L H Jr, Kakani V G, Vu J C V, Boote K J. Elevated CO2 increases water use efficiency by sustaining photosynthesis of water-limited maize and sorghum. Journal of Plant Physiology, 2011, 168(16): 1909-1918. |
| [5] | Zhang Y B, Duan B L, Qiao Y Z, Wang K Y, Korpelainen H, Li C Y. Leaf photosynthesis of Betula albosinensis seedlings as affected by elevated CO2 and planting density. Forest Ecology and Management, 2008, 255(5/6): 1937-1944. |
| [6] | Sinha P G, Saradhi P P, Uprety D C, Bhatnagar A K. Effect of elevated CO2 concentration on photosynthesis and flowering in three wheat species belonging to different ploidies. Agriculture, Ecosystems & Environment, 2011, 142(3/4): 432-436. |
| [7] | Vurro E, Bruni R, Bianchi A, di Toppi L. Elevated atmospheric CO2 decreases oxidative stress and increases essential oil yield in leaves of Thymus vulgaris grown in a mini-FACE system. Environmental and Experimental Botany, 2009, 65(1): 99-106. |
| [8] | Elizabeth A A, Alistair R. The response of photosynthesis and stomatal conductance to rising CO2: Mechanisms and environmental interactions. Plant, Cell and Environment, 2007, 30(3): 258-270. |
| [9] | Zhang X C, Yu X F, Ma Y F, Zhangguan Z P. The responses of photosynthetic energy use in wheat flag leaves to nitrogen application rates and light density under elevated atmospheric CO2 concentration. Acta Ecologica Sinica, 2011, 31(4): 1046-1057. |
| [10] | Zhou X R, Wang J H, Zhang H, Wang J C, Zhang Y B. Effects of elevated CO2 and nitrogen deposition on leaf nutrient quality of Fargesia rufa Yi. Acta Ecologica Sinica, 2012, 32(24): 7644-7653. |
| [11] | Thomley J H M. Mathematical models in plant physiology: a qualitative approach to problems. London; New York: Academic Press, 1976 |
| [12] | Farquhar G D, von Caemmerer S, Berry J A. A biochemical model of photosynthetic CO2 assimilation in leaves of C3 species. Planta, 1980, 149(1): 78-90. |
| [13] | Cai S Q, Xu D Q. Relationship between CO2 compensation point and photorespiration in soybean leaves. Acta Phytophysiologica Sinica, 2000, 26(6): 545-550. |
| [14] | Dong Z X, Han Q F, Jia Z K, Ren G X. Photosynthesis rate in response to light intensity and CO2 concentration in different alfalfa varieties. Acta Ecologica Sinica, 2007, 27(6): 2272-2278. |
| [15] | Xu D Q. The Efficiency of Photosynthesis. Shanghai: Shanghai Science and Technology Press, 2002. |
| [16] | Walker B, Steffen W. An overview of the implications of global change for natural and managed terrestrial ecosystem. Conservation Ecology, 1997, 1(2): 2-20. |
| [17] | Drake B G, Gonzlez-Meler M A, Long S P. More efficient plants, a consequence of rising atmospheric CO2. Annual Review of Plant Physiology and Plant Molecular Biology, 1997, 48(1): 609-639. |
| [18] | Larcher H, Stuart Chapin F, Pons T L. Physiological Plant Ecology. Springer-Verlag New York Inc.; 2nd ed. 2008. |
| [19] | Tartachnyk I I, Blanke M M. Effect of delayed fruit harvest on photosynthesis, transpiration and nutrient remobilization of apple leaves. New Phytologist, 2004, 164(3): 441-450. |
| [20] | Zhang X H, Liu N, Guo J M. Comparative study of photosynthetic responses to short-term elevated CO2 in two Myrica species. Guizhou Science, 2006, 24(2): 71-74. |
| [21] | Liang X, Zhang L Q, Zhao G Q. A comparison of photosynthetic characteristics between Spartina alterniflora and Phragmites australis under different CO2 concentrations. Acta Ecologica Sinica, 2006, 26(3): 842-848. |
| [22] | Zhou H H, Chen Y N, Li W H, Chen Y P. Photosynthesis of Populus Euphratica olive and its response to CO2 concentration and high temperature in arid environment. Acta Ecologica Sinica, 2009, 29(6): 2797-2810. |
| [23] | Cure J D, Acock B. Crop responses to carbon dioxide doubling: A literature survey. Agricultural and Forest Meteorology, 1986, 38(1/3): 127-145. |
| [24] | Rowland-Bamford A J, Backer J T, Allen L H, Bowes G. Acclimation of rice to changing atmospheric carbon dioxide concentration. Plant, Cell and Environment, 1991, 14(6): 577-583. |
| [25] | Tang J J, Xu L M, Chen X, Hu S J. Interaction between C4 barnyard grass and C3 upland rice under elevated CO2: Impact of mycorrhizae. Acta Oecologica, 2009, 35(2): 227-235. |
| [26] | Kimball B A, Kobayashi K, Bindi M. Responses of agricultural crops to flee-air CO2 enrichment. Advances in Agronomy, 2002, 77: 293-368. |
| [27] | Weiss I, Mizrahi Y, Raveh E. Effect of elevated CO2 on vegetative and reproductive growth characteristics of the CAM plants Hylocereus undatus and Selenicereus megalanthus. Scientia Horticulturae, 2010, 123(4): 531-536. |
| [28] | Jiang Y P, Cheng F, Zhou Y H, Xia X J, Shi K, Yu J Q. Interactive effects of CO2 enrichment and brassinosteroid on CO2 assimilation and photosynthetic electron transport in Cucumis sativus. Environmental and Experimental Botany, 2012, 75: 98-106. |
| [29] | Jensen A M, Löf M, Gardiner E S. Effects of above-and below-ground competition from shrubs on photosynthesis, transpiration and growth in Quercus robur L. seedlings. Environmental and Experimental Botany, 2011, 71(3): 367-375. |
| [30] | Pfanz H, Vodnik D, Wittmann C, Aschan G, Batic F, Turk B, Macek I. Photosynthetic performance (CO2-compensation point, carboxylation efficiency, and net photosynthesis) of timothy grass (Phleum pratense L.) is affected by elevated carbon dioxide in post-volcanic mofette areas. Environmental and Experimental Botany, 2007, 61(1): 41-48. |
| [31] | Peterhansel C, Offermann S. Re-engineering of carbon fixation in plants-challenges for plant biotechnology to improve yields in a high-CO2 world. Current Opinion in Biotechnology, 2012, 23(2): 204-208. |
| [32] | Zouzoulas D, Koutroubas S D, Vassiliou G, Vardavakis E. Effects of ozone fumigation on cotton (Gossypium hirsutum L.) morphology, anatomy, physiology, yield and qualitative characteristics of fibers. Environmental and Experimental Botany, 2009, 67(1): 293-303. |
| [33] | Farazdaghi H. The single-process biochemical reaction of Rubisco: A unified theory and model with the effects of irradiance, CO2 and rate-limiting step on the kinetics of C3 and C4 photosynthesis from gas exchange. Biosystems, 2011, 103(2): 265-284. |
| [34] | Kozaki A, Takeka G. Photorespiration protects C3 plants from photooxidation. Nature, 1999, 384(6609): 557-560. |
| [35] | Wingler A, Quick W P, Bungard R A, Bailey K J, Lea P J, Leegood R C. The role of photorespiration during drought stress: an analysis utilizing barley mutants with reduced activities of photorespiratory enzymes. Plant, Cell and Environment, 1999, 22(4): 361-373. |
| [36] | Haupt-Herring S, Fock H P. Exchange of oxygen and its role in energy dissipation during drought stress in tomato plants. Physiologia Plantarum, 2000, 110(4): 489-495. |
| [37] | Noguès S, Alogre L. An increase in water deficit has no impact on the photosynthetic capacity of field-grown Mediterranean plants. Functional Plant Biology, 2002, 29(5): 621-630. |
| [3] | 肖国举, 张强, 王静. 全球气候变化对农业生态系统的影响研究进展. 应用生态学报, 2007, 18(8): 1877-1885. |
| [9] | 张绪成, 于显枫, 马一凡, 上官周平. 高大气CO2浓度下小麦旗叶光合能量利用对氮素和光强的响应. 生态学报, 2011, 31(4): 1046-1057. |
| [10] | 周先容, 汪建华, 张红, 王进闯, 张远彬. CO2 浓度升高和模拟氮沉降对青川箭竹叶营养质量的影响. 生态学报, 2012, 32(24): 7644-7653. |
| [13] | 蔡时青, 许大全. 大豆叶片CO2补偿点和光呼吸的关系. 植物生理学报, 2000, 26(6): 545-550. |
| [14] | 董志新, 韩清芳, 贾志宽, 任广鑫. 不同苜蓿(Medicago sativaL.) 品种光合速率对光和CO2 浓度的响应特征. 生态学报,2007, 27(6): 2272-2278. |
| [15] | 许大全. 光合作用效率. 上海: 上海科学技术出版社, 2002. |
| [20] | 张喜焕, 刘宁, 郭建民. 杨梅属两种植物光合特性对CO2浓度升高响应的比较研究. 贵州科学, 2006, 24(2): 71-74. |
| [21] | 梁霞, 张利权, 赵广琦. 芦苇与外来植物互花米草在不同CO2浓度下的光合特性比较. 生态学报, 2006, 26(3): 842-848. |
| [22] | 周洪华, 陈亚宁, 李卫红, 陈亚鹏. 干旱区胡杨光合作用对高温和CO2浓度的响应. 生态学报, 2009, 29(6): 2797-2810. |