 2014, Vol. 34
2014, Vol. 34文章信息
- 阿米娜木·艾力, 常顺利, 张毓涛, 仇瑶, 何平
- Aminem ELI, CHANG Shunli, ZHANG Yutao, QIU Yao, HE Ping
- 天山云杉森林土壤有机碳沿海拔的分布规律及其影响因素
- Altitudinal distribution rule of Picea schrenkiana forest’s soil organic carbon and its influencing factors
- 生态学报, 2014, 34(7): 1626-1634
- Acta Ecologica Sinica, 2014, 34(7): 1626-1634
- http://dx.doi.org/10.5846/stxb201305311261
-
文章历史
- 收稿日期:2013-5-31
- 修订日期:2013-11-21






2. 新疆林科院森林生态研究所, 乌鲁木齐市 830063
2. Institute of Forest Ecology, Xinjiang Academy of Forestry, Urumqi 830063, China
土壤有机碳是全球气候变化的敏感指标[1]。有机碳库的动态变化可对生态系统主要功能、组成和发展方向产生显著的影响[2],与大气成分进行活性交换的土壤有机碳大约占到陆地生态系统碳的2/3[3],所以土壤有机碳的微小变化将会极大地缓和或加速大气CO2浓度的提高,进而改变全球碳循环[4]。
土壤有机碳来自植物分泌物及残体的归还,其储量主要受植被光合利用率、群落分布、水位波动规律[5],以及气候、土壤、人类活动等因素的影响[6, 7, 8]。因此,人类活动干扰以及外界环境条件的变化,均可对陆地生态系统土壤有机碳库产生影响[2]。在山地生态系统研究工作中,很难排除森林类型变化的影响,而只考虑水热梯度变化条件下土壤有机碳的空间分布规律。王海燕[9]等提出了气候变化影响下落叶松-云杉混交林土壤有机碳密度变化情况。王文凡[10]等提出同一山地不同植被类型土壤有机碳密度存在差异性。
天山是植被类型单一、结构简单的山地生态系统,土壤有机碳的变化受到气候条件与云杉纯林生长趋势的影响。目前针对于具有单一植被类型的山地生态系统进行土壤有机碳密度变化的研究还极少,本研究通过云杉纯林不同海拔平均胸径、树高的调查统计分析与水热配比关系结合在一起,综合考虑气候与单一植被类型云杉纯林的变化对土壤有机碳密度的影响,研究土壤有机碳密度沿海拔的分布规律。为正确评估天山土壤总有机碳密度变化提供一定的参考,并为该领域研究工作中排除植被的影响而只考虑气候对土壤有机碳变化的研究提供一定的科学依据。
1 材料与方法 1.1 样品采集地概况以中国森林生态系统研究网络(CFERN)天山森林生态系统定位站研究站(E87°07′—87°28′,N43°14′—43°26′)为依托进行样品采集。样品采集地位于天山北坡乌鲁木齐县板房沟林场,海拔1800—2800 m,属温带大陆性气候,年总辐射量达5.85×105 J/cm2 · a,年均气温约为2—3 ℃,历年极端最高温为30.5 ℃,极端最低温为-30.2 ℃,年降水量400—600 mm,最大积雪深度为65 cm,林下土壤为山地灰褐色森林土。该区植被类型以天山云杉(Picea Schrenkiana)纯林为主的温带针叶林,郁闭度介于0.4—0.8,其凋落物的分解是土壤中生成较厚的腐殖质的重要途径。主要地被植物有黑果小檗(Berberis heteropoda)、红果小檗(Berberis nummularia)、忍冬(Lonicera sp.)、天山花楸(Sorbus tianschanica)、黑果荀子(Cotoneastermelanocarpus)、蔷薇(Rosa sp.)、绣线菊(Spiraeahypericifolia)、老鹳草(Geranium dahuricum)、天山雨衣草(Alchemilla tianschanica)、羊角芹(Aegopodiumpodagraria)等[11]。
1.2 土样采集方法2011年7月,在水西沟采样区,按照一定的海拔梯度以及土壤颜色的变化情况进行采样区的划分。从云杉林海拔1800 m下限到2800 m上限,按每200 m一个海拔梯度,在每个梯度内选取了3块20 m×20 m的样地;在每块样地内,按照S型方法布设了3个土壤采样点,各挖1 m深土壤剖面,分5层取土样,共计取得225个土样;在每块样地内的3个土壤采样点中随机选取一个剖面进行分层环刀取样,分层计算土壤容重,并以此代表该样地的各层土壤容重。样品带回实验室风干后过2mm筛,保存备用。
1.3 样地调查方法在研究区从其森林分布的下限到上限,选取一条东西宽200 m的样带,以50 m垂直海拔梯度为间隔,设置20 m×20 m的固定样地44个,对应于各海拔梯度的样方数见表 1。对每个样方进行每木检尺,调查因子主要包括树高、胸径、冠幅、枝下高、病虫害状况等,按胸径、树高对调查数据进行分级统计,结果见表 1。
| 海拔 Elevation/m | 样地数 Plots | 株树 Trees | 各分级所占的比例The proportion of each grading/% | |||
| DBH<10 cm | 10—15 cm | 15—20 cm | >20 cm | |||
| 树高<10 m | 10—15 m | 15—20 m | >20 m | |||
| 横线上方为各胸径(DBH)径级分级所占比例(%),横线下方为各树高分级所占比例(%) | ||||||
| 1800—2000 | 8 | 285 | 36.14 | 21.41 | 15.43 | 27.02 |
| 38.87 | 13.07 | 29.68 | 18.38 | |||
| 2000—2200 | 9 | 331 | 22.96 | 22.96 | 22.05 | 32.03 |
| 23.56 | 21.15 | 33.84 | 21.45 | |||
| 2200—2400 | 10 | 236 | 25 | 19.49 | 17.8 | 37.71 |
| 30.08 | 23.73 | 25 | 21.19 | |||
| 2400—2600 | 12 | 278 | 34.89 | 24.1 | 9.72 | 31.29 |
| 44.24 | 31.65 | 13.68 | 10.43 | |||
| 2600—2800 | 5 | 43 | 16.28 | 32.56 | 27.91 | 23.25 |
| 79.07 | 13.95 | 2.33 | 4.65 | |||
土壤总有机碳根据国家标准GB7857—1987[12]与LY/T1237—1999[13]采用重铬酸钾氧化-外加热法:在外加热条件下(油浴温度为175—180 ℃、沸腾5 min)用一定浓度的重铬酸钾-硫酸溶液氧化土壤有机质,剩余的重铬酸钾用硫酸亚铁滴定。通过所消耗的硫酸亚铁标准溶液的体积得有机碳含量。硫酸亚铁标准溶液的体积与有机碳含量的关系式为:
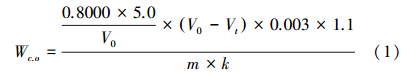
式中,Wc.o为有机碳含量(g/kg),0.8000为重铬酸钾标准溶液的浓度(mol/L),5.0为重铬酸钾标准溶液的体积(mL),V0为空白试验消耗硫酸亚铁标准溶液体积(mL),Vt为土样试验消耗硫酸亚铁标准溶液体积(mL),0.003为1/4碳原子的摩尔质量(g/mmol),1.1为氧化校正系数,m为风干土样质量(g),k为风干土样换算成烘干土样的水分换算系数。
在土壤有机碳含量的基础上,1 m深度土壤标准剖面不同层次土壤密度、厚度和非石砾所占的比例相乘得土壤有机碳密度SOCDk。
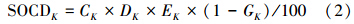
式中,SOCDk 为第k层土壤有机碳密度(kg/m),k为土壤层次,Ck为第k层土壤有机碳含量(g/kg),Dk为第k层土壤密度(g/cm3),Ek为第k层土层厚度(cm),Gk为第k层土层中直径大于2 mm石砾所占体积百分比(%)。
1.5 生物量计算方法刘广路[14]利用相对生长模型建立的天山云杉纯林生物量的回归模型W=0.075352(D2H)0.906442(W为生物量(kg),D为胸径(cm),H为树高(m))。本研究采用此模型,利用不同海拔云杉林标准木的胸径和树高的数据,来估计云杉林单木平均地上生物量。在单木平均地上生物量的基础上,得林份总生物量和单位面积生物量。即林分总生物量=整个研究区标准单木生物量×整个研究区云杉林总株数;单位面积生物量=(单木生物量×云杉林总株数)/样地总面积。
1.6 数据分析方法气象数据来自李艳忠[15]等从中科院新疆生地所地理信息系统实验室库所得的数据。综合考虑降水量与温度对植物的影响,分别表征年平均干湿度的Kira(K1)[16]干湿度指数和修正的生物干湿度指数(BK)[17, 18, 19]来表示地表干湿程度,从而判断不同海拔梯度上有机碳密度的变化与气候间的关系。
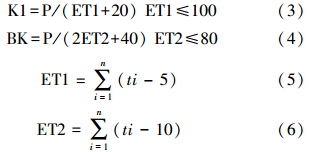
式中,P为年降水量,ET为有效温度,ti为i月份的平均温度。
本研究中所得到的数据采用Excel图标和SPSS17.0统计软件进行多重比较和相关性分析。
2 结果与分析 2.1 各海拔梯度土壤剖面的有机碳密度变化规律标准土壤剖面上,各海拔梯度的土壤有机碳密度随着剖面深度的增加呈减少的趋势(图 1)。海拔1800—2000 m的土壤有机碳密度从10.76—28.33 kg/m2(平均值为16.78 kg/m2)减少到3.12—7.36 kg/m2(平均值为5.24 kg/m2);海拔2000—2200 m的土壤有机碳密度从9.98—17.92 kg/m2(平均值为14.80 kg/m2)减少到2.10—3.13 kg/m2(平均值为2.60 kg/m2);海拔2200—2400 m的土壤有为碳密度从8.72—11.26 kg/m2(平均值11.06 kg/m2)减少到2.77—4.33 kg/m2(平均值为3.34 kg/m2);海拔2400—2600 m的土壤有机碳密度从10.68—19.04 kg/m2(平均值14.95 kg/m2)减少到1.94—3.18 kg/m2(平均值为2.50 kg/m2);海拔2600—2800 m的土壤有为碳密度从11.64—13.20 kg/m2(平均值12.19 kg/m2)减少到1.28—2.52 kg/m2(平均值为1.75 kg/m2)。海拔1800—2000 m土壤有机碳密度在标准剖面0—80 cm的四个土层中没有显著性差异;海拔2000—2200 m,2200—2400 m,2600—2800 m的标准土壤剖面40—100 cm的3个土层中没有显著性差异;海拔2400—2600 m的标准土壤剖面20—80 cm的3个土层中没有显著性差异。虽然不同海拔不同土壤深度有机碳密度的变化不存在显著差异性,但随着土壤深度的加深有机碳密度的整体性变小的趋势遵循了随着土壤深度的增加有机碳密度变小的规律。这与土壤不同深度的理化性质(容重、石砾含量、含水量)、微生物活性和凋落物的分解速率等有关。

|
| 图 1 同一海拔不同土层的有机碳密度变化规律 Fig.1 Soil organic carbon density variation of same altitudes at the different depth |
不同海拔梯度标准土壤剖面中同一土层的土壤有机碳密度呈先减少后增加再减少的趋势(图 2)。0—80 cm的4个土层中土壤有机碳密度在海拔1800—2000 m和2400—2600 m的梯度上出现双峰值,而在80—100 cm的土层中海拔1800—2000 m和2200—2400 m梯度上出现双峰值。不同海拔土壤0—80 cm的4个土层中土壤有机碳密度的平均值变化无显著性差异,在海拔1800—2000 m和2200—2400 m两个梯度上80—100 cm土层中的土壤有机碳密度平均值变化无显著性差别,而海拔2200—2400 m和其余的3个海拔梯度的土壤有机碳密度平均值无显著性差别。每个海拔梯度上同一土壤剖面土壤有机碳密度的变化规律最终导致从海拔下线到海拔上线的总有机碳密度呈现出先减少后增加再减少的趋势。

|
| 图 2 不同海拔同一土壤深度的有机碳密度变化规律 Fig.2 Soil organic carbon density variation of different altitudes at the same depth |
标准土壤剖面中土壤有机碳主要储存在于土壤表层中(图 3)。0—20 cm土层有机碳密度占全剖面所含的有机碳密度比例从30%(2200—2400 m)增加到43%(2600—2800 m);20—40 cm的比例从25%(1800—2000 m)增加到30%(2600—2800 m);最下3层所占的比例为25%到40%,可见土壤表层有机碳密度占全剖面的比例超过一半以上。这主要是以“表聚现象”所致[15],即植物的凋落物归还于土壤表层,在表层形成较厚的凋落叶层,经分解在土壤表层富集有机质从而形成相应大的有机碳密度。
2.3 总土壤有机碳密度沿海拔的变化规律随着海拔梯度的增加总土壤有机碳密度呈先减少后增加再减少的趋势(图 3)。总土壤有机碳密度平均在28.59—48.87 kg/m2之间,其中1800—2000 m海拔总有机碳密度最高,而海拔2600—2800 m的最低,整个变化趋势来看,各海拔有机碳密度变化没有显著性差异。
3 结论与讨论研究区的植被类型是以天山云杉纯林为主,可以避免受到其它植被的干扰,所以气候条件和云杉纯林的分布、长势是影响土壤有机碳密度变化的主要因素。
3.1 影响有机碳密度沿海拔分异的气候因素气候的变化通过影响天山云杉纯林的生长,凋落物的累积和分解而直接影响到土壤有机碳在不同海拔梯度上的分布情况。各海拔随着梯度的增加年均温度逐渐降低,到了2470 m左右时年均温度为零;随着海拔梯度的增加年均降水量出现先升高再下降的趋势(图 3)。
在不同海拔梯度上土壤有机碳密度的变化与气候单因素降水量和温度具有一定的相关性(表 2),根据Pearson相关系数的分析,所知土壤有机碳密度变化跟降水量呈负相关,而跟温度变化呈正相关。
| 总有机碳密度 Total organic carbon density/(kg/m2) | 降水量 Precipitation/ mm | 温度 Temperature/℃ | Kira干湿度指数 K1 | 修正的生物 干湿度指数 BK | |
| P<0.05 | |||||
| 总有机碳密度/(kg/m2) Total organic carbon density | 1 | ||||
| 降水量 Precipitation/mm | -0.688 | 1 | |||
| 温度 Temperature/℃ | 0.680 | -0.448 | 1 | ||
| Kira干湿度指数 K1 | -0.805 | 0.879* | -0.820 | 1 | |
| 修正的生物干湿度指数 BK | -0.428 | 0.615 | -0.202 | 0.499 | 1 |
由于仅仅利用气候的单因素不能对不同海拔土壤总有机碳密度变化的影响进行估测,因此,考虑综合指标地表干湿度指数对土壤有机碳密度的影响。为了能更好地评价气候要素对植被的影响,选取了最热月7月份的均温,此月森林带上下线相差为8.3 ℃,大于年均温,是对Kira干湿度指数和修正的生物干湿度指数的变化作用较大的温度。由于不同海拔月均温不同,所以K1和BK存在一定的差异(表 3)。
由于土壤总有机碳密度随海拔的变化情况跟地表干湿度指数有关,所以气候的变化导致不同海拔梯度上土壤总有机碳密度的差异性,即K1和BK与总土壤有机碳密度具有相关性(表 2)。
在此基础上进一步分析不同海拔梯度气候与土壤总有机碳密度间的相关性后发现,土壤有机碳密度跟地表干湿度指数具有负相关,表明温度和降水各指标与土壤有机碳密度变化的作用在所有云杉纯林区均具有同步性。
3.2 影响有机碳密度沿海拔分异的群落因素天山北坡的云杉林分布的上限和下限受温度和降水的综合影响[20, 21],乔木的胸径和树高能较好地指示森林带温度和降水特征[22, 23],也是影响土壤有机碳密度变化的重要指标,所以在研究区对不同海拔的树木进行每木调查,绘制了不同海拔树木胸径与树高关系的曲线图(图 3)。由图 3可知,从海拔下限1800 m到海拔上限2800 m,平均树高从14.99 m降低到8.25 m;平均胸径从海拔下限到海拔上限出现了先下降后增加再下降的趋势,跟不同海拔土壤总有机碳密度变化趋势大体上保持一致。
| 海拔 Elevation/m | 7月均温/℃ July average temperature | 有效温度 ET1 | 有效温度 ET2 | Kira干湿度指数 K1 | 修正的生物干湿度指数 BK |
| ET1为5 ℃以上的有效温度; ET2为10 ℃以上的有效温度 | |||||
| 1500 | 18.98 | 13.98 | 8.98 | 11.42 | 7.92 |
| 2000 | 15.06 | 10.06 | 5.06 | 18.06 | 12.05 |
| 2200 | 13.82 | 8.82 | 3.82 | 18.32 | 23.14 |
| 2600 | 11.34 | 6.34 | 1.34 | 18.79 | 11.97 |
| 2700 | 10.72 | 5.72 | 0.72 | 19.01 | 12.01 |

|
| 图 3 各层土壤有机碳密度占全剖面的百分比与不同海拔总土壤有机碳密度,年降水量,年均温度以及云杉林平均胸径树高的变化规律 Fig.3 Layers of soil organic carbon density as a percentage of the total cross-section and total organic carbon density,annual pricipitation,mean annual temperature,Picea schrenkiana DBH and tree height variation with different altitude |
生物量是一定时间,一定空间一种或数种生物有机体总干物质的重量[24],也是量度生态系统碳汇和碳素循环的关键数据[25]。对植物群落生物量进行研究,可以揭示生态系统中养分循环功能过程的变化规律[26]。因此,按照符合天山云杉林生物量的估测模型,对各海拔云杉林地上生物量进行估算(表 4),发现地上生物量呈随着海拔梯度的增加逐渐减少的趋势。海拔1800—2000 m的最高,而2600—2800 m的最低。最高和最低含量跟有机碳密度变化情况保持一致。
土壤有机碳密度变化与生物量存在紧密的联系。通过不同海拔云杉林单位面积上的生物量与有机碳密度的Pearson相关性分析(P<0.05),得生物量与有机碳密度呈正相关(相关系数为0.768)。即生物量的变化规律可直接反应土壤有机碳密度的变化规律。
| 海拔 Elevation/m | 平均胸径 Average DBH/cm | 平均树高 Average tree height/m | 生物量 Biomass/ (kg/m2) | ||
| ET1为5 ℃以上的有效温度; ET2为10 ℃以上的有效温度 | |||||
| 1800—2000 | 18.98 | 14.99 | 21.62 | ||
| 2000—2200 | 17.08 | 14.89 | 17.67 | ||
| 2200—2400 | 21.37 | 14.41 | 16.07 | ||
| 2400—2600 | 20.12 | 11.83 | 11.35 | ||
| 2600—2800 | 15.71 | 8.25 | 2.70 | ||
土壤总有机碳密度的变化是气候与树高、胸径共同作用的结果,因此导致不同海拔梯度有机碳密度大小存在差异性。海拔1800—2000 m平均树高最高、平均胸径占第3位,而BK相对整个海拔梯度而言较小,云杉林生物量最高,所以此梯度含有的有机碳密度最大。海拔2200—2400 mBK最大,云杉林平均胸径最高、平均树高占第3位,导致生物量的降低,因此,所具有的有机碳密度不是最多。海拔2400—2600 m平均胸径占第2位、平均树高占第4位,生物量跟平均树高的地位一样占第4位,对于整个研究区而言BK最小,导致有机碳密度在整个梯度上占第2位。海拔2600—2800 m虽然BK相对而言较小,但是平均胸径、平均树高和生物量都均最低,所以土壤有机碳密度最小。
3.4 结论本文通过不同海拔梯度云杉林的株树、平均胸径、树高调查与李艳忠[15]等提出的森林带发育于与气候土壤的关系时所提供的气象数据的基础上,对天山北坡水西沟区土壤有机碳密度与影响因素气候和云杉纯林胸径和树高的关系进行了探讨。结果表明:
(1)1m深度的标准土壤剖面上,各海拔梯度的土壤有机碳密度随着剖面深度的增加呈减少的趋势。平均有机碳密度从海拔1800—2000 m10—20 cm土壤深度的16.78 kg/m2减少到海拔2600—2800 m土壤深度80—100 cm的1.75 kg/m2。不同海拔各层土壤有机碳密度不显著,都出现先下降后增加再下降的趋势,导致总土壤有机碳密度具有同样的变化趋势。海拔1800—2000 m所具有的有机碳密度最大(48.87 kg/m2),其次是海拔2400—2600 m(41.66 kg/m2),2600—2800 m的最小(28.59 kg/m2)。
(2)土壤有机碳主要集中土壤表面(0—40 cm)。出现“表聚现象”,土壤表面所具有的有机碳密度所占的比例(60%—70%)超过总有机碳密度的一般以上。
(3)不同海拔云杉林的平均胸径、树高以及气候水热配比关系均出现一定的变化,土壤有机碳密度的变化跟生物量成正比而跟地表干湿度指数成反比关系。云杉林的生长趋势与气候的共同影响下导致不同海拔土壤有机碳密度差异性存在。
| [1] | Xiao H L. Climate change in relation to soil organic matter. Soil and Environmental Sciences, 1999, 8(4): 304-304. |
| [2] | Wang B, Sun G, Luo P, Wang M, Wu N. Labile and recalcitrant carbon and nitrogen pools of an alpine meadow soil from the eastern Qinghai-Tibetan Plateau subjected to experimental warming and grazing. Acta Ecologica Sinica, 2011, 31(6) 1506-1514. |
| [3] | Post W M, Emanuel W R, Zinke P J, Stangenberger A G. Soil carbon pools and world life zones. Nature, 1982, 298(5870): 156-159. |
| [4] | Yang Y S, Xie J S, Sheng H, Chen G S, Li X, Yang Z J. The impact of land use/cover change on storage and quality of soil organic carbon in midsubtropical mountainous area of southern China. Journal of Geographical Sciences, 2009, 19(1): 49-57. |
| [5] | Zang Q J, Yu X B, Qian J X, Xiong T. Distribution characteristics of plant communities and soil organic matter and main nutrients in the Poyang Lake Nanji Wetland. Acta Ecologica Sinica, 2012, 32(12): 3656-3669. |
| [6] | Bouwman A F. Global distribution of the major soils and land cover types // Soils and the Greenhouse Effect. Chichester: John Wiley & Sons, 1990: 33-59. |
| [7] | Davidson E A, Trumbore S E, Amundson R. Biogeochemistry: soil warming and organic carbon content. Nature, 2000, 408(6814): 789-790. |
| [8] | Nichols J D. Relation of organic carbon to soil properties and climate in the southern great plains. Soil Science Society of America Journal, 1984, 48(6): 1382-1384. |
| [9] | Wang H Y, Lei X D, Zhang H R, Yang P. Soil organic carbon in semi-natural mixed larch-spruce-fir stands of northeastern China. Journal of Bei jing Forestry University, 2009, 31(3): 11-16. |
| [10] | Wang W F, Liu B F, Liu G J, Liang S Y, Liu Y. Comparison of soil organic carbon contents of typical stand types in Xiaoxing'anling. Forestry Science & Technology, 2011, 36(1): 17-19. |
| [11] | Zhang Y T, Chang S L, Lu J J, Li X, Wang Z, Shi Q D, Zhang X P. Large scale permanent plot developed and its three-dimension realized in Tianshan forest. Scientia Silvae Sinicae, 2011, 47(10): 179-183. |
| [12] | Liu G S. Jiang N H, Zhang L D, Liu Z L. Soil Physical and Chemical Analysis & Description of Soil Profiles. Beijing: Standards Press of China, 1996: 166-167. |
| [13] | LY/T1237-1999. Determination of organic matter in forest soil and calculation of carbon-nitrogen ratio. Beijing: State Forestry Administration, 1999. |
| [14] | Liu G L. The Study on the Growth Rule of Picea schrenkiana var. Tianschanica and the Productivity of Communities in Tianshan[D]. Baoding: Agricultural University of Hebei, 2006. |
| [15] | Li Y Z, Luo G P, Xu W Q, Yin C Y, Han Q F, Feng Y X. Forest development and their relationships with climatic and soil in the mid-mountain area of Sangong river watershed, northern slope of Tianshan mountains. Journal of Mountain Science, 2011, 29(1): 33-42. |
| [16] | Yim Y, Kira T. Distribution of forest vegetation and climate in the Korean Peninsula. I. Distribution of some indices of thermal climate. Japanese Journal of Ecology, 1975, 25(2): 77-88. |
| [17] | Yim Y, Kira T. Distribution of forest vegetation and climate in the Korean Peninsula. II. Distribution of climatic humidity/aridity. Japanese Journal of Ecology, 1976, 26(3): 157-164. |
| [18] | Xu W D. The relation between distribution of edificator and companion in zonal vegetation and water-temperature condition in Northeast China. Journal of Integrative Plant Biology, 1983, (3): 264-274. |
| [19] | Ni J. Development of Kira's indices and its application to vegetation-climate interaction study of China. Chinese Journal of Applied Ecology, 1997, 8(2): 161-170. |
| [20] | Zhu H F, Wang L L, Shao X M, Fang X Q. Tree ring-width response of Picea schrenkiana to climate change. Acta Geogmraphica Sinica, 2004, 59(6): 863-870. |
| [21] | Sang W G, Wang Y X, Su H X, Lu Z H. Response of tree-ring width to rainfall gradient along the Tianshan Mountains of northwestern China. Chinese Science Bulletin, 2007, 52(21): 2954-2962. |
| [22] | Chen S Y, Guo J Y, Han T, Shi Y Y. Climate warming of autumn air temperature in arid and semiarid Regions in Northwest China over the recent 46 years. Journal of Desert Research, 2009, 29(3): 544-550. |
| [23] | Luo R Y. Forest Soil Science (Problems and Methods). Beijing: Science Press, 1983. |
| [24] | Wen J C. Tree size distribution functions of four boreal forest types for biomass mapping. Forest Science, 2004, 50(4): 436-439. |
| [25] | Houghton J T, Ding Y, Griggs D J, Noguer M, van der Linden P J, Dai X, Maskell K, Johnson C A. Climate Change 2001: The Scientific Basis. Cambridge: Cambridge University Press, 2001: 94-94. |
| [26] | Krebs C J. Ecology: The Experimental Analysis of Distribution and Abundance. New York, London: Harper-Collins College Publishers, 2001: 513-536. |
| [2] | 王蓓, 孙庚, 罗鹏, 王萌, 吴宁. 模拟升温和放牧对高寒草甸土壤有机碳氮组分和微生物生物量的影响. 生态学报, 2011, 31(6) 1506-1514. |
| [5] | 张全军, 于秀波, 钱建鑫, 熊挺. 鄱阳湖南矶湿地优势植物群落及土壤有机质和营养元素分布特征. 生态学报, 2012, 32(12): 3656-3669. |
| [9] | 王海燕, 雷相东, 张会儒, 杨平. 近天然落叶松云冷杉林土壤有机碳研究. 北京林业大学学报, 2009, 31(3): 11-16. |
| [10] | 王文帆, 刘滨凡, 刘广菊, 梁素钰, 刘禹. 小兴安岭典型林分类型土壤有机碳含量状况比较分析. 林业科技, 2011, 36(1): 17-19. |
| [11] | 张毓涛, 常顺利, 芦建江, 李翔, 王智, 师庆东, 张新平. 天山云杉森林8hm2样地的建立及三维可视化管理. 林业科学, 2011, 47(10): 179-183. |
| [12] | 刘广崧, 蒋能慧, 张连第, 刘兆礼. 土壤理化分析与剖面描述. 北京: 中国标准出版社, 1996: 166-167. |
| [13] | LY/T1237—1999. 森林土壤有机质的测定及碳氮比的计算. 北京: 国家林业局, 1999. |
| [14] | 刘广路. 天山云杉生长规律与天山植物群落生产力研究 [D]. 保定: 河北农业大学: 硕士毕业论文, 2006. |
| [15] | 李艳忠, 罗格平, 许文强, 尹昌应, 韩其飞, 冯异星. 天山北坡三工河流域中山带森林发育与气候土壤的关系. 山地学报, 2011, 29(1): 33-42. |
| [18] | 徐文铎. 东北地带性植被建群种及常见种的分布与水热条件关系的初步研究. 植物学报, 1983, (3): 264-274. |
| [19] | 倪健. Kira指标的拓展及其在中国植被与气候关系研究中的应用. 应用生态学报, 1997, 8(2): 161-170. |
| [20] | 朱海峰, 王丽丽, 邵雪梅, 方修琦. 雪岭云杉树轮宽度对气候变化的响应. 地理学报, 2004, 59(6): 863-870. |
| [22] | 陈少勇, 郭江勇, 韩通, 石圆圆. 中国西北干旱半干旱区近46a秋季气候变暖分析. 中国沙漠, 2009, 29(3): 544-550. |
| [23] | 罗汝英. 森林土壤学(问题和方法). 北京: 科学出版社, 1983. |