 2014, Vol. 34
2014, Vol. 34文章信息
- 李凡, 徐炳庆, 马元庆, 吕振波, 王田田
- LI Fan, XU Bingqing, MA Yuanqing, LÜ Zhenbo, WANG Tiantian
- 莱州湾鱼类群落同功能种团的季节变化
- Seasonal changes of functional guilds of fish community in Laizhou Bay, East China
- 生态学报, 2014, 34(7): 1736-1745
- Acta Ecologica Sinica, 2014, 34(7): 1736-1745
- http://dx.doi.org/10.5846/stxb201305060948
-
文章历史
- 收稿日期:2013-5-6
- 修订日期:2013-11-6






同功能种团(Functional guilds)是指在系统中起着相似生态作用、并占据相近或相似生态位的若干物种集合体,其划分通常是依据物种的摄食习性,把食性相近的物种进行集合,以显示它们在生态系统物质循环和能量流动中所起到的不同作用[1, 2, 3]。该方法把生物类群划分为利用相同资源基础(种团) 或具有共同功能特征(功能群)的类群,在生态学研究中简化了群落内物种之间的关系,弱化了物种的个别作用,减小了生态系统的研究的复杂性,同时可以增加群落研究的稳定性和可预测性[4, 5]。
应用同功能种团方法对鱼类群落的研究见于姜亚洲等[2对东海北部鱼类群落同功能种团的研究,吕振波等[6]对黄海山东海域鱼类群落同功能种团的研究;应用功能群对鱼类群落的研究见于张波等对东海[7]、黄海[8]、长江口海域[9]、渤海[10]鱼类群落或高营养层次生物功能群的研究以及Shan等[11]对东海陆架鱼类群落功能群的研究,应用功能群对莱州湾大型底栖动物群落的研究[12]也有报道,但未见应用同功能种团或功能群方法研究莱州湾鱼类群落的文献报道。本文利用2011年4季度月调查资料,开展同功能种团及其季节变化研究,并根据稳定同位素测定结果计算各功能种团营养级,为开展莱州湾食物网和营养动力学研究提供基础数据。
数据来自2011年春季(5月)、夏季(8月)、秋季(10月)和冬季(12月)莱州湾(119°05′—120°00′E、37°12′—37°40′N)(图 1)的调查资料。调查站位20个,调查船只功率260 kW,调查网具为单船底拖网,网口周长30.6 m,囊网网目20 mm。每站拖曵1 h,拖速3.5 kn。拖曳时,网口宽度约8 m。调查均在白天进行。采集样品冰鲜保存,带回实验室进行分类和生物学测定。记录每一种类的数量和重量,将其换算为单位时间的生物量(kg/h)和丰度(尾/h)。样品采样及分析均按《海洋监测规范第4部分:海水分析》(GB 17378.4—2007)和《海洋调查规范第6部分:海洋生物调查》(GB/T 12763.6—2007)执行。鱼类名称及分类地位以Nelson分类系统[13]为依据,并参考FAO Fishbase数据库[14]。

|
| 图 1 调查站位 Fig. 1 Sample stations |
同位素待测样品带回实验室后解冻后用蒸馏水洗涤,取适量背部白肌肉用于分析。所有样品-80℃冷冻后在冷冻干燥机(亿倍 YB-FD-1)中真空干燥,用石英研钵充分磨匀并加入1 mol/dm3的HCL酸化处理后再烘干,最后装入离心管中,并放入干燥器中保存以备稳定同位素分析。实验样品的稳定同位素比值由中国林业科学院稳定同位素实验室测定。
1.2 数据分析(1)数据标准化
根据各站位拖网时间和各鱼种的可捕系数,对调查数据进行标准化[15]。中上层鱼类可捕系数取0.3,近底层鱼类取0.5,底层鱼类取0.8。
(2)同功能种团划分
同功能种团的划分依据鱼类的摄食习性[2],参考杨纪明[16]、邓景耀等[17, 18]、张波等[10]和伍汉霖等[19]的文献资料。
(3)物种多样性指数和同功能种团多样性指数
物种多样性指数和同功能种团多样性指数均使用Wilhm[20]改进的Shannon-Wiener指数,计算式为:
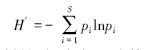
计算物种多样性时,式中pi为第i种渔获物生物量占总生物量的比例;计算同功能种团多样性时,pi为第i种同功能种团生物量占总生物量的比例。
(4)等级聚类与NMDS分析
为平衡不同种类在群落中的作用,将生物量数据进行平方根处理,并建立种类×站位矩阵(Matrix),利用PRIMER 6软件非加权组平均法(Unweighted pair group method using arithmetic averages)进行等级聚类(Cluster)和非度量多维标度(Non-metric multidimensional scaling,NMDS),NMDS的优劣使用协强系数(Stress)判定。
(5)营养级
利用稳定氮同位素评估营养级,将初级生产者营养级定为1,则营养级的计算公式为[21]:

式中,TL表示计算生物的营养级;δ15Nconsumer指系统中消费者氮同位素比值;δ15Nbaseline为该系统基线生物的氮同位素比值,本研究取渤海浮游动物氮同位素测定值6.14‰[22];Δδ15N为1个营养级的氮同位素富集度,本研究采用有关文献[23]采用的氮同位素值3.4‰。
(6)数据处理和图件绘制
多样性计算、等级聚类分析使用Primer 6,CCA分析使用Canoco 4.5软件。图件绘制使用ArcGIS 9、Excel 2010、Origin 8、Primer 6和Canoco 4.5完成。
2 结果与分析 2.1 同功能种团种类组成全年调查共渔获鱼类49种,根据鱼类摄食习性,可划分为7个同功能种团,其中底栖动物食性种团(FG3)种类数最多(14种),底栖动物/游泳动物食性种团(FG4)次之(12种),碎屑食性种团(FG7)种类最少,仅2种(表 1)。各功能种团种类数组成的季节变化趋势基本同全年组成一致,值得注意的是杂食性种团在冬季未出现。
| 序号 No. | 种类 Species | 同功能种团 Functional guild | 调查时间Survey time | |||
| 春季 Spring | 夏季 Summer | 秋季 Autumn | 冬季 Winter | |||
| FG1:浮游动物食性种团Planktivores;FG2:浮游动物/底栖动物食性种团Planktivores/Benthivores;FG3:底栖动物食性种团Benthivores;FG4:底栖动物/游泳动物食性种团Benthivores/Piscivores;FG5:游泳动物食性种团Piscivores;FG6:杂食性种团Omnivores;FG7:碎屑食性种团Detritivores | ||||||
| 1 | 鳀Engraulis japonicus | FG1 | √ | √ | √ | |
| 2 | 黄鲫Setipinna taty | FG1 | √ | √ | √ | |
| 3 | 赤鼻棱鳀Thryssa kammalensis | FG1 | √ | √ | √ | |
| 4 | 中颌棱鳀Thryssa mystax | FG1 | √ | √ | √ | |
| 5 | 斑鰶Konosirus punctatus | FG7 | √ | √ | √ | |
| 6 | 青鳞小沙丁鱼Sardinella zunasi | FG1 | √ | √ | √ | |
| 7 | 安氏新银鱼Neosalanx anderssoni | FG1 | √ | √ | ||
| 8 | 长蛇鲻Saurida elongata | FG5 | √ | √ | ||
| 9 | 鮻Liza haematocheilus | FG7 | √ | |||
| 10 | 莫氏海马Hippocampus coronatus | FG1 | √ | √ | √ | √ |
| 11 | 尖海龙Syngnathus acus | FG1 | √ | √ | √ | |
| 12 | 许氏平鲉Sebastes schlegelii | FG4 | √ | √ | ||
| 13 | 鲬Platycephalus indicus | FG4 | √ | √ | √ | |
| 14 | 大泷六线鱼Hexagrammos otakii | FG4 | √ | √ | √ | |
| 15 | 松江鲈Trachidermus fasciatus | FG5 | √ | √ | ||
| 16 | 网纹狮子鱼Liparis chefuensis | FG5 | √ | √ | ||
| 17 | 花鲈Lateolabrax japonicus | FG4 | √ | √ | √ | |
| 18 | 细条天竺鲷Apogon lineatus | FG2 | √ | |||
| 19 | 多鳞鱚Sillago sihama | FG3 | √ | √ | ||
| 20 | 黑棘鲷Acanthopagrus schlegelii | FG4 | √ | √ | ||
| 21 | 真鲷Pagrus major | FG5 | √ | |||
| 22 | 皮氏叫姑鱼Johnius belangerii | FG4 | √ | √ | √ | |
| 23 | 小黄鱼Larimichthys polyactis | FG6 | √ | √ | √ | |
| 24 | 银姑鱼Pennahia argentata | FG4 | √ | √ | ||
| 25 | 绵鳚Zoarces elongatus | FG3 | √ | |||
| 26 | 方氏锦鳚Pholis fangi | FG2 | √ | √ | √ | |
| 27 | 绯衔Callionymus beniteguri | FG3 | √ | √ | √ | √ |
| 28 | 黄鳍刺虾虎鱼Acanthogobius flavimanus | FG3 | √ | √ | ||
| 29 | 斑尾刺虾虎鱼Acanthogobius ommaturus | FG4 | √ | √ | √ | √ |
| 30 | 乳色刺虾虎鱼Acanthogobius lactipes | FG3 | √ | |||
| 31 | 六丝钝尾虾虎鱼Amblychaeturichthys hexanema | FG3 | √ | √ | √ | |
| 32 | 矛尾虾虎鱼Chaeturichthys stigmatias | FG4 | √ | √ | √ | √ |
| 33 | 中华栉孔虾虎鱼Ctenotrypauchen chinensis | FG3 | √ | √ | √ | |
| 34 | 裸项蜂巢虾虎鱼Favonigobius gymnauchen | FG6 | √ | |||
| 35 | 普氏细棘虾虎鱼Acentrogobius pflaumii | FG3 | √ | √ | √ | √ |
| 36 | 长丝虾虎鱼Myersina filifer | FG6 | √ | √ | ||
| 37 | 拉氏狼牙虾虎鱼Odontamblyopus lacepedii | FG3 | √ | √ | ||
| 38 | 髭缟虾虎鱼Tridentiger barbatus | FG3 | √ | √ | √ | √ |
| 39 | 纹缟虾虎鱼Tridentiger trigonocephalus | FG3 | √ | √ | ||
| 40 | 小带鱼Eupleurogrammus muticus | FG6 | √ | √ | ||
| 41 | 蓝点马鲛Scomberomorus niphonius | FG6 | √ | √ | ||
| 42 | 银鲳Pampus argenteus | FG2 | √ | √ | ||
| 43 | 褐牙鲆Paralichthys olivaceus | FG5 | √ | √ | ||
| 44 | 石鲽Kareius bicoloratus | FG3 | √ | √ | ||
| 45 | 钝吻黄盖鲽Pseudopleuronectes yokohamae | FG3 | √ | √ | ||
| 46 | 短吻红舌鳎Cynoglossus joyneri | FG3 | √ | √ | √ | √ |
| 47 | 半滑舌鳎Cynoglossus semilaevis | FG4 | √ | |||
| 48 | 淡鳍兔头鲀Gastrophysus lunaris | FG4 | √ | |||
| 49 | 假睛东方鲀Takifugu pseudommus | FG4 | √ | √ | √ | |
以生物量计,碎屑食性种团(FG7)在全年处于第1优势地位,占32.8%;其次为浮游动物食性种团(FG1),占31.6%;再次为底栖动物/游泳动物食性种团(FG4),占25.0%;浮游动物/底栖动物食性种团(FG2)、底栖动物食性种团(FG3)、游泳动物食性种团(FG5)和杂食性种团(FG6)所占比例均低于1%(图 2)。

|
| 图 2 莱州湾鱼类各同功能种团生物量组成。代码同表 1 Fig. 2 Biomass composition of different functional guilds of fishes in Laizhou bay FG1:浮游动物食性种团;FG2:浮游动物/底栖动物食性种团;FG3:底栖动物食性种团;FG4:底栖动物/游泳动物食性种团;FG5:游泳动物食性种团;FG6:杂食性种团;FG7:碎屑食性种团 |
同功能种团的生物量组成存在明显的季节变化(图 3),浮游动物食性种团(FG1)在春季和秋季均占绝对优势(主要贡献者为赤鼻棱鳀Thryssa kammalensis、青鳞小沙丁鱼Sardinella zunasi等);在夏季,由于群落第1优势种斑鰶(Konosirus punctatus)的生物量比例达54.0%,因此碎屑食性种 团占优势;冬季则为底栖动物/游泳动物食性种团(FG4)占优势(主要贡献者为矛尾虾虎鱼Chaeturichthys stigmatias和斑尾刺虾虎鱼Acanthogobius ommaturus)。杂食性种团生物量仅在夏季有一定比例(11.7%),在其他季节其生物量比例极低。

|
| 图 3 莱州湾鱼类各功能种团生物量和组成百分比的季节变化 Fig. 3 Seasonal changes of biomass and biomass composition of different functional guilds of fishes in Laizhou bay |
以占全年调查尾数97.4%的15种类(表 4)计算,各同功能种团平均营养级分别为:浮游动物食性种团3.71、浮游动物/底栖动物食性种团3.83、底栖动物食性种团3.43、底栖动物/游泳动物食性种团3.75、游泳动物食性种团3.86、杂食性种团3.73、碎屑食性种团3.34。
| 种类 Species | 同功能种团 Functional guilds | 营养级 Trophic level | 尾数百分比 N/% | 测定尾数 Numbers of measured |
| 青鳞小沙丁鱼Sardinella zunasi | 浮游动物食性种团(FG1) | 3.52 | 35.7 | 47 |
| 赤鼻棱鳀Thryssa kammalensis | 浮游动物食性种团(FG1) | 3.97 | 24.6 | 22 |
| 黄鲫Setipinna taty | 浮游动物食性种团(FG1) | 3.76 | 1.7 | 17 |
| 银鲳Pampus argenteus | 浮游动物/底栖动物食性种团(FG2) | 3.83 | 0.003 | 7 |
| 短吻红舌鳎Cynoglossus joyneri | 底栖动物食性种团(FG3) | 3.67 | 0.7 | 52 |
| 绯衔Callionymus beniteguri | 底栖动物食性种团(FG3) | 3.32 | 1.6 | 37 |
| 矛尾虾虎鱼Chaeturichthys stigmatias | 底栖动物/游泳动物食性种团(FG4) | 3.74 | 14.1 | 55 |
| 斑尾刺虾虎鱼Acanthogobius ommaturus | 底栖动物/游泳动物食性种团(FG4) | 3.65 | 2.8 | 47 |
| 鲬Platycephalus indicus | 底栖动物/游泳动物食性种团(FG4) | 3.91 | 0.2 | 50 |
| 银姑鱼Pennahia argentata | 底栖动物/游泳动物食性种团(FG4) | 3.95 | 0.9 | 33 |
| 皮氏叫姑鱼Johnius belangerii | 底栖动物/游泳动物食性种团(FG4) | 4.09 | 0.6 | 7 |
| 褐牙鲆Paralichthys olivaceus | 游泳动物食性种团(FG5) | 4.11 | 0.01 | 8 |
| 蓝点马鲛Scomberomorus niphonius | 杂食性种团(FG6) | 3.74 | 0.3 | 20 |
| 小黄鱼Larimichthys polyactis | 杂食性种团(FG6) | 3.62 | 0.03 | 27 |
| 斑鰶Konosirus punctatus | 碎屑食性种团(FG7) | 3.34 | 15.4 | 44 |
同功能种团多样性(HFD)和种类多样性(Hs)季节变化趋势相同,均在夏季达到最高值,冬季达到最低值,但同功能种团多样性均低于该季节的种类多样性(图 4),同功能种团多样性和种类多样性的关系使用指数方程(R2=0.8064)拟合效果最佳(图 5),其次为线性方程(R2=0.7587)。同功能种团多样性和种类多样性均为冬季和春季、冬季和夏季存在显著差异(P<0.05),其它季节间无显著差异(P>0.05)。

|
| 图 4 莱州湾鱼类的种类多样性和同功能种团多样性 Fig. 4 Species and functional guilds diversity indices of fishes in Laizhou Bay |

|
| 图 5 莱州湾鱼类的种类多样性和同功能种团多样性的关系 Fig. 5 Relationship between species diversity and functional guilds diversity of fishes in Laizhou Bay |
浮游动物/底栖动物食性种团(FG2)和游泳动物食性种团(FG5)两者聚为1组,共同特点是种类 少,生物量低,率先与其他同功能种团分离。浮游动物食性种团(FG1)和碎屑食性种团(FG7)聚为1组,共同特点是生物量高且种类数并不太高。底栖动物食性种团(FG3)和底栖动物/游泳动物食性种团(FG4)种类较多,两者聚为1组。FG6在冬季未出现,独自聚类为1组。NMDS排序显示协强系数(Stress)为0.01,表明排序结果完全可信。
对所有调查站位的聚类和NMDS排序结果显示,夏季、秋季的调查站位各自聚在一起,并且和其他调查区隔明显;春季、冬季调查站位聚类较分散,镶嵌在一起(图 7)。

|
| 图 6 基于欧氏距离的同功能种团聚类图(a)和NMDS排序图 (数据经log(X+1)转换) Fig. 6 The hierarchical cluster dendrogram (a) and the 2-dimentional MDS ordinal configuration (b) of functional guilds base on the Euclidean distance (Data was log(x+1) transformed) |

|
| 图 7 调查站位等级聚类图 (a) 和NMDS 排序图 (b) Fig. 7 hierarchical cluster dendrogram (a) and the 2-dimentional MDS ordinal configuration (b) of survey stations |
图 8为同功能种团与环境变量的双轴排序图,浮游动物生物量、浮游植物细胞密度以及无机磷浓度是影响第一轴的主要变量,大型底栖动物生物量和水深是影响第二轴的主要变量。

|
| 图 8 同功能种团与环境变量之间的双轴排序图 Fig. 8 Ordination biplots of functional guilds and environmental variables FG1—FG7代码同表 1; Deep:水深;DIN:总无机氮浓度;DIP:活性磷酸盐浓度;Chla:叶绿素a浓度;Plankton:浮游植物细胞密度;Zooplank:浮游动物生物量;Benthos:底栖动物生物量 |
根据鱼类食性划分,本研究海域鱼类群落包含7个同功能种团。姜亚洲等[2]在对东海北部鱼类群落的研究中,考虑软骨鱼类游泳速度慢,将软骨鱼类单独划为一个种团,共8个同功能种团。在莱州湾,2006年至2011年历次调查中均未发现软骨鱼类,莱州湾的软骨鱼类已失去作为一个独立功能种团的意义,在黄海山东海域也缺少软骨鱼类这一种团[6]。 张波等[10]根据2009年调查认为小黄鱼(Larimichthys polyactis)是渤海主要的饵料种类之一,同时由于其幼体和成体食性差异较大,认为小黄鱼是影响渤海鱼类功能群划分的主要种类。而根据本调查,小黄鱼仅列相对重要性指数排序的第35位(IRI=3.4<10,为少见种),不是2011年莱州湾主要种类,故认为其对莱州湾鱼类同功能种团划分影响较小。
目前莱州湾鱼类群落中浮游动物食性种团、底栖动物/游泳动物食性种团、碎屑食性种团所占比例接近九成(89.4%)。与1980年代调查结果相比,目前碎屑食性功能种团比例明显上升,主要是因为营碎屑食性的斑鰶比例大幅增高,许思思等[24]对1990年代渤海鱼类群落的研究也有相似的结果。斑鰶既摄食碎屑,也可摄食浮游动物、浮游植物,摄食弹性大,在海洋环境变化过程中,适应性较广,同时由于大型肉食性鱼类的衰退减轻了对斑鰶的捕食压力,而鳀(Engraulis japonicus)、黄鲫(Setipinna taty)、银鲳(Pampus argenteus)等浮游动物食性种类资源的大幅衰退减轻了对浮游动物、浮游植物的食物竞争,部分碎屑食性的种类如棘皮动物的减少[25]也减轻了碎屑的食物竞争,这些变化均为斑鰶种群规模增加[26, 27, 28]创造了有利条件。与1980年代、1990年代相比,游泳动物食性种团种类资源大幅下降,其或已基本灭绝(如鮸 Miichthys niiu、斑鳐Okamejei kenojei)或资源量已极低(如带鱼Trichiurus lepturus、褐牙鲆Paralichthys olivaceus、许氏平鲉Sebastes schlegelii),或者食性已发生转变(如蓝点马鲛Scomberomorus niphonius)[10],导致目前莱州湾游泳动物食性种团比例极低。需要指出的是,本研究在进行鱼类同功能种团划分时,依据的是其成体的摄食习性。考虑部分鱼类,如小黄鱼等,其幼体和成体食性差异较大,幼体更喜好摄食浮游动物。特别是夏季,幼鱼比例高,浮游动物食性种团所占比例理应高于本研究的结果。
CCA排序表明,浮游动物食性种团、浮游动物/底栖动物食性种团生物量与浮游动物生物量、无机氮浓度、叶绿素a浓度、浮游植物细胞密度的正相关;底栖动物食性种团、底栖动物/游泳动物食性种团、游泳动物食性种团生物量与底栖动物生物量、水深正相关,与浮游动物生物量、浮游植物细胞密度负相关;杂食性种团、碎屑食性种团生物量与浮游动物生物量、浮游植物密度、无机磷浓度相关度较高,与无机氮浓度和底栖动物生物量也有一定相关度。
种类多样性与同功能种团多样性相互关系分析表明,同功能种团多样性随种类多样性增加而增加,指数方程和线性方程均能较好拟合其相互关系。姜亚洲等[2]对东海北部鱼类群落的研究和李少文等[12]对莱州湾大型底栖动物群落研究发现直线方程拟合同功能种团多样性和种类多样性效果最佳;本研究中,指数方程拟合效果(R2=0.8064)略好于直线方程(R2=0.7587)。在多数站位,种类多样性和同功能种团多样性均处于较低水平,低于马克平和刘玉明[29]提出的多样性指数合理范围(1.5—3.5)。
在一个稳定的(未受扰动的)生态系统中,通常生物种类较多,食物网结构复杂,同功能种团由多个可以相互替代的种类构成,物质循环与能量流动渠道多。本研究种不同功能种团在种类数、生物量上巨大悬殊,及其季节间的剧烈波动,表现为等级聚类和NMDS排序区隔明显(图 6,图 7)。目前莱州湾鱼类同功能种团在种类和生物量上季节间的巨大差异以及较低的同功能种团多样性也从一个侧面表明群落结构的不稳定性,应引起足够的重视。
3.2 营养级营养级是揭示生态系统结构和功能的基础[4],群落平均营养级是衡量生物多样性水平的8个指标之一[30]。营养级取决于该种物组成,一般来说,高营养级(肉食性)鱼类通常为大型鱼类。在一个未受干扰的群落中,大个体的种类(高营养级)占较大比例,群落的平均营养级相应较高。在过度捕捞影响下,大量顶级捕食者被移除的同时其摄食对象(低营养级种类)由于被摄食压力降低而导致种群规模增加,群落平均营养级下降。Pauly等[31]报导1950—1994年间全球渔获物的平均营养级从3.3下降到3.1,Zhang和Lee[32]报道1967—2002年间黄海渔获物营养级以年均0.01的水平持续下降。
本研究中7个同功能种团的均营养级差距较小,均在3—4之间,处于低级肉食者水平[17]。与杨纪明[16]根据20世纪80年代和90年代渤海调查资料研究的结果相比(表 3),目前底栖动物食性种团和游泳动物食性种团的平均营养级明显下降,碎屑食性种团平均营养级有所上升,浮游动物食性种团平均营养级变化较小。
| 同功能种团 Functional guild | 平均营养级Average trophic level | |
| 本文This paper | 杨纪明,2000 | |
| FG1 | 3.71 | 3.5 |
| FG2 | 3.83 | |
| FG3 | 3.43 | 4.1 |
| FG4 | 3.75 | |
| FG5 | 3.86 | 4.6 |
| FG6 | 3.73 | |
| FG7 | 3.34 | 2.4 |
底栖动物食性种团平均营养级的下降可能与偏好摄食较高营养级的短尾类、长尾类(营养级2.8左右)的种类(如石首科鱼类,石鲽Kareius bicoloratus等)比例下降,而偏好摄食较低营养级的涟虫类、端足类(营养级2.1—2.3)的种类(绯 鱼 衔 Callionymus beniteguri、短吻红舌鳎Cynoglossus joyneri等)比例增加有一定关系;同时杨纪明等[16]的研究中并未区分底栖动物食性和底栖动物/游泳动物食性,将底栖动物/游泳动物食性鱼类(如银姑鱼)也划归底栖动物食性,这些种类营养级通常稍高于底栖动物食性鱼类。这些因素均导致目前计算的底栖动物食性种团平均营养级偏低,但底栖动物食性种团平均营养级的下降已是不争的事实。
莱州湾碎屑食性种团主要以斑鰶为主,对于其食性,不同文献报道并不相同。唐启升和叶懋中[33]认为斑鰶为浮游生物食性,主要摄食浮游植物(圆筛藻、根管藻等)和浮游动物(纺锤水蚤、剑水蚤等);杨纪明[16]认为斑鰶主要摄食腐屑(60.0%),也摄食动物性饵料(36.5%)和少量的植物性饵料(3.5%),并据此计算斑鰶营养级为2.4;而郭学武等[26]认为斑鰶食性有较大弹性,沉积碎屑是其食物重要组成部分。本研究中,根据稳定同位素测定结果计算得出斑鰶的营养级范围为2.71—4.29,且出现营养级随叉长增加而变小的现象,在进行功能种团划分时,仍将斑鰶视为碎屑食性种团,同时认为斑鰶摄食较大比例的动物性饵料。
浮游动物食性种团平均营养级较高(甚至高于底栖动物食性种团),可能与本研究中将赤鼻棱鳀(平均养级为3.97)划归为浮游动物食性种团有关。相关研究报道[10, 16, 17, 33]均认为赤鼻棱鳀为浮游动物食性,但也摄食较高比例的底栖虾类[10],而本文中仍将其归于浮游动物食性。
目前莱州湾游泳动物食性鱼类主要是1龄和当年生幼鱼,同位素测定样品也以1龄和当年生个体为主,可能与使用成体测定的结果有所偏差,但代表着目前鱼类群落平均营养级水平。本文营养级测定采用的是稳定同位素法,而杨纪明等[16]的研究采用的是胃含物分析法,不同方法测定值可能存在一定的误差,下一步应结合胃含物分析法,同时测定各种类可能的摄食对象的稳定同位素特征,进一步提高营养级及同功能种团划分的精确度。
| [1] | Cheng J H, Jiang Y Z. Methods for evaluating fishing effects on fish community. Journal of Fishery Sciences of China, 2008, 15(2): 359-366. |
| [2] | Jiang Y Z, Cheng J H, Li S F. Variation in fi sh community structure and biodiversity in the north of the East China Sea between two periods. Journal of Fishery Sciences of China, 2008, 15(3): 453-459. |
| [3] | Munoz A A, Ojeda F E. Guild structure of carnivorous intertidal fishes of the Chilean coast: implications of ontogenetic dietary shifts. Oecologia, 1998, 114: 563-573. |
| [4] | Ji W W, Li S F, Chen X Z. Application of fish trophic level in marine ecosystem. Journal of Fishery Sciences of China, 2010, 17(4): 878-887. |
| [5] | Steneck R S, Watling L. Feeding capabilities and limitations of herbivorous molluscs: A functional group approach. Marine Biology, 1982, 68: 299-319. |
| [6] | Lv Z B, Li F, Xu B Q, Wang B. Fish community diversity during spring and autumn in the Yellow Sea off the coast of Shandong. Biodiversity Science, 2012, 20(2): 207-214. |
| [7] | Zhang B, Tang Q S, Jin X S. Functional groups of fish assemblages and their major species at high trophic level in the East China Sea. Journal of Fishery Sciences of China, 2007, 14(6): 939-949. |
| [8] | Zhang B, Tang Q S, Jin X S. Functional groups of communities and their major species at high trophic level in the Yellow Sea ecosystem. Ecologica Sinica, 2009, 29(3):1109-1111. |
| [9] | Zhang B, Jin X S, Tang Q S. Functional groups of high trophic level communities in adjacent waters of Changjiang estuary. Chinese Journal of Applied Ecology, 2009, 20(2): 344-351. |
| [10] | Zhang B, Li Z Y, Jin X S. Functional groups of fish assemblages and their major species in the Bohai Sea. Journal of Fisheries of China, 2012, 36(1): 64-72. |
| [11] | Shan X J, Jin X S, Zhou Z P, Dai F Q. Fish community diversity in the middle continental shelf of the East China Sea. Chinese Journal of Oceanology and Limnology, 2011, 29(6): 1199-1208. |
| [12] | Li S W, Liu Y J, Li F, Zhang Y, Xu Z F, Lv Z B, Wang T T, Zhang A B. Macrobenthic functional groups in Laizhou Bay,East China. Chinese Journal of Ecology, 2013, 32(2): 380-388. |
| [13] | Nelson J S. Fishes of the World, 4th edn. John Wiley & Sons Inc, New Jersey, 2006. |
| [14] | Froese R, Pauly D. FishBase. (2011). http://www.fishbase.org. |
| [15] | Zhan B Y. Fish stock assessment. BeiJing: Agriculture Press, 2000. |
| [16] | Yang J M. A Study on Food and Trophic Levels of Baohai Sea Fish. Modern Fisheries Information, 2001, 16(10): 10-19. |
| [17] | Deng J Y, Meng T X, Ren S M. Food web of fishes in Bohai Sea. Acta Ecologica Sinica, 1986, 6(4): 356-364. |
| [18] | Deng J Y, Jiang W M, Yang J M,Li J. Species interaction and food web of major predatory species in the Bohai Sea. Journal of fishery sciences of China, 1997, 4(4): 1-7. |
| [19] | Wu H L, Zhong J S. Chinese animal: Osteichthyes, Perciformes, Gobioidei. Beijing: Science Press, 2008. |
| [20] | Wilhm J L. Use of biomass units in Shannon's formula. Ecology, 1968, 49(1): 153-156. |
| [21] | Post D M. Using stable isotopes to estimate trophic position: models, methods and assumptions. Ecology, 2002, 83(3): 703-718. |
| [22] | Ji W W, Chen X Z, Jiang Y Z, Liu Y, Hu F, Li S F. Stable isotope analysis of some representative nektonic organisms in the central and northern part of East China Sea. Marine Fisheries, 2011,33(3): 241-250. |
| [23] | Davenport S R, Bax N J. A trophic study of a marine ecosystem off southern Australia using stable isotopes of carbon and nitrogen. Canadian Journal of Fisheries and Aquatic Sciences, 2002, 59: 514-530. |
| [24] | Xu S S, Song J M, Li X G, Yuan H M, Li N, Zhang Y. Modeling of Fishing Effects on Fishery Resources and Ecosystems of the Bohai Sea. Resources Science, 2011, 33(6): 1153-1162. |
| [25] | Zhang Y, Liu Y J, Zhang Y, Xu B Q. Ecological characteristics of benthic polychaete community and its responses to environmental change in Laizhou Bay,Shandong Province of East China. Chinese Journal of Ecology, 2012, 31(4): 888-895. |
| [26] | Guo X W, Tang Q S, Sun Y, Zhang B. The consumption and ecological conversion efficiency of dotted gizzard shad(clupanodon punctatus). Marine fisherries reseach, 1999, 20(2): 17-25. |
| [27] | Jin X S, Deng J Y. Variations in community structure of fishery resources and biodiversity in the L aizhou Bay, Shandong. Biodiversity Science, 2000, 8(1): 65-72. |
| [28] | Lv Z B, Xu B Q, Li F, Song M Y, Zhang H J, Liu Y J. Structure and distribution of fish resources in the Yellow Sea off Shandong during spring and autumn 2006. Journal of Fishery Sciences of China, 2011, 18(6): 1335-1342. |
| [29] | Ma K P, Liu Y M. Measurement of biotic community diversity Iα diversity(part two). Chinese biodiversity, 1994, 2 (4): 231-239. |
| [30] | Pauly D, Watson R. Background and interpretation of the'Marine Trophic Index' as a measure of biodiversity. Phil Trans R Soc B, 2005, 360: 415-423. |
| [31] | Pauly D, Christensen V, Dalsgaard J, Froese R, Torres Jr. F. Fishing down marine food webs. Science, 1998, 279: 860-863. |
| [32] | Zhang C I, Lee S K. Trophic levels and fishing intensities in Korean marine ecosystems. J Korean Fish Soc, 2004, 6: 194-206. |
| [33] | Tang Q S, Ye M Z. Development and protection of fishery resources in Shandong offshore. Beijing: Agriculture Press, 1990. |
| [1] | 程家骅, 姜亚洲. 捕捞对海洋鱼类群落影响的研究进展. 中国水产科学, 2008, 15(2): 359-366. |
| [2] | 姜亚洲, 程家骅, 李圣法. 东海北部鱼类群落多样性及其结构特征的变化. 中国水产科学, 2008, 15(3): 453-459. |
| [4] | 纪炜炜, 李圣法, 陈雪忠. 鱼类营养级在海洋生态系统研究中的应用. 中国水产科学, 2010, 17(4): 878-887. |
| [6] | 吕振波, 李凡, 徐炳庆, 王波. 黄海山东海域春、秋季鱼类群落多样性. 生物多样性, 2012, 20(2): 207-214. |
| [7] | 张波, 唐启升, 金显仕. 东海高营养层次鱼类功能群及其主要种类. 中国水产科学, 2007, 14(6): 939-949. |
| [8] | 张波, 唐启升, 金显仕. 黄海生态系统高营养层次生物群落功能群及其主要种类. 生态学报, 2009, 29(3):1109-1111. |
| [9] | 张波, 金显仕, 唐启升. 长江口及邻近海域高营养层次生物群落功能群及其变化. 应用生态学报, 2009, 20(2): 344-351. |
| [10] | 张波, 李忠义, 金显仕. 渤海鱼类群落功能群及其主要种类. 水产学报, 2012, 36(1): 64-72. |
| [12] | 李少文, 刘元进, 李凡, 张莹, 徐宗法, 吕振波, 王田田, 张爱波. 莱州湾大型底栖动物功能群现状. 生态学杂志, 2013, 32(2): 380-388. |
| [15] | 詹秉义. 渔业资源评估. 北京: 农业出版社, 2000. |
| [16] | 杨纪明. 渤海鱼类的食性和营养级研究. 现代渔业信息, 2001, 16(10): 10-19. |
| [17] | 邓景耀, 孟田湘, 任胜民. 渤海鱼类食物关系的初步研究. 生态学报, 1986, 6(4): 356-364. |
| [18] | 邓景耀, 姜卫民, 杨纪明, 李军. 渤海主要生物种间关系及食物网的研究. 中国水产科学, 1997, 4(4): 1-7. |
| [19] | 伍汉霖, 钟俊生. 中国动物志: 硬骨鱼纲、鲈形目[五]、虾虎鱼亚目. 北京:科学出版社, 2008. |
| [22] | 纪炜炜, 陈雪忠, 蒋亚洲, 刘勇, 胡芬, 李圣法. 东海中北部游泳动物稳定碳氮同位素研究. 海洋渔业, 2011,33(3): 241-250. |
| [24] | 许思思, 宋金明, 李学刚, 袁华茂, 李宁, 张英. 渔业捕捞对渤海渔业资源及生态系统影响的模型研究. 资源科学, 2011, 33(6): 1153-1162. |
| [25] | 张莹, 刘元进, 张英, 徐炳庆, 吕振波. 莱州湾多毛类底栖生物生态特征及其对环境变化的响应. 生态学杂志, 2012, 31(4): 888-895. |
| [26] | 郭学武, 唐启升, 孙耀, 张波. 斑鰶的摄食与生态转换效率. 海洋水产研究, 1999, 20(2): 17-25. |
| [27] | 金显仕, 邓景耀. 莱州湾渔业资源群落结构和生物多样性的变化. 生物多样性, 2000, 8(1): 65-72. |
| [28] | 吕振波, 徐炳庆, 李凡, 宋明毅, 张焕君, 刘元进. 2006年春、秋季黄海山东海域鱼类资源结构与数量分布. 中国水产科学, 2011, 18(6): 1335-1342. |
| [29] | 马克平, 刘玉明. 生物群落多样性的测度方法 I α多样性的测度方法(下). 生物多样性, 1994, 2 (4): 231-239. |
| [33] | 唐启升, 叶懋中. 山东近海渔业资源开发与保护. 北京: 农业出版社, 1990. |