 2014, Vol. 34
2014, Vol. 34文章信息
- 冯永玖, 陈新军, 杨铭霞, 霍丹, 朱国平
- FENG Yongjiu, CHEN Xinjun, YANG Mingxia, HUO Dan, ZHU Guoping
- 基于ESDA的西北太平洋柔鱼资源空间热点区域及其变动研究
- An exploratory spatial data analysis-based investigation of the hot spots and variability of Ommastrephes bartramii fishery resources in the northwestern Pacific Ocean
- 生态学报, 2014, 34(7): 1841-1850
- Acta Ecologica Sinica, 2014, 34(7): 1841-1850
- http://dx.doi.org/10.5846/stxb201305191105
-
文章历史
- 收稿日期:2013-5-19
- 修订日期:2013-11-19






2. 大洋渔业资源可持续开发省部共建教育部重点实验室, 上海 201306;
3. 国家远洋渔业工程技术研究中心, 上海 201306
2. The Key Laboratory of Sustainable Exploitation of Oceanic Fisheries Resources (Shanghai Ocean University), Ministry of Education, Shanghai 201306, China;
3. National Distant-water Fisheries Engineering Research Center (Shanghai Ocean University), Shanghai 201306, China
柔鱼Ommastrephes bartramii广泛分布于西北太平洋海域,是一种经济大洋性头足类,已成为日本、韩国、中国(台湾省)等国和地区的重要捕捞对象[1, 2]。开展柔鱼资源时空格局分析与潜在规律的挖掘,能够为柔鱼资源的可持续开发与资源评估管理提供一定的参考依据。
目前有关学者对柔鱼基础生物学[1, 3]、资源量分布变动及其机制[4, 5, 6]、资源量评估[7]等,以及时空格局及其与海洋环境的关系[8, 9]、渔情预报[10, 11]等领域进行较为广泛的研究。在渔业资源空间分布或格局的研究中,利用传统方法分析柔鱼资源空间变动的文献较多[5, 6, 8],这些研究为初步掌握柔鱼资源分布规律奠定了基础。
传统GIS可直观表达渔业资源的空间分布,并通过数据或语言文字进行位置分布的描述[12],但却无法挖掘和揭示渔业资源的全局空间分布模式和内在的空间关联关系。迄今,渔业资源相关空间问题的深层次研究仍属鲜见,如有关渔业资源的空间聚类、渔业生态学过程和格局的空间异质性等问题。针对渔业资源的空间问题,国内学者利用遥感、GIS、空间分析和地学关联规则等方法,对东海区鱼类资源的时空格局和变化进行了一系列研究[13, 14],展示了GIS空间分析与地统计方法在渔业资源研究中的广阔前景。近年来,杨晓明等利用空间变异函数分析了印度洋黄鳍金枪鱼(Thunnusal bacares)围网渔获量的空间异质性[15],并利用空间自相关和变异函数分析了中西太平洋鲣鱼(Katsuwonus pelamis)围网渔业资源的空间热点和空间异质性[16];牛明香则利用空间自相关和标准差椭圆分别研究了东海鳀鱼(Engraulis japonicus)的空间格局与定向分布特征[17],展示了空间统计分析在渔业资源研究中的最新成果。
探索性空间数据分析方法(ESDA)[18, 19]是一种典型的GIS数据分析方法,能够有效探测空间数据中潜在的关联关系并发现数据分布趋势。为此,本研究拟综合利用全局和局部空间自相关方法[20, 21],有效识别柔鱼资源的空间自相关结构、热点和冷点分布格局,利用克里金插值[23]获取柔鱼资源热冷点分布的面状格局,并利用变化检测和景观分析等方法分析柔鱼资源热冷点格局的变动,多角度揭示西北太平洋柔鱼资源的整体空间模式、热冷点区域及其变动规律,为柔鱼资源空间分布规律的掌握提供科学方法。
1 材料与方法 1.1 研究区域与数据以西北太平洋柔鱼资源为研究对象,研究范围为150°E—160°E、38°N—45°N(图 1)。数据获取时间为2007年和2010年,其中2007年为5—11月,2010年为5—12月。本文采用的是原始数据,即每个数据点的位置就是捕捞渔船所记录的空间位置,2007年获取2212个数据点,2010年获取7918个数据点。将数据点的值换算为单位捕捞努力量渔获量(CPUE),用以代表每个年度的柔鱼资源丰度。本文采用的CPUE换算公式为C/E,式中C表示1艘渔船1d的产量(t),E表示其对应的作业次数[22]。此外,为了准确地获取柔鱼资源空间热点范围,利用GIS中的不等边三角网TIN(Triangulated Irregular Network)方法[23]建立渔获量数据范围(图 1)。通过建立TIN能够剔除无渔获量的海域并产生有效数据区域,使热冷点的分析不受或少受无数据区域的影响,从而提高空间分析的可靠性。

|
| 图 1 研究区域与数据 Fig.1 The study area and the dataset used |
为研究柔鱼资源在全局空间上可能存在的聚集、离散或随机模式,采用ESDA方法中的全局空间自相关统计量Moran′s I进行度量,其计算公式如下:
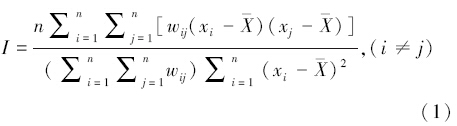
式中,n是参与分析的要素数量(即样本数量),xi是要素i的属性值,xj是要素j的属性值,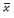 是全部要素的平均值,wij是空间权重矩阵,表示要素i 和j的邻近关系,它可以根据邻接标准或者距离标准来度量。wij=1表示第i和j个要素相邻,wij=0表示第i和j个要素不相邻。
是全部要素的平均值,wij是空间权重矩阵,表示要素i 和j的邻近关系,它可以根据邻接标准或者距离标准来度量。wij=1表示第i和j个要素相邻,wij=0表示第i和j个要素不相邻。
全局空间自相关Moran′s I值域范围为[-1, 1],该值大于0表示正相关,小于0表示负相关;Moran′s I绝对值越大表示空间分布的自相关程度越高,表明空间分布呈现聚集现象;Moran′s I绝对值越小代表空间分布的自相关程度越低,说明空间分布呈现分散格局;当Moran′s I值等于0 时,表示空间分布呈现随机分布[24]。在实际计算中Moran′s I返回另外两个值:Z得分和P值,其中Z得分是I标准差的倍数,当Z较大时表示要素呈聚集分布状态。P值表示样本空间模式是某一随机分布的概率,当P值很小时表示探测所得的空间模式不太可能是随机分布,当P值较大则表示空间模式为随机分布的概率较大[24]。
1.3 局部空间自相关分析方法全局空间自相关统计量Moran′s I反映的是柔鱼资源空间整体自相关状态,侧重于柔鱼资源的全局空间模式[18, 19, 20, 21],但是却无法揭示资源局部的分布状态。此外,全局空间自相关还存在一些局限性,例如整体聚集的情况下可能存在局部的随机分布,同样整体随机分布的情况下也可能存在局部聚集分布。因此,通过局部空间自相关统计量分析不仅可以识别局部分布特征,更能探测柔鱼资源的热点和冷点区域[18, 19, 20, 21]。
Getis-Ord G*i是空间热点分析中常用的方法,该统计量产生两个值:即每个要素的Z得分和显著性P值。Getis-Ord G*i统计量的计算公式如下:
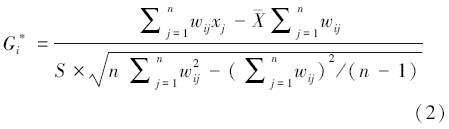
式中,xj是要素j的属性值,wij表示要素i和j之间的空间权重,其意义与式(1)相同,n是要素数量, 为均值,S为标准差。G*i统计结果返回Z得分和P值,其意义与全局空间自相关类似。当Z得分大于2倍标准差,表示空间热点区域;当Z得分介于1倍与2倍标准差之间、-2倍与-1倍标准差之间,均表示可能出现一定的热冷点分布,但不能否定随机分布的可能;当Z得分介于-1倍与1倍标准差之间,则表示空间模式有极大可能是随机分布;当Z得分小于-2倍标准差,表示空间冷点区域[24]。此外,热点区域表示要素高值被高值包围,而冷点表示要素低值被低值包围,可用于揭示柔鱼资源渔获量高值或者低值在空间上发生聚类的位置[16]。
1.4 柔鱼资源空间热冷点格局的评价方法与传统研究不同,本文侧重从地理信息科学的视角去解释渔业资源空间格局及其变动。GIS中图层叠置方法(Overlay)是空间分析的经典方法,在遥感与GIS、资源与环境等领域中应用广泛[23]。本研究中,利用叠置方法分析柔鱼资源热冷点格局随时间的变化。
同时,从生态学和空间科学的角度来看,柔鱼资源热冷点分布图的基本组成单元是斑块(Patch),因此是一种典型的景观镶嵌图,故而非常适合于利用景观指数(Landscape Metrics)来评价和分析其结构[25, 26]。景观指数是能够高度浓缩景观格局信息,反映其结构组成和空间配置的简单定量指标[27, 28, 29]。景观指数包括景观层次(Landscape-level)、类型层次(Class-level)和斑块层次(Patch-level)3种指标[26],可从不同层次评价渔业资源的空间热冷点结构。
2 结果 2.1 常规统计与全局空间自相关分析为了解柔鱼资源整体情况,对其获取的样本数据进行常规统计量及全局空间自相关统计量测算(表 1)。结果显示,柔鱼样本在2007和2010年的偏态Sk均大于0,频数分布均为正偏;2007年的峰态Ku小于3,呈现平峰分布,表明高产值海域较多,2010年的峰态Ku大于3,呈现尖峰分布,表明2010年低产值海域较多;变异值Cv显示,柔鱼样本量在2007和2010年的Cv值均大于0,表明不同空间位置柔鱼资源差异较大,而2010年差异程度大于2007年;此外,2007和2010年的S2/m值均大于1,表明柔鱼资源呈现较强的聚集分布特征。
|
年份
Year | 最大值
Max./ (T/d) | 均值m/
(T/d) | 标准差S
Std deviation | 偏态
Skewness | 峰态
Kurtosis | Cv=s/m | S2/m | Global
Moran′s I* | Z得分
Z-score* | P |
| *:Global Moran′s I、Z-score和P均为全局空间自相关统计量 | ||||||||||
| 2007 | 18 | 5.2114 | 2.7278 | 0.9835 | 1.3936 | 0.5234 | 1.4278 | 0.1619 | 177.8163 | 0.0000 |
| 2010 | 15 | 2.0018 | 1.4959 | 1.9772 | 6.8805 | 0.7473 | 1.1179 | 0.2453 | 409.6437 | 0.0000 |
全局自相关Moran′s I统计量表明,西北太平洋柔鱼资源在2007和2010年均为正相关,且呈现一定的聚集特征,印证了S2/m的计算结果。同时,两个年份的Z得分均非常高,且P值均为0,表明柔鱼资源呈现出显著的聚集分布模式。
2.2 局部空间自相关与空间热冷点分布格局根据上述理论和方法,在ArcGIS桌面软件中对2007和2010年进行计算和渲染,所得的热冷点区域分布如图 2。其中,GiZscore表示Getis-Ord G*i值的Z得分,点状GiZscore是利用ArcGIS渲染之后的可视化图形,而面状GiZscore为经过ArcGIS克里金插值方法得到的可视化结果[24, 30]。

|
| 图 2 西北太平洋柔鱼资源空间热点和冷点分布特征比较 (2007和2010年) Fig.2 A comparison of distribution of hot/cold spots between 2007 and 2010 for the fishery resource of O. bartramii in the northwestern Pacific Ocean |
图 2表明,2007年柔鱼资源的空间热点有8个区域,但面积足够大且视觉上可见的只有3个,其中心位置分别为156°E/44°N(A区)、152°E/ 42°N(B区)和158°E /40°N(C区),其中A区柔鱼资源高值最为密集,C区密集度次之但范围最大,而B区密集度最低且范围最小;同时2007年具有3个冷点区域,但只有1个冷点区域范围较大且视觉上可见,其中心位置为154°E/43°N(D区)。图 2表明,2010年柔鱼资源的空间热点有1个区域,其中心位置为157°E/43°N(E区),与2007年空间热点A区在空间距离上较为接近,但其范围和密集度大于2007年A区;此外,2010年具有5个冷点区域,其中4个区域视觉上可见,中心位置分别为151°E /42°N(F区)、153°E/42°N(G区)、155°E/40°N(H区)和157.5°E/39.5°N(I区),其中H区范围最大且资源低值的密集度也较大,G区范围最小且低值密集度也最小。
2.3 空间热冷点的构成为了研究柔鱼资源空间热冷点的构成及百分比,基于面状的空间热点区域,对GiZscore值包含的5种类型和“No sample”数据区进行统计如图 3。图 3表明,2007年无数据区域占研究区范围46.1%,2010年无数据区则仅占26.8%。2007年热点区域占10.4%,2010年仅占3.8%,表明柔鱼资源高值被高值包围的范围减少;2007年冷点区域占11.3%,2010年占13.1%,比之2007年增加了近2%,表明柔鱼资源低值被低值包围的范围有所增大;不管是2007年还是2010年,其余非热点和冷点(即聚集性不显著或呈随机分布)所占的面积较大,尤其是2010年GiZscore值在[-2.0,-1.0]范围内的面积达到了31.0%,表明研究区内大部分渔获量并不高。对比2010年原始点位数据,CPUE≤2.0的数据点为4048个,占总数据量的70.5%。

|
| 图 3 2007和2010年西北太平洋柔鱼资源热冷点构成 Fig.3 Percentage of hot/cold spots for the fishery resource of O. bartramii in the northwestern Pacific Ocean in 2007 and 2010,respectively |
基于GIS的空间叠置分析,本文检测了柔鱼资源热冷点在空间位置上的变动(图 4)。

|
| 图 4 2007和2010年西北太平洋柔鱼资源热冷点变动的比较 Fig.4 A comparison of hot/cold spots change between 2007 and 2010 for the fishery resource of O. bartramii in the northwestern Pacific Ocean |
图 4表明,对比2007和2010年仅有1个区域为空间热点不变,其中心位置为156.5°E/44°N;从2007年空间热点变成2010年空间冷点的区域有2个,中心点分别位于151.5°E/42°N和157.5°E/39.5°N;对比2007和2010年呈现空间冷点不变的区域有1个,其中心位置为156.5°E/44°N。此外,聚集性不显著或呈随机分布的非热点和冷点区域之间的变动,在研究区域占据了主导地位,但是热点和冷点之间的变动是本区域的柔鱼资源的关键性信息。
3 分析与讨论 3.1 关于渔区划分和TIN预处理在传统渔业资源研究中,无论是有关渔情预报[1, 2, 11]、HSI建模[10]还是空间分析[13, 14, 15, 16, 17],均将渔获量分成不同的渔区进行统计,如0.5°×0.5°为一个渔区。这样的渔区划分方法非常经典,也具有其理论依据与实践意义,在渔业资源研究中被广泛接受。从空间角度来看,渔区的划分是否能够准确地代表采样点的空间特性,是需要结合渔业捕捞作业和空间采样理论来综合考虑的。在这方面,形成了一个既有实践价值又具有科学意义的主题,即渔业资源CPUE的标准化[22]。渔区划分和CPUE标准化之后的空间数据,在空间中形成了规则的分布模式。但是,渔业资源的空间分析主要是从空间视角对其分布进行考察;而从GIS的角度来看,空间分析偏好不规则的数据分布形式,并从中挖掘潜在的规律[24]。因此,本文利用的数据为原始捕捞记录数据,即每个数据点位置就是捕捞渔船所记录的空间位置,其属性值即为CPUE。基于这种不规则的原始空间数据,能够获取更为准确的空间热点和冷点区域。
此外,研究范围(150°E—160°E、38°N—45°N)内很多区域是没有渔业捕捞数据的,即渔船并没有到达这些区域生产。但是,这并不表明这些区域没有渔业资源。如果将这些没有数据的区域纳入空间热点的研究,很可能会增加空间分析的不确定性和误差。因此,将无数据区域划分为“No samples”类型,不参与空间热点的分析,这将极大降低GIS空间分析的不确定性。然而,无数据区域的确定是一个困难的问题。本文采用了GIS中的TIN三角网方法[23],将数据采样区域和无采样区域分开。规则的渔区分布数据不能体现TIN的不规则特性,很难构造有效的TIN三角网,再次印证了利用原始数据进行分析的必要性。由于GIS中的TIN三角网必须是凸包[23],而图 1中所示的TIN包含了凹多边形,因此它实际上并不是GIS中的标准TIN三角网。这是因为,如果采用标准TIN,很多无渔获量区域将被包括在有数据的区域内。因此,本文对标准TIN进行修改,将被包含在标准TIN内、但无渔获量的区域剔除,形成了图 1所示的三角网。由于剔除了数据无关区域,故而在点状和面状热冷点区域的探测中,由于原始数据造成的误差则会减小;因此,TIN三角网的构建能够很好地辅助柔鱼资源的空间热点分析,从而减小分析的不确定性。
3.2 基于景观指数的空间热冷点格局评价本文同时采用了景观和类型两种层次的指数,分别对柔鱼资源的空间热冷点进行评价。景观层次指数评价的是柔鱼资源每一年的整体空间结构,类型层次指数评价的是柔鱼资源的热点、冷点或随机分布等每一类型的空间结构。在计算景观指数时去除了“No samples”,即将无样本的海域当作背景进行处理,结果分别见表 2和表 3。
| 年份Year | NP | LPI/% | LSI | PAFRAC | CONTAG | DIVISION | SHDI | SIDI |
| NP:区域中与周围环境在外貌和性质上有别的均质性生态景观单元的数量;LPI:景观中最大斑块占整体景观的比例;LSI:反映斑块形状的破碎化程度和复杂程度;PAFRAC:景观中斑块的自相似及复杂程度的指标;CONTAG:景观中不同斑块类型的非随机性或聚集程度;DIVISION:景观分裂指数;SHDI:香浓多样性指数,表明斑块类型的均衡化趋势;SIDI:辛普森多样性指数,意义与SHDI类似[26, 27, 28, 29] | ||||||||
| 2007 | 31 | 16.2849 | 3.4097 | 1.2031 | 47.7832 | 0.9641 | 1.5378 | 0.7744 |
| 2010 | 19 | 30.0079 | 3.2199 | 1.1542 | 52.4525 | 0.8763 | 1.4241 | 0.7236 |
|
年份
Year | 空间热冷点
Hot/Cold spots | NP | LPI /% | LSI | IJI | COHESION | DIVISION | AI |
| LPI:景观中最大斑块占某类型景观的比例;IJI:散布与并列指数;COHESION:斑块的整体性和凝聚度;AI:斑块异质性增加与景观多样性退化过程中的景观聚集指数;其余NP、LSI和COHESION的意义与表 2相同,但在本表中属于类型层次[26, 27, 28, 29] | ||||||||
| 2007 | >2.0 | 8 | 5.7816 | 2.5146 | 21.4769 | 98.6967 | 0.9955 | 98.5103 |
| 1.0—2.0 | 9 | 9.3199 | 5.0506 | 50.0000 | 99.1187 | 0.9873 | 96.8180 | |
| -1.0—1.0 | 6 | 6.6647 | 4.7110 | 67.2694 | 98.8160 | 0.9947 | 96.5395 | |
| -2.0—-1.0 | 5 | 3.5828 | 4.6412 | 49.1255 | 98.1711 | 0.9987 | 94.3356 | |
| <-2.0 | 3 | 16.2849 | 1.8364 | 17.1953 | 99.3280 | 0.9880 | 99.2108 | |
| 2010 | >2.0 | 1 | 3.7564 | 1.1345 | 0.0000 | 98.6933 | 0.9986 | 99.7695 |
| 1.0—2.0 | 2 | 9.2049 | 2.4691 | 45.4337 | 99.1891 | 0.9915 | 98.4677 | |
| -1.0—1.0 | 5 | 12.8751 | 3.8091 | 49.2476 | 99.2928 | 0.9831 | 97.6315 | |
| -2.0—-1.0 | 6 | 30.0079 | 3.8246 | 48.4071 | 99.7475 | 0.9099 | 98.3298 | |
| <-2.0 | 5 | 7.0042 | 2.5785 | 0.0000 | 98.6430 | 0.9933 | 98.5594 | |
表 2中NP指数显示,2007和2010年西北太平洋柔鱼资源分别包含31和19个斑块,表明2007年热冷点的组成结构较2010年复杂;LPI指数表示景观中最大斑块占整体景观的比例,对应2007和2010年的GiZscore值,最大斑块所属类型分别为“1.0—2.0”和“-2.0—-1.0”,与图 3所示结果一致;LSI指数大于1,表明热冷点格局的几何形状稍显复杂,但是这种复杂程度并不高;PAFRAC指数的意义为热冷点格局的分维数,表 2中具体数值说明其复杂性和自相似并不高,但2007年比2010年复杂程度稍高;CONTAG指数较高,表明热冷点格局的聚集性非常强,即存在优势热点或冷点斑块;DIVISION指数介于0到1之间,0表示热冷点只由1个斑块组成,而1表示热冷点由很多复杂的斑块组成,表 2显示了在斑块组成结构上2007年的复杂程度高于2010年;同时,SHDI和SIDI指数均表明了柔鱼资源热冷点格局的多样性呈均衡化趋势,也说明了柔鱼资源的异质性较强。该空间异质性反映出,在空间结构上有多种类型的柔鱼分布形式(热点、冷点、统计不显著和随机分布等),交错地存在于西北太平洋海域,且在各个方向上其分布性质也不相同。
表 3中NP指数显示了热点、冷点等各类型GiZscore值对应的斑块数量,对应于“>2.0”为空间热点,对应于“<-2.0”为空间冷点,同时各类型斑块的总数与表 2一致;LPI指数揭示各类型中最大斑块占总景观(热点和冷点分布)面积的比例,如2010年中30.0079%表示GiZscore值介于-2.0和-1.0之间的面积占总景观的比例;LSI表明各类型结构的复杂性,可以看出2007年空间热点的复杂程度(2.5146)大于2010年(1.1345),2007年有8个热点而2010 年只有1个热点,很显然2007年更加复杂;与此类似,2007年的空间冷点复杂程度低于2010年;IJI指数表明,其值越高则热冷点的空间异质性越高,各类型之间的连接性下降,这说明2007年柔鱼资源的异质性高于2010年,而2010年热点(>2.0)和冷点(<-2.0)呈规则形状分布的趋势(IJI为0)较明显;COHESION指数表明了柔鱼资源空间热冷点各类型的关联程度,从具体数据可知COHESION较大且接近于100,表明各类型的整体性和凝聚度均很高;表 3中各类型的AI指数均较大且接近于100,表明各类型的聚集度较高。
可见,将柔鱼资源的热冷点空间格局看成一种景观分布,利用景观和类型层次两种指数对其进行刻画和分析,能够深入挖掘其内在结构与特征。总结起来,西北太平洋柔鱼资源的空间特征包括:1)中心位置位于156.5°E/44°N的区域可能是多年的热点和高产量区域;2)冷点或统计不显著的区域在该海域是优势斑块;3)热冷点格局的聚集性非常强;4)资源分布的异质性较高。
3.3 空间热冷点与海表温的关系柔鱼资源热冷点格局的变动,很大程度上反映了中心渔场的变动,而引起这种变动的因素是多方面的,如渔业捕捞本身、海洋环境要素(亲潮、表温及饵料对象等)以及全球气候变化等。本文以国际海洋水色协调组(IOCCG,http://oceancolor.gsfc.nasa.gov)提供的年平均海表温度(Annual Mean SST)为例,分析柔鱼资源空间热冷点分布与海洋环境的关系(图 5)。

|
| 图 5 西北太平洋年平均海表温度与柔鱼空间热冷点的关系 (2007和2010年) Fig.5 The relationship between annual mean sea surface temperature and hot/cold in the northwestern Pacific Ocean in both 2007 and 2010 黑线:热点区域,白线:冷点区域 |
根据图 5中西北太平洋柔鱼资源热冷点与年平均海表温的叠加显示,同时结合日本气象厅发布的数据和报告(http://www.data.kishou.go.jp),可知2007年西北太平洋海域平均海表温度总体上略低于正常水温,但从6月中旬开始在整个盛渔期平均海表温有不同程度的升高,月平均增长3℃,持续至10月中旬开始有所回落。此阶段黑潮和亲潮交汇相互作用最为强烈,2007年亲潮势力为1997年以来最弱,但是受影响的水域面积较小,相对来说黑潮势力占主导优势[1]。黑潮自31°N向北蜿蜒,2月产生了向东的离岸流,并形成几个小分支蜿蜒至西北太平洋[9]。西北太平洋柔鱼渔区的形成受温度和海流的影响,近一半作业渔区产量相对较高,但并未充分保证空间热点的形成(图 2),最终形成空间热冷点分布面积大致相当的格局。
图 5表明西北太平洋海域2010年平均海表温度稍高于2007年,亦为非大弯曲年份。该年亲潮势力强盛,3月到达37°N附近,至5月亲潮最南端靠近39°N,慢慢形成了暖涡,在暖涡附近产生的上升流带来了丰富的营养盐等条件,形成了一个中心渔场,一直持续至8月[1]。同时2010年黑潮势力较弱,越往东流力量越弱,没有形成主流。因此整个西北太平洋柔鱼渔场受亲潮势力影响,盛渔期渔场集中分布在40°—45°N、154°—158°E,随亲潮的流动渔场有所迁移[1]。根据本文对空间热冷点分布的探测,显示在38°—45°N、150°—160°E海域空间冷点较多,较为分散,大多分布在43°N以南;而热点区域非常集中于42°—45°N、155°—158°E海域(图 2)。这与受亲潮势力影响的渔区所在海域基本一致,反应了柔鱼鱼群为集中分布,并指示了中心渔场所在的区域。
4 结论本文利用ESDA中全局和局部空间自相关方法,对西北太平洋柔鱼资源的分布格局进行了探索性分析。基于局部空间自相关方法,对2007和2010年西北太平洋柔鱼资源的空间热冷点结构进行了可视化制图,结果显示该区域2007年存在3个显著的热点和1个冷点,2010年存在1个热点和4个冷点;聚集性不显著或呈随机分布的非热冷点区域之间的变动,在研究区域占据了主导地位。结合海洋环境因素,并利用百分比堆积图、空间变化检测和景观指数等方法进行了深入分析与挖掘,揭示了西北太平洋柔鱼资源中的内在特征。
此外,所有空间分布及其规律均是在建立在某种空间尺度的基础之上的。本文采用不是经过渔区划分的柔鱼资源数据,而是捕捞作业的原始点位数据,因此在后续研究中将对建立在渔区划分之上的热冷点格局进行补充分析,并与本研究的结果进行比较,同时考虑渔区空间尺度对热冷点分析的影响。
| [1] | Chen X J, Tian S Q, Chen Y, Cao J, Ma J, Li S L, Liu B L. Fishery Biology for Ommastrephes Bartramii in the Northwestern Pacific Ocean. Beijing: Science Press of China, 2011. |
| [2] | Chen X, Liu B, Chen Y. A review of the development of Chinese distant-water squid jigging fisheries. Fisheries Research, 2008, 89(3): 211-221. |
| [3] | Lin Q, Jin X S, Zhang B. Trophic interactions, ecosystem structure and function in the southern Yellow Sea. Chinese Journal of Oceanology and Limnology, 2013, 31(1): 46-58. |
| [4] | Liu W, Lin Z J, Jiang Y E, Huang Z R. Spatial distribution of demersal fishery resources in the continental shelf of the northern South China Sea. Journal of Tropical Oceanography, 2011, 30(5): 95-103. |
| [5] | Li S F, Yan L P, Li H Y, Li J S, Cheng J H. Spatial distribution of cephalopod assemblages in the Region of the East China Sea. Journal of Fishery Sciences of China, 2006, 13(6): 936-944. |
| [6] | Chen X J, Xu L X, Tian S Q. Spatial and temporal analysis of Ommastrephe bartrami resources and its fishing ground in North Pacific Ocean. Journal of Fisheries of China, 2003, 27(4): 334-342. |
| [7] | Wang H Z, Li C H, Jia X P. The fishery production from the north part of the South China Sea using the primary production method. Marine Fisheries Research, 2005, 26(3): 9-15. |
| [8] | Yang S L, Zhang Y, Zhang H, Wang D, Ma J J, Zhang J, Cui X S. The relationship between the temporal-spatial distribution of fishing ground of yellowfin tuna (Thunnus albacares) and themocline characteristics in the tropic Indian Ocean. Acta Ecologica Sinica, 2012, 32(3): 671-679. |
| [9] | Wang W Y, Zhou C H, Shao Q Q, Mulla D J. Remote sensing of sea surface temperature and chlorophyll-a: Implications for squid fisheries in the north-west Pacific Ocean. International Journal of Remote Sensing, 2010, 31(17/18): 4515-4530. |
| [10] | Feng Y J, Chen X J, Yang X M, Gao F. HSI modeling and intelligent optimization for fishing ground forecasts using a genetic algorithm. Acta Ecologica Sinica, 2013, doi: 10.5846/stxb201212171813. |
| [11] | Fan W, Cui X S, Wu Y M, Zhou W F, Chen X Z. Key technologies of operational application in fishing ground analysis and forecasting. Journal of Fishery Sciences of China, 2013, 20(1): 234-241. |
| [12] | Xu H L, Ma Z H, Qiao X T, Yang Y. Status of research and application of Geographic Information System (GIS) in Chinese marine fishery. Marine Science Bulletin, 2012, 31(1): 113-119. |
| [13] | Shao Q Q, Zhou C H, Shen X Q, Yang C J. Operational GIS and RS technology & methods for marine fishery. Journal of Remote Sensing, 2003, 7(3): 194-200. |
| [14] | Su F Z, Du Y Y, Yang X M, Liu B G. Geo-association rule with spatiotemporal reasoning. Geo-information Science, 2004, 6(4): 66-70. |
| [15] | Yang X M, Dai X J, Zhu G P. Geostatistical analysis of spatial heterogeneity of yellowfin tuna (Thunnus albacares) purse seine catch in the western Indian Ocean. Acta Ecologica Sinica, 2012, 32(15): 4682-4690. |
| [16] | Yang X M, Dai X J, Tian S Q, Zhu G P. Hot spot analysis and spatial heterogeneity of skipjack tuna (katsuwonus pelamis) purse seine resources in the western and central Pacific Ocean. Acta Ecologica Sinica, 2013, doi: 10.5846/stxb201211101579. |
| [17] | Niu M X. Spatiotemporal Dynamics of Anchovy Population and Its Response to Environmental Factors in the Yellow Sea Based on Ocean RS and GIS . Taian: Shandong Agricultural University, 2012. |
| [18] | Getis A, Ord J K. The analysis of spatial association by use of distance statistics. Geographical Analysis, 1992, 24(3): 189-206. |
| [19] | Anselin L. Local indicators of spatial association-LISA. Geographical Analysis, 1995, 27(2): 93-115. |
| [20] | Zhang S L, Zhang K. Contrast study on Moran and Getis-Ord indexes of local spatial autocorrelation indices. Journal of Geodesy and Geodynamics, 2007, 27(3): 31-34. |
| [21] | Zhang S L, Zhang K. Comparison between general Moran's Index and Getis-Ord General G of spatial autocorrelation. Acta Scientiarum Naturalium Universitatis Sunyatseni, 2007, 46(4): 93-97. |
| [22] | Tian S Q, Chen Y, Chen X J, Xu L X, Dai X J. Impacts of spatial scales of fisheries and environmental data on catch per unit effort standardization. Marine and Freshwater Research, 2009, 60(12): 1273-1284. |
| [23] | Longley P A, Goodchild M, Maguire D J, Rhind D W. Geographic Information Systems and Science 3ed ed. New York: Wiley, 2009. |
| [24] | Mitchell A. The ESRI Guide to GIS Analysis (Volume 2). California: ESRI Press, 2005. |
| [25] | Wu J G. Landscape Ecology: Pattern, Process, Scale and Hierarchy. Beijing: Higher Education Press, 2007. |
| [26] | McGarigal K, Cushman S A, Ene E. FRAGSTATS v4: Spatial pattern analysis program for categorical and continuous maps. Computer Software Program Produced by the Authors at the University of Massachusetts, Amherst. Available at the following web site: http://www.umass.edu/landeco/research/fragstats/fragstats.html, 2012 (accessed 10 May 2013). |
| [27] | Feng Y J, Han Z. RS and GIS derived spatio-temporal evolution of water landscape in coastal areas: A case study of Shanghai section on the northern Bank of Hangzhou Bay. Remote Sensing for Land and Resources, 2011, 23(1): 123-127. |
| [28] | Feng Y J, Liu Y, Han Z. Land use simulation and landscape assessment by using genetic algorithm based on cellular automata under different sampling schemes. Chinese Journal of Applied Ecology, 2011, 22(4): 957-963. |
| [29] | Feng Y J, Han Z. Impact of neighbor configurations on spatially-explicit modeling results. Geographical Research, 2011, 30(6): 1055-1065. |
| [30] | Liu A L, Wang P F, Ding Y Y. An Introduction to Geostatistics. Being: Science Press of China, 2012. |
| [1] | 陈新军, 田思泉, 陈勇, 曹杰, 马金, 李思亮, 刘必林. 北太平洋柔鱼渔业生物学. 北京: 科学出版社, 2011. |
| [4] | 刘维, 林昭进, 江艳娥, 黄梓荣. 南海北部陆架区底层渔业资源的空间分布特征. 热带海洋学报, 2011, 30(5): 95-103. |
| [5] | 李圣法, 严利平, 李惠玉, 李建生, 程家骅. 东海区头足类群聚空间分布特征的初步研究. 中国水产科学, 2006, 13(6): 936-944. |
| [6] | 陈新军, 许柳雄, 田思泉. 北太平洋柔鱼资源与渔场的时空分析. 水产学报, 2003, 27(4): 334-342. |
| [7] | 王增焕, 李纯厚, 贾晓平. 应用初级生产力估算南海北部的渔业资源量. 海洋水产研究, 2005, 26(3): 9-15. |
| [8] | 杨胜龙, 张禹, 张衡, 王栋, 马军杰, 张晶, 崔雪森. 热带印度洋黄鳍金枪鱼渔场时空分布与温跃层的关系. 生态学报, 2012, 32(3): 671-679. |
| [10] | 冯永玖, 陈新军, 杨晓明, 高峰. 基于遗传算法的渔情预报HSI建模与智能优化. 生态学报, 2013, doi: 10.5846/stxb201212171813. |
| [11] | 樊伟, 崔雪森, 伍玉梅, 周为峰, 陈雪忠. 渔场渔情分析预报业务化应用中的关键技术探讨. 中国水产科学, 2013, 20(1): 234-241. |
| [12] | 徐海龙, 马志华, 乔秀亭, 杨翼. 我国海洋渔业地理信息系统发展现状. 海洋通报, 2012, 31(1): 113-119. |
| [13] | 邵全琴, 周成虎, 沈新强, 杨崇俊. 海洋渔业遥感地理信息系统应用服务技术和方法. 遥感学报, 2003, 7(3): 194-200. |
| [14] | 苏奋振, 杜云艳, 杨晓梅, 刘宝银. 地学关联规则与时空推理的渔业分析应用. 地球信息科学, 2004, 6(4): 66-70. |
| [15] | 杨晓明, 戴小杰, 朱国平. 基于地统计分析西印度洋黄鳍金枪鱼围网渔获量的空间异质性. 生态学报, 2012, 32(15): 4682-4690. |
| [16] | 杨晓明, 戴小杰, 田思泉, 朱国平. 中西太平洋鲣鱼围网渔业资源的空间热点和空间异质性分析. 生态学报, 2013, doi: 10. 5846/stxb201211101579. |
| [17] | 牛明香. 基于海洋遥感和GIS的黄海鳀鱼种群时空动态及对海洋环境因子的响应[D]. 泰安: 山东农业大学, 2012. |
| [20] | 张松林, 张昆. 空间自相关局部指标Moran指数和G系数研究. 大地测量与地球动力学, 2007, 27(3): 31-34. |
| [21] | 张松林, 张昆. 全局空间自相关Moran指数和G系数对比研究. 中山大学学报: 自然科学版, 2007, 46(4): 93-97. |
| [25] | 邬建国. 景观生态学——格局、过程、尺度与等级. 北京: 高等教育出版社, 2007. |
| [27] | 冯永玖, 韩震. 基于遥感和GIS 的海岸带水域生态景观格局演变研究—以杭州湾北岸上海市段为例. 国土资源遥感, 2011, 23(1): 123-127. |
| [28] | 冯永玖, 刘艳, 韩震. 不同样本方案下遗传元胞自动机的土地利用模拟及景观评价. 应用生态学报, 2011, 22(4): 957-963. |
| [29] | 冯永玖, 韩震. 元胞邻域对空间直观模拟结果的影响. 地理研究, 2011, 30(6): 1055-1065. |
| [30] | 刘爱利, 王培法, 丁园圆. 地统计学概论. 北京: 科学出版社, 2012. |