 2014, Vol. 34
2014, Vol. 34文章信息
- 李宁, 王征, 鲁长虎, 熊天石, 傅文源, 吴锦平
- LI Ning, WANG Zheng, LU Changhu, XIONG Tianshi, FU Wenyuan, WU Jinping
- 斑块生境中食果鸟类对南方红豆杉种子的取食和传播
- Seed foraging and dispersal of Chinese yew (Taxus chinensis var. mairei) by frugivorous birds within patchy habitats
- 生态学报, 2014, 34(7): 1681-1689
- Acta Ecologica Sinica, 2014, 34(7): 1681-1689
- http://dx.doi.org/10.5846/stxb201303220485
-
文章历史
- 收稿日期:2013-3-22
- 修订日期:2013-10-21






2. 南京大学生命科学学院, 南京 210093;
3. 梅花山国家级自然保护区管理局, 龙岩 364207
2. School of Life Science, Nanjing University, Nanjing 210093, China;
3. Administration Bureau of the Meihuashan National Nature Reserve, Longyan 364207, China
人类干扰导致自然生境持续丧失,全球大量森林生态系统被人类景观所占据[1, 2, 3]。在亚热带和温带,人类的土地利用尤其强烈,房屋、农田等人为景观侵占了原始的自然生境,导致连续的自然生境被分割为斑块状,进而影响生境中动植物种子传播互惠关系[3, 4]。对于某些肉质果植物而言,生境斑块化常导致植物孤立地聚集在生境斑块中(“种源”斑块,简称源斑块),且源斑块与更新生境斑块(适宜于植物幼苗萌发的斑块)可能存在一定的隔离[5],鸟类从而成为了联系植物源斑块和更新斑块的媒介[6, 7, 8],它们的种子传播行为和传播效率决定了斑块生境中植物种群的更新效率[9, 10]。但由于斑块生境中的人为干扰较强[11, 12],抗干扰能力较强的森林泛性鸟类(Forest generalist)具有较高的出现频率,少数隐蔽能力较强的森林专性鸟类(Forest specialist)亦有活动[13]。而近年来的一些研究表明,斑块化生境中,植物依然能吸引鸟类取食并传播种子[10, 13];专性鸟类的传播距离要比泛性鸟类近,二者对植物种群更新贡献不同[10, 12, 14]。但植物的种群更新并非仅取决于种子被鸟类传播多远[15, 16],更决定于种子被鸟类散布在什么样的生境,尤其当植物的更新生境镶嵌在斑块生境中时,鸟类能否将种子散布至更新生境就对植物更新至关重要。因此,从传播距离和传播生境两方面能更全面地评估斑块生境中专性鸟类和泛性鸟类的种子传播效率。
南方红豆杉 (Taxus chinensis var. mairei) 为国家一级保护植物,广泛分布于中国南方山区,野生种群具两种生活状态:散生在连续的森林中或与其他阔叶树伴生并集中分布在斑块生境中,且以第2种状态更普遍[17]。前期研究表明,南方红豆杉果实被红色肉质假种皮所包裹,依赖于鹎科和鸦科的鸟类传播种子以实现更新;但花期不育,幼苗竞争力弱、种子捕食压力和种子萌发条件限制等因素导致南方红豆杉野生种群更新不佳[17, 18, 19, 20, 21]。然而,当原本的连续生境变成斑块生境后,生境斑块化如何影响专性鸟类和泛性鸟类的种子传播就对于濒危植物的续存至关重要,仍需进一步研究。为此,以分布于福建梅花山保护区红豆杉生态园南方红豆杉种群为对象,研究斑块生境中食果鸟类对南方红豆杉种子的取食和传播,并从传播距离和传播生境等两方面评估专性鸟类和泛性鸟类的传播效率,以此分析鸟类如何影响南方红豆杉的种群更新。试图回答以下科学问题:(1)斑块生境中南方红豆杉种群能吸引多少种专性鸟类和泛性鸟类取食并传播其种子?(2)专性鸟类和泛性鸟类的传播距离、传播生境有何差异及其对植物种群更新的影响?
1 材料与方法 1.1 研究地概况红豆杉生态园是福建省著名的国家森林公园,它位于福建梅花山自然保护区南部的崇头自然村。海拔895—1 218 m,生态园总面积15 hm2,属明显的中山地貌。南方红豆杉种群集中分布在阔叶林源斑块中,约有500株。年龄500a以上的南方红豆杉古树约200株,最大年龄约900a。源斑块内其他优势乔木还有福建柏(Fokienia hodginsii)、深山含笑(Michelia maudiae)和红楠(Machilus thunbergii)等。竹林斑块、竹阔边缘生境、农田斑块围绕着源斑块,形成明显的斑块生境(图 1)。

|
| 图 1 红豆杉生态园生境斑块的空间分布 Fig.1 The spatial distribution of patchy habitat in yew ecological garden |
野外调查于2011年和2012年南方红豆杉果期(10月底—12月中,果期持续时间45—50 d)进行。在30棵结实母树中选择其中5棵作为目标树,观察鸟类对目标树种子的取食。目标树的选择基于两点:一是所选目标树均为冠盖度大、结果量大的母树,能吸引最多的鸟取食其种子;二是5棵母树集中分布在一起。
每天6:30—17:00对目标树进行取食鸟类调查。采用焦点扫描取样法记录取食目标树果实或种子的鸟类种类、取食频次(观察期所记录每种鸟类的取食总次数)、每次取食量(颗)及取食时间、食物处理方式(整吞或啄食)等信息。如果是一群鸟同时取食,则选择视角最好的1只鸟进行观察。由于每粒果实内包含1粒种子,对于整吞果实的鸟种,可以认为其种子的吞食量等于种子搬运量[22]。调查以母树上果实变红开始,直至树上的红色果实完全消失为止。
1.3 专性鸟类和泛性鸟类的取食后行为当鸟类取食完南方红豆杉果实后,以红嘴蓝鹊(Urocissa erythrorhyncha)(体长: 65—68 cm;体重: 150—210 g;泛性鸟类)和黑短脚鹎(Hypsipetes leucocephalus)(体长: 24—25 cm;体重: 25—28 g;专性鸟类)两种典型的传播鸟类为观察对象,对鸟类取食后移动行为进行跟踪,以探明专性鸟类和泛性鸟类传播距离和传播生境的差异。具体操作如下:当一种鸟类取食完离开母树时,采用焦点扫描取样法记录红嘴蓝鹊、黑短脚鹎取食后移动行为[23]。每隔 15 s记录鸟类停歇的生境斑块、停歇树和停歇位置,确定位置后采用Newcon LR7×40型激光测距仪测量其与红豆杉母树的距离,以此定义为鸟类的潜在传播距离[24]。记录直至鸟类消失在视野内为止或者鸟类混入群体中无法区分。2011年累积观察时间600 h,共观察红嘴蓝鹊65次,黑短脚鹎60次,2012年累计观察时间550 h,共观察红嘴蓝鹊30次,黑短脚鹎65次。
1.4 专性鸟类和泛性鸟类的传播效率一些研究表明,植物1年生幼苗的数量与空间分布取决于传播动物的空间行为,因此本研究中以鸟类移动距离与幼苗更新距离、鸟类偏好微生境与幼苗更新生境的空间一致性程度来评价专性鸟类和泛性鸟类的传播效率;当二者空间一致性程度较高时,则认为鸟类的传播效率较高[24]。于2011年冬季,在竹林斑块和源斑块中设置长3 km,宽10 m的样线。行走样线时,统计1年生幼苗的数量(高度≤10 cm)[17, 25],发现幼苗后,测量幼苗与邻近母树的距离。此外,依据研究区内主要优势乔木,将微生境分为四种类型:(1)毛竹(Phyllostachys heterocycla)生境;(2)南方红豆杉同种成树生境;(3)甜楮(Castanopsis eyrei)及其他阔叶树生境;(4)空旷地生境。当鸟类停歇在微生境中时,记录该微生境位置,类型及幼苗数量,以此比较专性鸟类和泛性鸟类传播生境的差异。
1.5 数据分析采用独立样本T检验分析两年间食果鸟类取食频次的差异。并将整吞的鸟类作为南方红豆杉的潜在传播鸟类构建南方红豆杉种子传播系统,参考Bascompte等[26]互惠网络构建方法评价南方红豆杉对传播鸟类的依赖程度(传播鸟类取食频次占所有食果鸟类访问频次的比例)。
比较专性鸟类和泛性鸟类传播平均距离的差异。并采用Ivlev资源选择函数(Ei)计算鸟类对微生境的选择利用[27]。
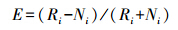
式中,Ri表示资源利用率,Ni表示资源利用率;若Ei=0,表示动物对资源 i 无选择性,用“0”表示;若Ei<0表示动物回避资源 i,用“-”表示;若Ei>0表示动物偏好选择资源 i,用“+”表示。
为分析专性鸟类和泛性鸟类传播效率的差异,分别分析鸟类传播距离和幼苗更新距离的空间一致性、鸟类传播生境与幼苗更新生境的空间一致性。具体操作如下:
根据前期统计各距离范围的幼苗相对数量,将距母树距离共分为0—5,5—15,15—25,25—35和>35 m等5个等级并计算一年生幼苗数量的相对比例,以此分析幼苗的更新距离;同时统计各等级中鸟类的停歇频次,利用Pearson卡方检验和似然比卡方检验评价以比较鸟类传播距离与幼苗更新距离间的一致性程度,P值越小时,表示二者一致性程度越低。
根据各生境斑块和微生境中幼苗的数量,计算相对比例。与此同时,统计各生境中鸟类的停歇频次。利用Pearson卡方检验和似然比卡方检验评价鸟类停歇地和幼苗分布的空间一致性。P值越小时,表示二者一致性程度越低[28]。
2 研究结果 2.1 鸟类-南方红豆杉的种子传播系统野外共观察到22种鸟取食南方红豆杉的果实(表 1),其中9种鸟啄食假种皮,而另外13种鸟类则整吞果实。共记录取食频次667次,不同年间鸟类对南方红豆杉取食频次差异不显著(t=0.523,P>0.05),但取食南方红豆杉种子的鸟类在种类上呈现出一定的年间变化:2011年,黑短脚鹎、红嘴蓝鹊和灰喉山椒鸟(Pericrocotus solaris)是南方红豆杉的主要取食鸟类,三者的取食频次达到该年总取食频次的91.6%,取食强度由高到低依次为黑短脚鹎、红嘴蓝鹊和灰喉山椒鸟;2012年,栗背短脚鹎(Hemixos castanonotus)、黑短脚鹎和红耳鹎(Pycnonotus jocosus)的取食频次较高,三者的取食频次达到该年总取食频次的79.9%,由高到低依次为黑短脚鹎、栗背短脚鹎和红耳鹎(表 1)。
斑块生境中,南方红豆杉共与13种鸟类传播者构成种子传播系统,其中7种鸟类为专性鸟类,6种鸟类为泛性鸟类。专性鸟类黑短脚鹎为南方红豆杉的最主要传播鸟类。除黑短脚鹎外,其他传播鸟类呈现出一定的年间变化。2012年,泛性鸟类红嘴蓝鹊的作用被栗背短脚鹎、红耳鹎和灰树鹊(Dendrocitta formosae)等鸟类所替代(表 1)。
2.2 专性鸟类和泛性鸟类的取食后行为取食后鸟类传播者取食后行为呈现出一定的差异,从而导致种子的传播距离和传播生境出现差异。与泛性鸟类红嘴蓝鹊相比,专性鸟类黑短脚鹎飞行的平均距离较短((16.3±11.0)m,Mean±SD,N=125),为南方红豆杉提供相对较近的传播距离,它们取食后偏好在阔叶林斑块中活动(阔叶林利用频次0.90 vs 竹林利用频次0.10,n = 100),喜栖息在同种成树(E = 0.23),甜楮及其他阔叶树(E = 0.19)等栖树上。泛性鸟类红嘴蓝鹊取食后常在斑块间移动,它们取食后常将大量的种子搬运至竹林斑块中(竹林利用频次0.84 vs阔叶林利用频次0.16,n = 100),常停歇在同种成树(E = 0.12)和毛竹(E = 0.33)上,提供相对较远的传播距离((24.9±20.0)m,Mean±SD,n = 95)(图 2)。
| 鸟种 Bird species | 取食频次Feeding frequency | 平均取食量/颗 Mean feeding amount | 平均取食时间/s Average feeding time | 取食方式 Feeding pattern | |
| 2011 | 2012 | ||||
| S,整吞Swallow; P,啄食Peak; *:泛性鸟类Generalist[29] | |||||
| 传播者Dispersers | |||||
| 黑短脚鹎 HypsipetesleucocephalusHL | 233 | 253 | 16.0±14.6 | 16.2±13.6 | S |
| 红嘴蓝鹊 UrocissaerythrorhynchaUE* | 22 | 8 | 16.3±9.1 | 18.4±16.4 | S |
| 鹊鸲 CopsychussaularisCS* | 7 | 5 | 8.0±3.5 | 11.4±5.5 | S |
| 灰树鹊 DendrocittaformosaeDF* | 6 | 15 | 19.9±11.5 | 23.8±11.9 | S |
| 栗背短脚鹎 Hemixos castanonotusHC | 3 | 22 | 9.6±9.9 | 20.6±12.0 | S |
| 大嘴乌鸦 CorvusmacrorhynchosCM* | 2 | - | 11.0 | 20.0 | S |
| 绿翅短脚鹎 Hypsopetes mcclellandiiHM | 1 | 2 | 12.0±4.0 | 15.0±5.0 | S |
| 领雀嘴鹎 SpizixossemitorquesSS* | 1 | 6 | 11.3±7.9 | 27.7±10.1 | S |
| 小黑领噪鹛 GarrulaxmonilegerGM | 1 | - | 6.0 | 10.0 | S |
| 灰翅噪鹛 Garrulax cineraceusGC | 1 | - | 2.0 | 6.0 | S |
| 红耳鹎 Pycnonotus jocosusPJ* | - | 19 | 16.2±14.2 | 33.3±25.8 | S |
| 黑领噪鹛 GarrulaxpectoralisGP | - | 5 | 14.4±6.9 | 22.0±21.4 | S |
| 红头咬鹃 Harpactes erythrocephalusHE | - | 1 | 3.0 | 5.0 | S |
| 取食者foragers | |||||
| 灰喉山椒鸟 Pericrocotus solarisPS | 19 | 6 | 16.2±14.2 | 33.0±25.8 | P |
| 赤红山椒鸟 Pericrocotus flammeusPF | 2 | 5 | 14.8±15.6 | 18.8±10.0 | P |
| 灰眶雀鹛 Alcippe morrisonia huetiAM | 1 | 3 | 22.3±9.3 | 32.7±23.7 | P |
| 白腹凤鹛 Yuhina zantholeuca.YZ | - | 13 | 4.1±2.2 | 25.6±16.9 | P |
| 北红尾鸲 PhoenicurusauroreusPA* | - | 4 | 7.3±2.2 | 12.0±5.4 | P |
| 红喉姬鹟 Ficedula( parva) albicillaFA | - | 1 | 5.0 | 10.0 | P |
| 大山雀 ParusmajorPM* | - | 5 | 4.0±1.9 | 17.4±9.1 | P |
| 黄腹山雀 PeriparusvenustulusPV | - | 2 | 7.5±0.5 | 22.5±2.5 | P |
| 丝光椋鸟 SturnussericeusST* | - | 1 | 10.0 | 25.0 | P |
| 总计Total | 299 | 368 | |||
幼苗更新显示出Janzen-Connell格局,大量幼苗聚集在距母树15 m处。在幼苗更新聚集最多的区域,黑短脚鹎移动停歇比例高,黑短脚鹎移动距离对幼苗更新距离的空间一致性程度(χ2=5.844,P =0.211)要大于红嘴蓝鹊(χ2=27.630,P <0.001)(图 3)。
幼苗集中分布在竹林斑块(N=233株),而源斑块较少(N=30颗)。4种微生境中均发现更新幼苗,其中毛竹生境内幼苗数量最多(图 4)。在幼苗聚集的生境斑块和微生境中,红嘴蓝鹊停歇较多,且它们选择的生境与幼苗更新生境一致性程度高(χ2=4.582,P =0.205),而黑短脚鹎生境与幼苗生境一致性低(χ2=26.371,P <0.001)。
3 讨论 3.1 斑块生境中鸟类-南方红豆杉的种子传播系统斑块生境中植物的种子传播系统特点影响着植物种群续存。传播者种类多是斑块生境中南方红豆 杉种子传播系统的主要特点之一。在红豆杉生态园,南方红豆杉依赖于7种专性鸟类,6种泛性鸟类传播种子(表 1)。红豆杉园植物种群所依赖的传播者种类高于浙江天目山、安徽仙寓山种群[19, 20, 21]。传播者种类的增加往往对植物种群更新更为有利,因为当种子传播系统中拥有较多种类的传播鸟类时,动物与植物之间的种子传播互惠关系更趋于稳定[22]。依赖更多的泛性鸟类传播也是斑块生境中南方红豆杉种子传播系统的重要特点。除黑短脚鹎对南方红豆杉的取食和传播占绝对优势,而其他种子传播者发生了明显的年间变化,泛性鸟类的传播作用在第2年有所增强(表 1)。这对于南方红豆杉种子传播系统的稳定往往是有利的。因为泛性鸟类抗干扰能力强,动植物种子传播系统能应对环境干扰的能力更强[30, 31],促使植物形成稳定的种子传播系统为斑块生境中植物种群的续存提供了可能[13, 22, 32]。

|
| 图 2 红嘴蓝鹊和黑短脚鹎的取食后行为比较 Fig.2 Post-foraging behavior comparison between H. leucocephalus and U. erythrorhyncha SDD: 短的传播距离Short dispersal distance; LDD: 长的传播距离Long dispersal distance |

|
| 图 3 红嘴蓝鹊和黑短脚鹎取食后移动距离与红豆杉幼苗更新距离的关系 Fig.3 Relationship between post-foraging movement distance of H. leucocephalus,U. erythrorhyncha and recruitment distance of yew seedling |
食果鸟类组合呈现出一定的年间变化,在取食南方红豆杉果实的主要鸟类中,仅黑短脚鹎同时出现在不同年份且取食量明显大于其他食果鸟类(表 1),这说明斑块生境中南方红豆杉与黑短脚鹎呈现了一种较为依赖的种子传播关系。这是濒危植物在斑块生境中续存的关键。除黑短脚鹎外,其他食果鸟类发生一定的年间变化。在2011年,红嘴蓝鹊和 灰喉山椒鸟为南方红豆杉种子的主要取食者,而在2012年,这两种鸟类的取食呈现出明显下降,栗背短脚鹎和红耳鹎代替了它们的作用(表 1)。表明第2年中传播者的取食比例更大,从而提高了鸟类对南方红豆杉种子的传播回报。因为当传播者的取食频次大于取食者时,更多的种子能被鸟类传播[33]。

|
| 图 4 不同生境中幼苗更新现状及红嘴蓝鹊和黑短脚鹎的生境利用 Fig.4 Recruitment pattern of seedling in different habitats and habitat use by H. leucocephalus,U. erythrorhyncha |
在斑块生境中,南方红豆杉以红嘴蓝鹊、鹊鸲、灰树鹊等泛性森林鸟类和黑短脚鹎、栗背短脚鹎等专性森林鸟类为主要的鸟类传播者(表 1)。南方红豆杉果实与两类传播鸟类形成种子传播关系,这亦是南方红豆杉在斑块生境中仍有更新幼苗出现的主要原因[18, 19, 20]。相关研究亦显示,虽然生境斑块化常导致传播动物多样性的下降,但并不意味着种子传播就一定会下降。一些以不同适应能力的鸟类为主要传播媒介的肉质果植物仍能获得较高的传播效率[22, 24, 34]。一方面,专性鸟类黑短脚鹎在斑块内搬运种子,为种子提供了短距离传播;而另一方面,泛性鸟类将种子搬离源斑块,为种子提供较长的传播距离(图 2,图 3),促进植物占据新的生境斑块[10]。这种传播距离上的互补往往对植物更新更有利[9]。
3.3 斑块生境中专性鸟类和泛性鸟类对南方红豆杉种子的传播效率鸟类是否能将种子搬运至植物的更新生境决定了斑块生境中植物种群续存的成败[35, 36, 37]。在红豆杉生态园中,大量幼苗聚集在竹林、毛竹冠幅下(图 4),这可能受到种子萌发条件的影响。南方红豆杉种子需两冬一夏才能萌发且萌发对微生境要求较高,需潮湿但排水条件好的土壤[17, 18, 38],而林下灌木密度过高又不利于幼苗的成活,因为红豆杉幼苗在与其他植物的竞争中常处于劣势[17]。在红豆杉生态园,农田等人为干扰斑块由于高强度的管护,地面几乎没有草本植被,土壤湿度太小,不利于种子萌发;红豆杉源斑块成树较多,Janzen-connell效应较强[39, 40],阔叶树下草本密度过高。因此,在红豆杉园中仅有竹林和毛竹才是南方红豆杉适宜的更新生境。那么,动物是否能将种子排放至竹林斑块、毛竹冠幅下就显得格外重要,但排放在哪儿往往取决于动物生境选择的决策[41, 42]。
在红豆杉园,专性鸟类和泛性鸟类取食后生境选择存在一定差异,导致种子排放生境不同。取食后,泛性鸟类红嘴蓝鹊偏好停歇在竹林斑块中,而专性鸟类黑短脚鹎则偏好在源斑块内活动(图 2,图 4),表明不同适应能力的鸟类对南方红豆杉种群更新的贡献不同,泛性鸟类更能将种子传播至更新斑块中。微生境的安全性,食物可利用性是影响鸟类生境利用的主要因子[41]。较高郁闭度的庇护所是鸟类取食红豆杉后的首要停歇点,因为红豆杉成树不仅为鸟类提供较好的隐蔽所,其果实也成为鸟类的选择食物,故鸟类重复利用红豆杉所在的微生境(图 2)。这种重复利用虽给幼苗带来较大的密度制约死亡率,但也为幼苗提供符合种群生长的适宜生境,极少的幼苗等待着建成的机会成为幼树。除偏好选择南方红豆杉同种成树外,两种鸟类对异种成树的利用显示出明显的差异。红嘴蓝鹊取食后更偏好选择毛竹栖息,而黑短脚鹎则少量利用甜楮等其他阔叶树(图 2)。异种成树下往往会给植物更新提供较好的空间。毛竹生境是幼苗萌发的适宜生境(图 2,图 4)。这暗示红嘴蓝鹊更能将种子沉积在适宜的更新生境中。由此可见,在南方红豆杉与鸟类传播者的种子传播系统中,泛性鸟类红嘴蓝鹊取食后的利用生境与幼苗更新生境空间一致性更高,更能促进斑块生境中南方红豆杉种群续存。因为鸟类对同种成树的重复利用,会促使少量幼苗获得建成幼树的潜在机会,而对庇护植物的利用则会促进植物更新[43, 44]。同时,野外观察也发现黑短脚鹎也会停歇在竹林中的一些阔叶树种上(图 2,图 4),造成种子沉积在竹林中,可推测它对南方红豆杉种群更新的贡献更多体现在于鸟类对更新生境的随机利用。
| [1] | Millennium Ecosystem Assessment (MEA). Chapter 11: biodiversity regulation of ecosystem services // Hassan R, Scholes R, Ash N. Ecosystems and Human Well-being: Current State and Trends, Volume 1, Findings of the Condition and Trends Working Group. Washington, DC: Island Press, 2005: 297-329. |
| [2] | Tylianakis J M, Didham R K, Bascompte J, Wardle D A. Global change and species interactions in terrestrial ecosystems. Ecology Letters, 2008, 11(12): 1351-1363. |
| [3] | Markl J S, Schleuning M, Forget P M, Jordano P, Lambert J, Traveset A, Wright J, Bhning-Gaese K. Meta-analysis of the effects of human disturbance on seed dispersal by animals. Conservation Biology, 2012, 26(6): 1072-1081. |
| [4] | McConkey K R, Prasad S, Corlett R T, Campos-Arceiz A, Brodie J F, Rogers H, Santamaria L. Seed dispersal in changing landscapes. Biological Conservation, 2012, 146(1): 1-13. |
| [5] | Franzén M, Nilsson S G. Both population size and patch quality affect local extinctions and colonizations. Proceedings of the Royal Society B: Biological Sciences, 2010, 277(1678): 79-85. |
| [6] | Snow B, Snow D. Birds and Berries: A Study of An Ecological Interaction. London: T &A D Limited, 1988. |
| [7] | Levey D J, Bolker B M, Tewksbury J J, Sargent S, Haddad N M. Effects of landscape corridors on seed dispersal by birds. Science, 2005, 309(5731): 146-148. |
| [8] | Jordano P, Forget P M, Lambert J E, Bhning-Gaese K, Traveset A, Wright S J. Frugivores and seed dispersal: mechanisms and consequences for biodiversity of a key ecological interaction. Biology Letters, 2011, 7(3): 321-323. |
| [9] | Nathan R, Getz W M, Revilla E, Holyoak M, Kadmon R, Smouse P E. A movement ecology paradigm for unifying organismal movement research. Proceedings of the National Academy of Sciences of the United States of America, 2008, 105(49): 19052-19059. |
| [10] | Spiegel O, Nathan R. Incorporating dispersal distance into the disperser effectiveness framework: frugivorous birds provide complementary dispersal to plants in a patchy environment. Ecology Letters, 2007, 10(8): 718-728. |
| [11] | Gómez J M. Spatial patterns in long-distance dispersal of Quercus ilex acorns by jays in a heterogeneous landscape. Ecography, 2003, 26(5): 573-584. |
| [12] | Lenz J, Fiedler W, Caprano T, Friedrichs W, Gaese B H, Wikelski M, Bhning-Gaese K. Seed-dispersal distributions by trumpeter hornbills in fragmented landscapes. Proceedings of the Royal Society of London B: Biological Sciences, 2011, 278(1716): 2257-2264. |
| [13] | Neuschulz E L, Botzat A, Farwig N. Effects of forest modification on bird community composition and seed removal in a heterogeneous landscape in South Africa. Oikos, 2011, 120(9): 1371-1379. |
| [14] | Carlo T A, García D, Martínez D, Geditsch J M, Morales J M. Where do seeds go when they go far? Integrating distance and directionality of avian seed dispersal in heterogeneous landscapes. Ecology, 2013, 94(2): 301-307. |
| [15] | Schupp E W, Jordano P, Gómez J M. Seed dispersal effectiveness revisited: a conceptual review. New Phytologist, 2010, 188(2): 333-353. |
| [16] | García D, Zamora R, Amico G C. Birds as suppliers of seed dispersal in temperate ecosystems: conservation guidelines from real-world landscapes. Conservation Biology, 2010, 24(4): 1070-1079. |
| [17] | Gao Z W. Research on Taxus chinensis var. mairei. Beijing: China Forestry Publishing House, 2006 |
| [18] | Lu C H, Zhu Q Q, Deng Q S. Effect of frugivorous birds on the establishment of a naturally regenerating population of Chinese yew in ex situ conservation. Integrative Zoology, 2008, 3(3): 186-193. |
| [19] | Shi G L, Bai B, Lu C H. Seed rain and seed bank of Chinese yew (Taxus chinensis var. mairei) population in Tianmu Mountain. Acta Ecologica Sinica, 2010, 30(5): 276-279. |
| [20] | Deng Q S, Zhu Q Q, Lu C H. Natural regeneration of Taxus chinensis var. mairei and its seed dispersl by frugivorous birds. Chinese Journal of Ecology, 2008, 27(5): 712-717. |
| [21] | Deng Q S, Chen S J, Lu C H. Foraging and transporting of Chinese yew Seed by Frugivorous Birds. Scientia Silvae Sinicae, 2010, 46(2): 157-161. |
| [22] | Farwig N, Bhning-Gaese K, Bleher B. Enhanced seed dispersal of Prunus africana in fragmented and disturbed forests? Oecologia, 2006, 147(2): 238-252. |
| [23] | Altmann J. Observational study of behavior: sampling methods. Behavior, 1974, 49(3/4): 227-263. |
| [24] | Breitbach N, Laube I, Steffan-Dewenter I, Bhning-Gaese K. Bird diversity and seed dispersal along a human land-use gradient: high seed removal in structurally simple farmland. Oecologia, 2010, 162(4): 965-976. |
| [25] | Swamy V, Terborgh J, Dexter K G, Best B D, Alvarez P, Coenejo F. Are all seeds equal? Spatially explicit comparisons of seed fall and sapling recruitment in a tropical forest. Ecology Letters, 2010, 14(2): 195-201. |
| [26] | Bascompte J, Jordano P, Olesen J M. Asymmetric coevolutionary networks facilitate biodiversity maintenance. Science, 2006, 312(5772): 431-433. |
| [27] | Manly B F J, McDonald L L, Tomas D L, McDonald T L, Erickson W P. Resource Selection by Animals: Statistical Design and Analysis for Field Studies. London: Kluwer Academic Press, 2002. |
| [28] | Fowler J, Cohen L, Jarvis P. Practical Statistics for Field Biology. 2nd ed. West Sussex: Open University Press, 1998. |
| [29] | MacKinnon J R, Phillipps K, He F Q. A Field Guide to the Birds of China. Oxford: Oxford University Press, 2000. |
| [30] | Moore J E, Swihart R K. Importance of fragmentation-tolerant species as seed dispersers in disturbed landscapes. Oecologia, 2007, 151(4): 663-674. |
| [31] | Carlo T A, Yang S A. Network models of frugivory and seed dispersal: Challenges and opportunities. Acta Oecologica, 2011, 27(6): 619-624. |
| [32] | Aslan C E. Implications of newly-formed seed-dispersal mutualisms between birds and introduced plants in northern California, USA. Biological Invasions, 2011, 13(12): 2829-2845. |
| [33] | Sawat S, Chen J, Wang B. Comparison of two plant-bird mutualistic systems in Asian tropical rainforests. Chinese Journal of Applied Ecology, 2011, 22(6): 1579-1584. |
| [34] | Graham C, Martinez-Leyva J E, Cruz-Paredes L. Use of fruiting trees by birds in continuous forest and riparian forest remnants in Los Tuxtlas, Veracruz, Mexico. Biotropica, 2002, 34(4): 489-597. |
| [35] | Howe H F, Smallwood J. Ecology of seed dispersal. Annual Review of Ecology and Systematics, 1982, 13(1): 201-228. |
| [36] | Zhao J, Chen J, Ma S B. Seed predation and dispersal of Ardisia solanacea in habitats with different degree of disturbance. Biodiversity Science, 2008, 16(1): 34-43. |
| [37] | Wei M S, Chen Z H, Ren H, Zou F S, Yin Z Y. Seed dispersal of the pioneer shrub Rhodomyrtus tomentosa by frugivorous birds and ants. Biodiversity Science, 2004, 12(5): 494-500. |
| [38] | Yue H J, Tong C, Zhu J M, Huang J F. Seed rain and soil seed bank of endangered Taxus chinensis var. mairei in Fujian, China. Acta Ecologica Sinica, 2010, 30(6): 4389-4400. |
| [39] | Janzen D H. Herbivores and the number of tree species in tropical forests. The American Naturalist, 1970, 104(940): 501-528. |
| [40] | Connell J H. On the role of natural enemies in preventing competitive exclusion in some marine animals and in rain forest trees // denBoer P J, Gradwell G R, eds. Dynamics of Populations. Wageningen: Centre for Agricultural Publishing and Documentation, 1971: 298-313. |
| [41] | Cody M L. Habitats Selection in Birds. London: Academic Press, 1985. |
| [42] | Barraquand F, Benhamou S. Animal movements in heterogeneous landscapes: identifying profitable places and homogeneous movement bouts. Ecology, 2008, 89(12): 3336-3348. |
| [43] | Alvarez-Loayza P, Terborgh J. Fates of seedling carpets in an Amazonian floodplain forests: intra-cohort competition or attack by enemies? Journal of Ecology, 2011, 99(4): 1045-1054. |
| [44] | Li N, Bai B, Lu C H. Recruitment limitation of plant population-from seed production to sapling establishment. Acta Ecologica Sinica, 2011, 31(21): 6624-6632. |
| [17] | 高兆蔚. 中国南方红豆杉研究. 北京: 中国林业出版社, 2006. |
| [20] | 邓青珊, 朱琼琼, 鲁长虎. 南方红豆杉的天然更新格局及食果鸟类对其种子的传播. 生态学杂志, 2008, 27(5): 712-717. |
| [21] | 邓青珊, 陈思静, 鲁长虎. 鸟类对南方红豆杉种子的取食与搬运. 林业科学, 2010, 46(2): 157-161. |
| [36] | 赵瑾, 陈进, 马绍宾. 不同扰动生境中动物对酸苔菜种子的捕食和散布. 生物多样性, 2008, 16(1): 34-43. |
| [37] | 韦明思, 陈章和, 任海, 邹发生, 殷祚云. 鸟类和蚂蚁对桃金娘种子传播的初步研究. 生物多样性, 2004, 12(5): 494-500. |
| [38] | 岳红娟, 仝川, 朱锦懋, 黄佳芳. 濒危植物南方红豆杉种子雨和土壤种子库特征. 生态学报, 2010, 30(6): 4389-4400. |
| [44] | 李宁, 白冰, 鲁长虎. 植物种群更新限制——从种子生产到幼树建成. 生态学报, 2011, 31(21): 6624-6632. |