 2014, Vol. 34
2014, Vol. 34文章信息
- 张香凝, 孙向阳, 王保平, 乔杰, 崔令军, 孟伟, 杨超伟, 郝明丽
- ZHANG Xiangning, SUN Xiangyang, WANG Baoping, QIAO Jie, CUI Lingjun, MENG Wei, YANG Chaowei, HAO Mingli
- 土壤水分胁迫对拉瑞尔小枝水分参数的影响
- Water parameters of the branch of Larrea tridentata under different soil drought stress
- 生态学报, 2014, 34(8): 1984-1993
- Acta Ecologica Sinica, 2014, 34(8): 1984-1993
- http://dx.doi.org/10.5846/stxb201211261669
-
文章历史
- 收稿日期:2012-11-26
- 修订日期:2013-10-30






2. 北京林业大学水土保持学院, 教育部水土保持与荒漠化防治重点实验室, 北京 100083;
3. 国家林业局泡桐研究开发中心, 郑州 450003;
4. 开封县农业局, 开封 450000
2. Key Laboratory of Soil and Water Conservation & Desertification Combating of Ministry of Education, School of Soil and Water Conservation, Beijing Forestry University, 100083, China;
3. Paulownia Research and Development Center of China, State Forestry Administration, Zhengzhou 450003, China;
4. Kaifeng County Bureau of Agriculture, Kaifeng 450000, China
水分是树木生长的重要生态因子之一。我国约有52%的国土面积处于干旱、半干旱地区[1]。植物与水分的关系研究成为陆地生态系统对全球变化响应研究的热点问题[2, 3, 4],也是植物生理生态学研究领域的重要课题之一[5, 6, 7, 8, 9, 10]。因此加强林木抗旱性研究,探明树木对干旱胁迫的响应及其机理,已经成为目前我国林业领域一个亟待解决的问题[11]。为了丰富适宜干旱、半干旱地区的造林树种,国家林业局从美国莫哈为沙漠地区引入了蒺藜科(zygophyllaceae) 一个古老的灌木树种拉瑞尔(Larrea tridentata)[12],目前L. tridentata已在河南原阳生长了8a。
Larrea tridentata是亚热带沙漠中一种常绿含树脂的灌木,有很高的生态价值和药用价值。Larrea tridentata在年降雨量548.5 mm至50 mm的地方都能生长,但其主要生长地为美国德克萨斯州的南部和西部,直至新墨西哥州南部,海拔约1500m[12]的干旱地区[13]。国外已有研究主要集中在高温、干旱以及CO2浓度升高等对其生理生态特性的影响以及药用价值的研究。已有研究表明,L. tridentata保持膨压的能力很强,特别是通过细胞壁弹性维持膨压的能力很强[14],能在极端干旱的情况下保持代谢活动及生理活性[15, 16, 17, 18]。在Chihuahuan沙漠,当一般植物都已经呈现出脱水状态时,Larrea tridentata小枝黎明前水势还能达到-10 MPa[19]。气孔调节是L. tridentata 应对短期CO2浓度提高和干旱的主要措施[20]。CO2浓度的提高有利于干旱情况下L. tridentata保持小枝水势[21](CO2浓度从550提高的700 mol/mol,正午小枝水势从-4.6 MPa升到-3.4 MPa)。
但植物的耐旱性毕竟是一个非常复杂的复合性状。它不仅受到植物本身遗传因素的控制,也受到外界自然环境条件的影响,植物在相同时期处在不同生境条件下的耐旱性和水分亏缺的敏感程度也不同[22]。鉴于此,本文意在前些年对引种到中国的L. tridentata光合、酶活性等耐旱生理生态特性研究结果的基础上,借助PV曲线技术,测得多个水分指标如植物内部组织的水分组成、潜在的组织最大渗透势和对水分胁迫的反馈调节能力等[23],对其种植到中国境内后水分参数与原产地的差异进行研究,更全面的探讨其水分利用特性。为L. tridentata在我国的推广种植提供更全面的理论依据。
1 研究地概况和研究方法 1.1 研究地概况试验地位于河南省中部的原阳县境内——国家林业局泡桐研究开发中心实验基地,地理位置为东经113°34′—113°52′,北纬34°53′—35°05′之间。属暖温带大陆性季风气候区。地处黄河故道,为典型平原地区,区内土壤为黄河冲积风沙土,沙粒较细,土层深厚,透气性强。土质pH值7.5—8.5,养分含量较低,有机质、有效氮、有效磷平均含量分别为8.79%、20.53 mg/kg、2.75 mg/kg。
1.2 小枝水势测定方法选取的L. tridentata为1年生未经过移栽的苗木。平均苗高为30—50 cm,冠幅40—60 cm。从2005年8月3日开始,自然干旱法,选择晴朗无云的天气,每隔1 d测1次水势日变化及PV曲线,直到8月19日下雨为止。从中选取4d有代表性土壤水分条件(8月3日,15日,16日,18日)下测定的数据。水势的测定用压力室法[24] (每次测定时取3个不同植株的位于L. tridentata中上部的小枝测定后取平均值),测定时间为6:00。水势日变化从6:00开始,每隔2 h测1次,直到20:00。用6:00的水势值来表示当天的小枝水势[25]。PV曲线每隔2 d测1次,于黎明时从供试苗木中上部截取10 cm左右的小枝,立即用JA5003A电子天平(精度0.0001g)称其鲜重,然后将小枝基部插入盛有清水的烧杯中,上覆塑料袋保持24 h,让小枝饱和吸水;将已恢复到近饱和的叶片迅速装入压力室钢筒,用Hammel法分次测定压出的水量,最后取出样品时再测定鲜重,并于75 ℃下烘干至恒重,然后以各次平衡压的倒数为纵坐标,相应的水分亏缺为横坐标,绘制PV曲线[24]。借助PV曲线计算出每个供试小枝的有关水分参数。由于PV曲线重复性好,每个处理仅测1次即可[26, 27]。
1.3 土壤水分状况测定方法土壤含水量的测定用烘干法,每隔1 d测1次,测定时间是每天10:00。根据2005年12月进行的根系调查可知,L. tridentata的主要根系分布于20—60 cm的土壤中,采集20—40、40—60 cm 2个层次的土壤,每个层次取3个重复,计算其平均值即为该层土壤的质量含水量。同时做土壤水分特征曲线(图 1),测定时用离心机法。通过土壤水分特征曲线计算出该含水量下的土壤水势值。用20—60 cm这2个层次土壤水势的平均值来描述该时期的土壤水分状况。
选取4种土壤水分梯度处理,土壤水势分别为-0.021 MPa(A处理)、 -0.121 MPa(B处理)、 -0.698 MPa(C处理)、 -0.968 MPa(D处理)。根据土壤水有效性的划分标准[28]可知处理A土壤属于田间持水量状态,B属于半干旱状态,C、D处理已经接近凋萎系数,属于极度干旱状态,(该地区土壤田间持水量为:-0.01—-0.03 MPa,凋萎系数为-1.5 MPa)。

|
| 图 1 土壤水分特征曲线 Fig. 1 Soil water characteristic curve |
判断L. tridentata耐旱机理可用李吉跃的方法[25],如果已经测得某一树种苗木叶水势与土壤水势的关系,可以用Fuzzy数学中的择近原则来解决。即,设Ai为各类中心模型,B为待判树种的关系方程,Ai,B∈F u ,i=1,2,3,4。
N ·,· 为Ai与B的贴近度,若存在i0,使得N(Ai0,B)=max{N(A1,B),N(A2,B),N(A3,B),N(A4,B)}则判定B属于Ai0类。基于这一原则,采用Euclid贴近度来构造判别模型。其基本模型为:

u= u1,u2,…,un ,依据基本模型可建立如下判别模型。
基于Ψwp与Ψws关系(y=axb)的判别模型如下(ψws∈{0.1,0.3,0.5,1.0,1.5,2.0,2.5,3.0,3.5,4.0,4.5,5.0})。
高水势延迟脱水耐旱树种判别模型:
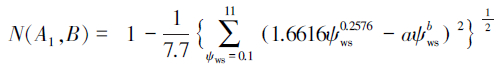
亚高水势延迟脱水耐旱树种判别模型:
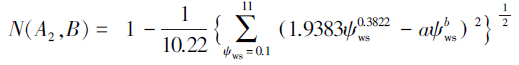
低水势忍耐脱水耐旱树种判别模型:
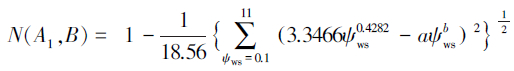
亚低水势忍耐脱水耐旱树种判别模型:
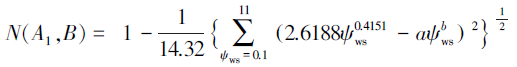
L. tridentata日变化测定都是在晴朗无云的天气下进行,由图 2看出,水势的最高点出现在6:00,然后逐渐降低,16:00以后开始回升,但是晚间的水势还是比清晨的水势略低,这与大多数研究相似[25]。

|
| 图 2 水势日变化 Fig. 2 Diurnal changes of water potential of L. tridentata |
在土壤水分状况较好的情况下,L. tridentata水势的日变化呈双峰曲线(处理A、B),水势的最低点出现在12:00,14:00时稍有升高,在16:00时又出现一个峰值。水势的日变幅(最高水势与最低水势之差)较大为2.23、2.03。随着土壤水分胁迫的逐渐加重(处理C、D),水势日变化的峰值出现提早,双峰不明显。水势日变幅减小为1.7、1.35。可能是因为随着土壤含水量的继续下降,植物组织水分含量减少,植物体内水流阻力增加,使白天的小枝水势较早的达到最低值,其后维持在一个接近膨压为零的渗透势的水平上[29]。
2.2 不同土壤水势(Ψws)对清晨小枝水势(Ψwp)的影响植物黎明前的水势是土壤水分有效性、甚至是某些植物根系活力的良好标志,已有研究表明,随着土壤水势的降低,苗木水势也随之降低,拟合关系以幂函数方程最好y=axb (0<b<1),式中,y为小枝水势,x为土壤水势[30]。
利用SPSS对L. tridentata土壤水势与小枝水势的关系进行拟合,结果符合已有的研究结果(图 3)。对拟合结果进行F检验,F=21.99,达到极显著水平。从图 3可以看出,在水分条件较好的情况下,小枝水势的变化比较平缓,随着土壤干旱胁迫的进一步加重,苗木水势急剧下降,以便在苗木水势和土壤水势之间形成较大的水势梯度,从而有利于苗木从土壤中吸收水分,这是苗木对土壤干旱胁迫的反应和适应。

|
| 图 3 土壤水势与L. tridentata小枝水势关系 Fig. 3 Relationship between Ψwp and Ψws of L. tridentata |
表 1为由PV曲线计算测得的小枝各水分参数值。
2.4 不同土壤水势(Ψws)对膨压(Ψp)与小枝水势(Ψwp)关系的影响在植物水势下降时,保持一定的膨压对维持植物的生长和生存以及正常的生理功能都至关重要。在不同土壤水分条件下,L. tridentata苗木膨压与小枝水势呈极显著直线线性关系,相关系数均在0.9以上,具体结果见表 2。这与其他许多研究的结果是一致的[25, 28, 31]。
| 处理 | Ψwp
/MPa | RWC
/% | RWC0
/% | ROWC0
/% | Ψ100π
/MPa) | Ψ0π
/MPa | Va
/% | Vs
/% | Va/ Vs | εmax
/MPa |
| Ψwp为小枝水势;Ψ100π为饱和含水量时的最大渗透势;Ψ0π膨压为零时的渗透势;εmax为最大体积弹性模量;Vs自由水含量;Va束缚水含量;RWC0膨压为零时的相对含水量;ROWC0膨压为零时的渗透水相对含量;Ψp为膨压 | ||||||||||
| A | -0.15 | 98.85 | 58.91 | 33.86 | -0.21 | -1.7 | 53.59 | 46.26 | 1.12 | 10.13 |
| B | -0.35 | 98.41 | 83.55 | 26.2 | -0.49 | -1.9 | 76.71 | 21.7 | 3.54 | 9.48 |
| C | -0.78 | 92.55 | 60.48 | 13.12 | -0.75 | -2.6 | 36.57 | 55.98 | 0.65 | 2.37 |
| D | -1.13 | 87.02 | 76.25 | 28.19 | -1.01 | -2.29 | 43.7 | 43.32 | 1.01 | 2.27 |
|
处理
Treatment | 膨压与水势的相关方程
Dependent equation of water potential and turgor | R2 |
| A | y=0.4055x+0.2014 | 0.8165 |
| B | y=0.6137x+0.2385 | 0.9803 |
| C | y=0.198x+0.6754 | 0.8429 |
| D | y=0.3161x+0.7228 | 0.8009 |
根据清晨苗木的小枝水势来判断植物的水分胁迫状态,轻度水分胁迫(ψwp:-0.5—-1.0MPa)、中度水分胁迫(ψwp:-1.0—-1.5MPa)、严重水分胁迫(ψwp:< -1.6MPa)[25]。从图 1可以看出在A、B处理时L. tridentata植株体未表现出受到干旱胁迫的特征,小枝水分状况良好,在处理C时,小枝水分参数显示已受到到轻度胁迫,在处理D时达到中度胁迫。但是处理B的土壤水分状态已经达到了半干旱,到处理C时土壤水分含量已低于萎蔫系数。可见L. tridentata小枝水势出现的胁迫状态要滞后于土壤出现的水分胁迫状态,并且无论土壤水分胁迫状态如何,L. tridentata的水势最低值基本保持不变,L. tridentata能在土壤水分胁迫时保持良好的水分状况,表现出了很强的抗旱性,有高水势耐旱植物的生理特征。
从土壤水势与小枝水势拟合模型来看,土壤水势与小枝水势(y=axb (0< b<1))的相关关系显著,模型为y=1.0541x0.5033,拟合方程中的a、b值越大,表明苗木叶水势随土壤水势变化的幅度(下降幅度)越大,土壤与植物之间形成的水势梯度越大,苗木保持水分吸收的能力越强,耐旱性就越差[25]。张建国、李吉跃等测的北方几种耐旱性较强树种的Ψwp与Ψws的关系方程中的a,b值如下[28],沙枣(Elaeaanus angustifolia)(2.564,0.3509),新疆杨(Populus bolleana Lauche)(2.4900,0.5100),文冠果(Xanthoceras sorbifolia)(2.652,0.3053),火炬树(Rhus typhina L.)(1.6853,0.4777),长白松(Pinus Sylverstis var.Sylvestriformis)(1.553,0.3710),华北落叶松(Larix principis-rupprechtii)(1.9539,0.3534),油松(Pinus tabulaeformis)(河北平原)(1.679,0.9184),油松(甘肃)(1.897,0.2059),油松(河北遵化)(1.387,0.2321),樟子松(Pinus sylvestris)(1.6301,0.2817)。将图 2所得的方程与上述值进行比较发现,L. tridentata的a、b值都很小,说明L. tridentata小枝水势随着土壤水势下降的幅度较小。其中a值比上述张建国等测得的大多针叶树种都小,而针叶树由于叶组织构造不同,组织抗脱水的能力本身就比阔叶树强,说明在相同的土壤水分状况下,L. tridentata更容易保持较高的水势,叶片保水能力与针叶树接近。b值也较小,说明随着Ψws的下降的Ψwp变化较小,保持膨压的效果越好,组织能维持较好的水分状态。而作为阔叶树的 L. tridentata在这方面表现出了的抗旱性比针叶树都强。说明在相同的土壤水分状况下,L. tridentata与其他常见的阔叶树种相比更容易保持较高的水势,维持较好的水分状态,其组织抗脱水的能力接近针叶树。随着土壤水分含量的下降,L. tridentata苗木小枝水势下降的幅度逐渐减小,一方面由于土壤水分有效性降低,植物体内水分以束缚水为主,传导力下降,使得水势下降幅度降低,另一方面,苗木器官受到环境影响而减弱功能,而生物体内部具有恢复该器官的反应和能力,如苗木受到干旱胁迫时通过气孔运动及渗透调节等对干旱胁迫的反应和适应,使得苗木小枝水势下降的速度随之减弱。这是一种生物补偿机制,该补偿机制是生物保存自身的一种重要机能[32]。因而从这方面表现出L. tridentata的对干旱具有较强和较快的适应性。
3.2 不同土壤水势(Ψws)对L. tridentata细胞最大弹性模量εmax)的影响εmax反映了细胞膨压随体积而变化的速率。εmax值越小,显示出细胞有比较大的弹性,因而保持膨压的能力也较强[25, 27]。当苗木受到土壤干旱胁迫的影响时,不同树种的εmax值都会发生变化。由PV曲线测得的数值(表 1)可以看出,随着土壤水分含量的逐渐减少L. tridentata的εmax随之下降,从B处理到C处理,εmax由9.48下降到2.37下降了75%左右。当土壤水分状况达到极干旱状况(即Ψws在-1.5 MPa以下)时,εmax为2.27,比国内测得几种较耐旱的树种侧柏(Platycladus orientalis)(4.59)[27]、柠条(Caragana Korshinskii Kom.)(11.98)[33]、青海云杉(Picea crassifolia Kom.)(3.53)[34]在类似的土壤水分状况下都要低的多。说明L. tridentata的细胞柔软而富有弹性,对干旱胁迫反映很快。这不但有利于L. tridentata在低水势下保持正膨压,更有利于原生质忍耐脱水而在负膨压下生存[27]。
3.3 不同土壤水势(Ψws)对Ψ0π和Ψ100n的影响Ψ0π(膨压为零时的渗透势)反映了树种保持膨压能力的大小,Ψ0π值越低,则在水分胁迫条件下植物体的膨压越易维持,对干旱的忍耐性也就越强[34]。Ψ0π反映了树木维持最低膨压的极限渗透势,是植物生命活动发生质变的一个临界指标[35]。Ψ100n(组织充分膨胀时的渗透势)也反映苗木保持最大膨压的能力,Ψ100n值越小,苗木维持膨压的能力也就越强[36],则越有利于植物在水分胁迫条件下继续保持细胞伸长生长[34]。从表 1可以看出,随着土壤干旱胁迫的发展Ψ100n和Ψ0π都逐渐减小,降幅分别为380.95%、34.71%,比类似土壤水分状况下山梨(131.4%,37.16%)[37]、青海云杉(9.59%,6.58%)[34]、侧柏(10.12%,4.91%)[26]这些较为耐旱树种的降幅都要大。反映出L. tridentata组织在失去较多水时才发生质壁分离,通过渗透调节维持膨压的能力也较强,有利于其叶片在干旱胁迫下保持膨压,推迟萎蔫进程,避免造成不可逆的伤害,这与F. C. Meinzer的研究结果一致[20]。
3.4 不同土壤水势(Ψws)对束缚水含量(Va)以及束自比(Va/ Vs)的影响渗透调节的另一种方式就是通过细胞内溶质的增加或束缚水含量的变化,亦即束自比值的大小来实现的。束缚水含量是一个重要的抗旱指标,通常认为束缚水比例越大,抗旱性就越强。并且Va/ Vs值越高,则越有利于植物吸水增强耐旱性。在干旱情况下,有一些树种束缚水含量不高,但可通过束缚水含量的增加来进行渗透调节的,如茶条槭、山桃、山梨[37],而另一些树种则是因为自身束缚水含量较高,通过细胞内溶质增加来进行渗透调节的[38, 39],L. tridentata就属于这类树种。
L.tridentata的束缚水含量是36.57%—76.71%,在类似土壤水分条件下,油松的束缚水含量为31.34%—41.17%[27]、侧柏为22.16%—26.16%[27]、山荆子(Malus baccata)40.69%—72.09%[37],治沙旱生植物白柠条34.28%—48.77%[40]、花棒(Hedysarum scoparium Fisch.)29.91%—52.79%[41],显然,L.tridentata的束缚水含量比它们都大,并且始终大于自由水的比例,其原因可能是L. tridentata细胞原生质粘滞性及原生质胶体亲水性更强,吸水能力更大[27, 34]。
L.tridentata的Va/Vs值(0.65%—3.73%)与一般用于治沙的旱生植物白柠条的Va/Vs值1.18—3.25、花棒Va/Vs值0.76—3.73差不多[31]。从表 1可以看出在土壤轻度水分胁迫下Va/Vs值有所提高,但是随着干旱程度的加重,Va/Vs值反而降低了,说明随着干旱胁迫的加重,通过提高Va/Vs值来进行渗透调节的功能逐渐消失。L.tridentata不是通过提高束缚水比例而是通过细胞溶质的积累来实现渗透调节。
3.5 不同土壤水势(Ψws)对RWC0和ROWC0的影响RWC0和ROWC0是组织细胞初始质壁分离时的相对含水量和相对渗透水含量。RWC0和ROWC0值越低,表明细胞在很低的含水量下才发生质壁分离,忍耐脱水的能力就越强。所以这两个指标在一定程度上反映了组织细胞对脱水的忍耐能力的大小[25]。
L. tridentata的RWC0(83.55%—58.91%) ROWC0(13.12%—33.86%)随着土壤含水量的降低,RWC0的含量呈上升趋势,ROWC0的含量呈下降趋势,表明随着土壤干旱胁迫的发展,L. tridentata原生质忍耐脱水能力提高,细胞在很低的含水量下才发生质壁分离,由细胞壁特性所反映的渗透调节能力增强。
3.6 不同土壤水势(Ψws)对膨压(Ψp)与小枝水势(Ψwp)关系的影响直线方程(Ψp =a+bΨwp)中a表示苗木充分膨胀时的最大膨压[26]。b值表示随叶水势下降,膨压下降的速度,能反映植物保持膨压能力的大小[27]。b值越小,表明随水势的下降,膨压下降的速率越小,因而树木保持膨压的能力也越强。但是a值的下降则不利于苗木的生长[26]。L. tridentata苗木小枝水势在处理A、B下处于无水分胁迫的状态,处理C时达到轻度水分胁迫状态,处理D时达到中度水分胁迫状态。从表 3可以看出,随着土壤水分含量的减少,在土壤处于轻度水分胁迫的状态下,L. tridentata的b值呈减小的趋势,a值呈增大趋势。说明在土壤轻度干旱胁迫下L. tridentata维持膨压的能力较强,苗木保持最大膨压的能力也得到加强。从处理C到处理D,b值和a值都增加,表明,随着土壤干旱胁迫的加重,L. tridentata维持膨压的能力有所减弱,但是苗木保持最大膨压的能力依然很强,能维持苗木的正常生长。
3.7 耐旱性综合评价鉴于苗木的耐旱性受到多个水分参数的影响,用不同的水分参数对其进行评价得出的结论不同,为了作出准确的评价,需要从多个角度,采用多个指标,综合评价[36]。目前,综合评定植物抗旱性的方法主要有两种,一种是张建国等,利用模糊数学反隶属函数进行定量转换,得出模糊综合评判值(Fuzzy value)来评定植物的抗旱性;另一种是王孟本[29]创立了用DI综合抗旱指数(DI index)的求算植物抗旱性的方法。两种方法效果相同,这里我们采用第一种方法[41, 42]。选取经过不同干旱处理后的L. tridentata的6种水分参数(εmax、Ψ0n、Ψ100n、RWC0、ROWC0和膨压与水势关系方程中的b值)进行隶属函数定量转换评价经过不同干旱处理后的L. tridentata的耐旱能力。这6个值的共同点是:水分参数数值越小,则越有利于树木保持膨压,增强其耐旱性。6个评价指标与耐旱性呈负相关,所以用反隶属函数来计算每一个指标的耐旱隶属函数值
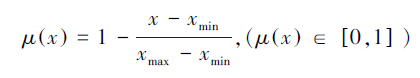
然后把4个处理的6个指标的耐旱隶属函数值累加,求平均值,平均值越大,耐旱性越强。
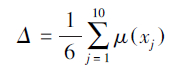
综合评价的结果见表 3。
|
处理
Treatment | 耐旱指标隶属函数值
Drought- enduringsubordinate function values | 耐旱指数排序
Order of drought- enduring index | 综合耐旱指数
Drought- enduring index | |||||
| μ(RWC0) | μ(Ψ0π) | μ(Ψ100π) | μ(εmax) | μ(ROWC0) | μ(b) | |||
| A | 1 | 0 | 0 | 0 | 0 | 0.5 | 0.25 | 3 |
| B | 0 | 0.35 | 0.222 | 0.083 | 0.369 | 0 | 0.17 | 4 |
| C | 0.936 | 0.675 | 1 | 0.987 | 1 | 1 | 0.933 | 1 |
| D | 0.296 | 1 | 0.656 | 1 | 0.273 | 0.716 | 0.657 | 2 |
可以看出,处理C时L. tridentata的耐旱能力最强,其次是处理D,在土壤水分条件较好及轻度水分胁迫下,L. tridentata表现出的耐旱性相对较弱。因为在土壤水分良好的条件下,土壤中的水分主要是易被根系吸收利用的自由水,此时苗木蒸腾作用较强,耗水较大,土壤水分含量下降初期,土壤中有效水分耗失较快,使得水分的有效性大为下降,传导力下下降,苗木叶片及气孔运动等一些对干旱的反应和适应机制还未完全启动,因而苗木的水势变化较大,此时苗木表现的抗旱能力最弱。经过适度干旱处理后在L. tridentata的耐旱性有所提高,表现出很强的耐旱生理特征,随着土壤干旱程度的加重和干旱胁迫时间的加长,L. tridentata的耐旱性稍有降低,但仍保持在较高的水平。
3.8 树种耐旱机理的判定Turner[43]对植物耐旱机理做了归纳,分为避旱型和耐旱型两大类。避旱型是指植物在严重干旱胁迫之前就完成生命周期,L. tridentata是多年生的灌木很显然不属于这种类型;耐旱型又分为两种高水势延迟脱水和低水势忍耐脱水两种耐旱机理[25]。
对于本实验测得的a=0.5607,b=0.3642,代入上述判别模型得:N(A1,B)=0.4415,N(A2,B)=0.3685,N(A3,B)=0.2577,N(A4,B)=0.2984。显然,N(A1,B)最大。根据判别模型得出的结论是一年生L. tridentata属于高水势延迟脱水耐旱机理。
综上所述,L. tridentata在所设计的土壤水分梯度内,随土壤水势的下降,均能在一定程度上提高自身的渗透调节和维持膨压的能力。渗透调节能力主要是通过拥有较高的束缚水含量和细胞内溶质增加来进行的;维持膨压能力主要是靠改变细胞壁弹性。随着土壤干旱胁迫的发展,L. tridentata原生质忍耐脱水能力得到提高,细胞在很低的含水量下才发生质壁分离。这些与国外的研究结果完全一致[15, 16, 17, 18, 20],说明现引种在中国河南生长的L. tridentata保存了在原产地生长时拥有的优良耐旱特性。
本实验中,经过适度的干旱处理,L. tridentata的耐旱性有显著提高,在土壤水势为-0.698 MPa时最强。L. tridentata的抗旱性较强,它的耐旱机理主要是高水势延迟脱水耐旱机理,即通过限制水分丧失或通过水分吸收(深广的根系)来延迟脱水的发生,保持较高的组织水势;同时也兼有部分低水势忍耐脱水耐旱机理,即原生质及其主要器官在严重脱水的情况下伤害很轻或基本不受伤害,细胞壁弹性大,能在严重水分胁迫下维持生长。
由于本次研究由于实验设备所限制,并没有涉及苗木根部水势变化,若能同时研究苗木的各个部位在干旱胁迫条件下的水势梯度变化,就更能全面的评价苗木的吸水能力和调节体内水分平衡能力。另外,单一的植物水势值不能全面反映植物的水分状况以及抗旱特性。因此,一方面应继续探索和完善职务水势的基础理论研究,另一方面应该从SPAC入手,分析各个组分水势的变换及其相关性以及水分在植物体内的传输过程及其调控机理。
| [1] | Guo L S, Tian Y L. Study on drought-resistance evaluation of common afforestation species in North China by PV technique. Journal of Inner Mongolia Forestry College, 1998, 20(3): 1-8. |
| [2] | Jiang G M. Review on some hot top ICS towards the researches in the field of plant physioecology. Acta Phytoecologica Sinica, 2001, 25(5): 514-519. |
| [3] | Yuan Y H, Deng X P. Effect of deficit and rewatering on wheat photosynthesis and yield. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(7): 1250-1254. |
| [4] | Lin X L, Xu Z Z, Wang Y H, Zhou G S. Modeling the responses of leaf photosynthetic parameters of Leymus chinensis to drought and rewatering. Acta Ecologica Sinica, 2008, 28(10): 4718-4724. |
| [5] | Dewar R C, Medlyn B E, Mcmutrie R E. Acclimation of the respiration and photosynthesis ratio to temperature: insights from a model. Global Change Biology, 1999, 5(5): 615-622. |
| [6] | Islam M R, Hamid A, Karim M A, Haque M M, Khaliq Q A, Abdul K Q, Ahmed J U. Gas exchanges and yield responses of mungbean (Vigna radiata (L.) Wilczek) genotypes differing in flooding tolerance. Acta Physiologiae Plantarum, 2008, 30(5): 697-707. |
| [7] | Sofo A, Dichio B, Montanaro G, Xiloyannis C. Photosynthetic performance and light response of two olive cultivars under different water and light regimes. Photosynthetica, 2009, 47(4): 602-608. |
| [8] | Montanaro G, Dichio B, Xiloyannis C. Shade mitigates photoinhibition and enhances water use efficiency in kiwifruit under drought. Photosynthetica, 2009, 47(3): 363-371. |
| [9] | Zhang S Y, Zhou Z F, Zhang G C, Wang M J, Zhan H X. Gas exchange characteristics of natural secondary shrubs prunus davidiana and prunus sibirica under different water stresses. Acta Botanica Boreali-Occidentalia Sinica, 2008, 28(12): 2492-2499. |
| [10] | Bu L D, Zhang R H, Chang Y, Xue J Q, Han M M. Response of photosynthetic characteristics to water stress of maize leaf in seeding. Acta Ecologica Sinica, 2010, 30(5): 1184-1191. |
| [11] | Wang D, Yao J, Yang X, Xue J H. Changes of leaf water potential and water absorption potential capacities of six kind of seedlings in Karst mount area under different drought stress intensities: Taking six forestation seedlings in karst mountainous region for example. Acta Ecologica Sinica, 2011, 31(8): 2216-2226. |
| [12] | Barbour M G. Germination requirements of the desert shrub Larrea Divaricata. Ecology, 1968, 49(5): 915-923. |
| [13] | Beatley J C. Effects of rainfall and temperature on the distribution and behavior of Larrea tridentata (Creosote-bush) in the Mojave desert of Nevada. Ecology, 1974, 55(2): 245-261. |
| [14] | Meinzer F C, Sharifi M R, Nilsen E T, Rundel P W. Effects of manipulation of water and nitrogen regime on the water relations of the desert shrub Larrea tridentate. Oecologia, 1988, 77(4): 480-486. |
| [15] | Ochel W C, Strain B R, Odening W R. Tissue water potential, photosynthesis, C-labeled photosynthate utilization, and growth in the desert shrub Larrea divaricata cav. Ecological Monographs, 1972, 42(2): 127-141. |
| [16] | Mooney H A, Björkman O, Collatz C J. Photosynthetic acclimation to temperature in the desert shrub, Larrea divaricata. I. Carbon dioxide exchange characteristics of intact leaves. Plant Physiology, 1978, 61(3): 406-410. |
| [17] | Giardi M T, Cona A, Geiken B, Kucera T, Masojidek J, Mattoo A K. Long-term drought stress induces structural and functional reorganization of photosystem Ⅱ. Planta, 1996, 199(1): 118-125. |
| [18] | Hamerlynck E P, Knapp A K. Photosynthetic and stomatal responses to high temperature and light in two oaks at the western limit of their range. Tree Physiology, 1996, 16(6): 557-565. |
| [19] | Cunningham G L, Burk J H. The effect of carbonate deposition layers (‘caliche’) on the water status of Larrea divaricata. American Midland Naturalist, 1973, 90(2): 474-480. |
| [20] | Schulze E D. Carbon dioxide and water vapor exchange in response to drought in the atmosphere and in the soil. Annual Review of Plant Physiology, 1986, 37: 247-274. |
| [21] | Erik P H, Travis E H, Michael E L, Smith S D. Effects of extreme high temperature, drought and elevated CO2 on photosynthesis of Mojave desert evergreen shrub, Larrea tridentata. Plant Ecology, 2000, 148(2): 183-193. |
| [22] | Shi S L, Wang Y C, Zhou J H. The change of water parameters in Tetraena mongolica with season and habitat. Acta Ecologica Sinica, 208, 28(12): 6079-6089. |
| [23] | Xi B Y, Wang Y, Di N, Jia L M, Li G D, Huang X F, Gao Y Y. Effects of soil water potential on the growth and physiological characteristics of Populus tomentosa pulpwood plantation under subsurface drip irrigation. Acta Ecologica Sinica, 2012, 32(17): 5318-5329. |
| [24] | Wang W L. Using pressure chamber to study the water characteristics of plant. Plant Physiology Communications, 1984, (3): 52-57. |
| [25] | Zhang J G, Li J Y, Shen G F. Studies of the Drought Tolerance Characteristics and Mechanism in Trees. Beijing: China Forestry Publishing House, 2000. |
| [26] | Li J Y, Zhang J G, Jiang J P. A study on water parameters of plantations in mountain areas of west Beijing(Ⅱ). Journal of Beijing Forestry University, 1994, 16(2): 1-8. |
| [27] | Li J Y. An application of PV technique to study drought resistance of pinus tabulaeformis and platycladus orientalis seedlings. Journal of Beijing Forestry University, 1989, 11(1): 3-9. |
| [28] | Sun X Y. Soil Science. Beijing: China Forestry Publishing House, 2005: 145-156. |
| [29] | Li H J, Wang M B, Chai B F. A study on water potential characteristics in four tree species in loess region. Bulletin of Botanical Research, 2001, 21(1): 100-105. |
| [30] | Li J Y. Mechanisms of drought tolerance in plants. Journal of Beijing Forestry University, 1991, 13(3): 92-97. |
| [31] | Yang M S, Pei B H, Yu D M. Influence of water stress on the abilities to maintain turgor and adjust osmosis in seedlings of hybrid clones of Populus tomentosa. Acta Ecologica Sinica, 1997, 17(4): 364-370. |
| [32] | Yuan B Z, Wang J, Zhao S L, Sun J. An approach to the mechanism of plant compensation. Chinese Journal of Ecology, 1998, 17(5): 45-49. |
| [33] | Feng J Z. Water characteristic curves and water relations of desert plants. Journal of Desert Research, 1995, 15(3): 222-227 |
| [34] | Dang H Z, Zhao Y S, Cen X W. Study on drought resistance of tree species on Plateau Hill in Gansu. Science of Soil and Water Conservation, 2003, 1(3): 21-25. |
| [35] | Li H T, Chen L Z. The hydro physiological characteristics of major plant species of the forest ecosystem in the warm temperate zone of northern China. Acta Phytoecologica Sinica, 1998, 22(3): 202-213. |
| [36] | Liu Y L. Plant Physiology in Water Adversity. Beijing: China Agriculture Publishing House, 1992: 139-141. |
| [37] | Sun Z H, Wang Q C. The drought resistance of four broad leaved species in the north of China with PV technique. Scientia Silvae Sinicae, 2003, 39(2): 33-38. |
| [38] | Li J Y. A Preliminary Study on drought resistance of pinus tabulaeforrnis and platycladus orientalis seedling. Journal of Beijing Forestry University, 1998, 10(2): 23-29. |
| [39] | Liu J P, Li Z J, Han L, Gong W J, Zhou L Z. Preliminary study on water parameters of Populus euphratica and P. pruinosa by using P-V curve. Acta Botanica Boreali-Occidentalia Sinica, 2004, 24(7): 1254-1259. |
| [40] | Fu Y R, Wang X Y, Li D M. A research on the Xerophytic from and physiologyical characteristic of sand-control plants. Bulletin of Soil and Water Conservation, 1994, 14(7): 53-58. |
| [41] | Zhang J G, Li J Y, Jiang J P. A study on water parameter of plantation in the westernmountain area of Beijing(Ⅰ). Journal of Beijing Forestry University, 1994, 16(1): 1-11. |
| [42] | Zang J G, Li J Y, Jiang J P. A study on water parameters of plantations in mountain areas of west Beijing(Ⅲ). Journal of Beijing Forestry University, 1994, 16(4): 46-54. |
| [43] | Turner N C. Adaptation to water deficits: A changing perspective. Plant Physiology, 1986, 13(1): 175-190. |
| [1] | 郭连生, 田有亮. 运用PV 技术对华北常见造林树种耐旱性评价的研究. 内蒙古林学院学报, 1998, 20(3): 1-8. |
| [2] | 蒋高明. 当前植物生理生态学研究的几个热点问题. 植物生态学报, 2001, 25(5): 514-519. |
| [3] | 袁永慧, 邓西平. 干旱与复水对小麦光合和产量的影响. 西北植物学报, 2004, 24(7): 1250-1254. |
| [4] | 林祥磊, 许振柱, 王玉辉, 周广胜. 羊草(Leymus chinensis) 叶片光合参数对干旱与复水的响应机理与模拟. 生态学报,2008, 28(10): 4718-4724. |
| [9] | 张淑勇, 周泽福, 张光灿, 王梦军, 战海霞. 水分胁迫下天然次生灌木山桃和山杏光合气体交换特征. 西北植物学报,2008, 28(12): 2492-2499. |
| [10] | 卜令铎, 张仁和, 常宇, 薛吉全, 韩苗苗. 苗期玉米叶片光合特性对水分胁迫的响应. 生态学报, 2010, 30(5): 1184-1191. |
| [11] | 王丁, 姚健, 杨雪, 薛建辉. 干旱胁迫条件下6 种喀斯特主要造林树种苗木叶片水势及吸水潜能变化. 生态学报2011, 31(8): 2216-2226. |
| [22] | 石松利, 王迎春, 周健华. 四合木(Tetraena mongolica)水分参数季节变化及生境间差异. 生态学报, 2008, 28 ( 12):6079-6089. |
| [23] | 席本野, 王烨, 邸楠, 贾黎明, 李广德, 黄祥丰, 高园园. 地下滴灌下土壤水势对毛白杨纸浆林生长及生理特性的影响.生态学报, 2012, 32(17): 5318-5329. |
| [24] | 王万里. 压力室(PRESSURE CHAMBER)在植物水分状况研究中的应用. 植物生理学通讯, 1984, (3): 52-57. |
| [25] | 张建国, 李吉跃, 沈国肪. 树木耐旱特征及其机理研究. 北京: 林业出版社, 2000. |
| [26] | 李吉跃, 张建国, 姜金璞. 京西山区人工林水分参数的研究(域). 北京林业大学学报, 1994, 16(2): 1-8. |
| [27] | 李吉跃. PV 技术在油松侧柏苗木抗旱特性研究中的应用. 北京林业大学学报, 1989, 11(1): 3-9. |
| [28] | 孙向阳. 土壤学. 北京: 中国林业出版社, 2005: 145-156. |
| [29] | 李洪建, 王孟本, 柴宝峰. 黄土区4 个树种水势特征的研究.植物研究, 2001, 21(1): 100-105. |
| [30] | 李吉跃. 植物耐旱性及其机理. 北京林业大学学报, 1991, 13(3): 92-97. |
| [31] | 杨敏生, 裴保华, 于冬梅. 水分胁迫对毛白杨杂种无性系苗木维持膨压和渗透调节能力的影响. 生态学报, 1997, 17(4):364-370. |
| [32] | 原保忠, 王静, 赵松岭, 孙劼. 植物补偿作用机制探讨. 生态学杂志, 1998, 17(5): 45-49. |
| [33] | 冯金朝. 沙生植物水分特征曲线及水分关系的初步研究. 中国沙漠, 1995, 15(3): 222-227. |
| [34] | 党宏忠, 赵雨森, 陈祥伟. 甘肃省高原山地树种的抗旱性研究. 中国水土保持科学, 2003, 1(3): 21-25. |
| [35] | 李海涛, 陈灵芝. 暖温带森林生态系统主要树种若干水分参数的季节变化. 植物生态学报, 1998, 22(3): 202-213. |
| [36] | 刘友良. 植物水分逆境生理. 北京: 农业出版社, 1992:139-141. |
| [37] | 孙志虎, 王庆成. 应用PV 技术对北方4 种阔叶树抗旱性的研究. 林业科学, 2003, 39(2): 33-38. |
| [38] | 李吉跃. 油松侧柏苗木抗旱特性初探. 北京林业大学学报,1988, 10(2): 23-29. |
| [39] | 刘建平, 李志军, 韩路, 龚卫江, 周正立. 胡杨、灰叶胡杨P-V曲线水分参数的初步研究. 西北植物学报, 2004, 24(7):1254-1259. |
| [40] | 符亚儒, 王晓云, 李冬梅. 治沙植物旱生结构及生理特性的研究. 水土保持通报, 1994, 14(7): 53-58. |
| [41] | 张建国, 李吉跃, 姜金璞. 京西山区人工林水分参数的研究(玉). 北京林业大学学报, 1994, 16(1): 1-11. |
| [42] | 张建国, 李吉跃, 姜金璞. 京西山区人工林水分参数的研究(芋). 北京林业大学学报, 1994, 16(4): 46-53. |