 2014, Vol. 34
2014, Vol. 34文章信息
- 李洁, 张远东, 顾峰雪, 黄玫, 郭瑞, 郝卫平, 夏旭
- LI Jie, ZHANG Yuandong, GU Fengxue, HUANG Mei, GUO Rui, HAO Weiping, XIA Xu
- 中国东北地区近50年净生态系统生产力的时空动态
- Temporospatial variations in net ecosystem productivity in Northeast China since 1961
- 生态学报, 2014, 34(6): 1490-1502
- Acta Ecologica Sinica, 2014, 34(6): 1490-1502
- http://dx.doi.org/10.5846/stxb201305291221
-
文章历史
- 收稿日期:2013-5-29
- 修订日期:2013-11-14






2. 中国林业科学研究院森林生态环境与保护研究所, 国家林业局森林生态环境重点实验室, 北京 100091;
3. 中国科学院地理科学与资源研究所生态系统观测与模拟重点实验室, 北京 100101
2. Key Laboratory of Forest Ecology and Environment, State Forestry Administration, Institute of Forest Ecology, Environment and Protection, Chinese Academy of Forestry, Beijing 100091, China;
3. Key Laboratory of Ecosystem Network Observation and Modeling, Institute of Geographic Sciences and Natural Resources Research, CAS, Beijing 100101, China
陆地生态系统在全球碳循环中具有重要作用[1],在一定程度上具有减缓大气CO2浓度增加和气候变化的作用[2]。由于京都议定书已将陆地生态系统碳吸收纳入到温室气体减排的框架之中[3],因此对陆地碳汇时空变化的研究对京都议定书的执行具有重要意义[4, 5]。气候变化是影响陆地生态系统碳循环的因素之一[6, 7, 8]。在气候变暖条件下,中高纬度地区生态系统NPP通常表现为增加,土壤呼吸作用增强[9],这两个过程都是由温度升高所导致,对陆地碳汇却具有相反的作用。因此,综合分析生态系统碳循环过程对气候变化的响应,对于量化和理解陆地碳汇的区域分布和年际变化具有重要意义[5]。
作为净初级生产力NPP和异养呼吸HR的差值,净生态系统生产力(NEP)直接定性定量地描述陆地生态系统碳源/汇的性质和能力。在不考虑各种自然和人为扰动的情况下,NEP可近似看作是陆地生态系统与大气系统之间的净碳交换量[10],最近十几年来,越来越受到科学家的关注[11, 12, 13]。以往研究表明,从20世纪80年代到90年代,陆地生态系统的碳蓄积量明显增加了[2, 14, 15],并且北半球中高纬度陆地生态系统是其中一个巨大碳汇[2, 12, 16],但对陆地碳汇时空格局的变化及其与气候变化关系的方面仍存在很大不确定性[17, 18]。Cao等研究了气候变化对中国陆地生态系统碳蓄积的影响,在气候变化和CO2浓度增加的共同作用下,中国陆地生态系统总体上具有碳汇作用,并且碳源汇分布存在区域差异,中国大部分地区NEP接近于零[6]。Piao基于ORCHIDEE模型研究了气候变化对青藏地区草地生态系统NPP、NEP的时空变化的影响,结果表明降水对NEP增加的贡献为36%,是引起NEP年际变化的主要驱动因素,温度对该区净碳蓄积增加没有明显作用[13]。Lu等基于TEM模型在美国中西部地区的研究表明,美国中西部地区陆地生态系统NEP具有0.62TgC/a的增加趋势,年平均NEP与降水呈正相关,与温度的相关性不显著[8]。以上研究结果表明,气候变化对NEP的空间格局变化的影响存在区域差异,而且研究时段主要在1980年以后的20a,而陆地碳汇的变化是长期气候变化的结果[2, 5],因此有必要在更长时间尺度上对NEP的时空动态及其对气候变化的响应进行研究。
东北地区处于我国最高纬度地区,是中国最大的天然林区和重要的商品粮生产基地,也是世界三大黑土带之一[19, 20, 21]。东北地区是全球气候变化的敏感区域之一[22]。鉴于东北地区特殊的地理位置以及在全球变化中的重要作用,国内许多学者开展了对东北地区碳收支的研究[20, 23, 24, 25],但研究大多是对NPP和土壤呼吸这两个碳收支过程的单独研究,而且主要集中于森林和农田生态系统[20, 24, 25],不能反映整个东北地区的净碳收支状况。目前仅有的对东北地区NEP的研究,研究时间段短,并且没有对该地区NEP的时空动态变化以及与气候变化的关系进行深入探讨。王军邦等模拟了东北地区NPP和NEP的空间格局[26]。赵俊芳等人模拟了当前气候状况和不同气候情景对东北森林生态系统NPP和NEP的影响,认为NPP和NEP对温度升高的反应比对降水的反应更为敏感[27]。基于此,本研究应用高分辨率的气候数据库和生态系统过程模型CEVSA,对1961—2010年东北地区净生态系统生产力(NEP)的时空格局及变化趋势进行分析,并讨论气候变化与NEP的关系,阐明影响东北区域NEP变化的关键气候因子。
1 资料与方法 1.1 模型简介CEVSA建立于1998年,该模型是一个基于生理生态过程模拟植物-土壤-大气系统能量交换和水碳氮耦合循环的生物地球化学循环模型[28, 29, 30]。CEVSA模型包含3个子模型:植被子模型用来计算植被的NPP、分配和凋落物产量;生物物理子模型计算蒸散、土壤水分动态和气孔导度;生物地球化学子模型计算凋落物和土壤有机碳的分解和传输[28]。CEVSA模型已应用于区域和全球尺度,模拟分析了陆地生态系统碳循环对气候变化的响应[5, 28, 29, 30, 31, 32, 33, 34, 35, 36]。以下是对模型主要过程模拟方法的简要描述,关于模型更详细的结构可以参考相关文献[28, 37, 38]。
1.1.1 光合和气孔导度光合作用速率决定于叶肉组织光合酶对CO2的利用效率和CO2向叶肉组织的扩散速率,气孔导度的变化采用改进的Ball-Berry模型模拟。
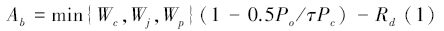
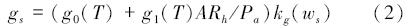
式中,Ab代表由酶系统活性决定的碳同化速率,gs为气孔传导度,Wc由Rubisco所决定的羧化速率,Wj是由电子传递速度决定的羧化速率,Wp决定于光合反应过程对磷酸丙糖(Triose phosphate)的利用效率。Po和Pc分别是叶肉组织中O2和CO2的分压。τ是Rubisco的特异因子,Rd为白昼非光合呼吸速率。go是在光补偿点下光合速率为零时的气孔传导度,g1是灵敏度参数,A为实际光合速率,T为绝对温度,kg(ws)为气孔导度对土壤含水量ws的反应函数。
1.1.2 氮吸收和自养呼吸植物的氮含量决定了植物的最大光合速率,氮吸收对温度的依赖性(NT)可表达为:
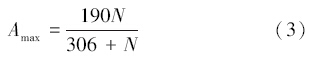

式中,Amax为最大光饱和的光合速率,N为某一层叶片的氮含量,NT则为植物的总氮吸收量,u1、u2、u3和kT(T)均为温度的响应函数,Tk为绝对温度。叶片的氮含量(N)还决定了叶片的维持呼吸(Rml)[39]:
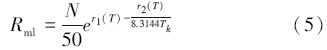
式中,r1(T)和r2(T)分别是温度的响应函数。
1.1.3 土壤碳氮动态CEVSA模型将土壤有机质分为8个碳库,每个碳库都具有特定的分解速率,并与碳库本身特性、温度、湿度、氮的有效性和土壤质地有关。氮矿化量(Na)和土壤异养呼吸(HR)可分别用下式表达:

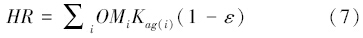
式中,i表示不同的碳库。OM是每个碳库的大小,K是潜在衰变率,CN是碳氮比,Kag(i)是实际各个土壤有机碳库的衰变率,ε为同化效率。
1.2 输入数据来源、参数获取和模型运行模型中所使用的气象数据来自于国家气象信息中心,包括1961—2010年全国756个气象台站的每旬平均气温,降水量,云量和相对湿度。使用ANUSPLIN4.1插值软件的样条函数插值法对气象数据进行内插,得到全国1961—2010年的0.1°气象栅格数据。大气CO2浓度资料来源于美国夏威夷Mauna Loa观测所(http://co2now.org/Current-CO2/CO2-Now/noaa-mauna-loa-co2-data.html)。土壤类型和质地资料取自1 ∶ 100万土壤类型图和第二次土壤普查数据,对其进行数字化并重采样到0.1°。植被数据来自于(European Commission,Joint Research Centre,2003. http://bioval.jrc.ec.europa.eu/products/glc2000/glc2000.php),共有22种土地覆被类型,在ArcGIS中进行插值重采样,匹配于气象数据的分辨率。
CEVSA模型包含43个参数和11个初始的状态变量。在CEVSA模型应用于区域和全球的模拟时,初始的状态变量,如植被碳、土壤碳、土壤碳氮比、初始的土壤含水量等来源于静态模型的输出。与土壤质地有关的参数包含田间持水量、饱和含水量、萎蔫系数、土壤分解的最佳持水量等参数,这些土壤水分参数由土壤颗粒组成确定,依据Raich等[40]、McGuie等[41]和Cao和Woodward[28]等文献中提供的数据进行估算。CEVSA最初包含13种土地覆盖类型,与植被类型相关的参数主要是根、茎、叶的周转时间、根、茎、叶的分配比例和比叶面积等,主要来源于已发表文献的估计。在本研究中使用的22种土地覆盖类型,如与最初模型中已有土地覆盖类型直接对应的,就使用原有参数,不直接对应的植被类型,本研究通过查阅文献和模型调试的方法获取相对应植被类型的相关参数。详细的参数列表和取值方法可以参考已发表的文章,如Cao和Woodward[28]、Zhang等[38] 、Gu等[42]。
本研究中,模型运行的时间分辨率为10d,空间分辨率为0.1°。首先应用1961—2010年的平均气候数据运行模型至生态系统平衡态,即各个状态变量如植被、土壤碳贮量以及土壤含水量等年际变化量小于0.1%,且NPP、凋落物产生量(LT)与HR相等,然后用1961—2010年每旬资料进行动态模拟,并反复运行模型以消除假定的生态系统状态变量的初始值(即平衡态假设)对模拟结果的影响。从最终模拟结果中提取中国东北地区的数据进行分析。
1.3 数据分析(1)时间序列变化分析:对年平均温度、降水、NEP序列做一元线性回归分析,线性方程的斜率代表温度、降水、NEP的变化趋势。(2)空间序列变化分析:逐个格点计算NEP一元线性回归的回归系数,得到NEP的空间变化趋势;(3)由于本研究中的NEP的计算结果,是通过温度与降水等因子在模型中运算得到的,NEP与温度和降水之间不具有独立性,但出于分析的需要,本研究不考虑这一关系,采用相关分析的方法,研究温度、降水对NEP的影响程度。NEP与温度降水的相关分析:用PEARSON相关系数法,计算NEP与年平均温度、降水量的相关系数,并进行显著性检验,当相关系数通过显著性水平(P<0.05)时,认为NEP与温度、降水有显著相关关系。
1.4 研究区域概况本研究中,东北地区(北纬 38°48′—53°33′N,东经 115°31′—135°05′E)主要包括黑龙江、吉林、辽宁以及内蒙古地区东北部,北部与俄罗斯接壤,东南部与朝鲜半岛相接,南部滨临中国渤海和黄海[43]。本研究中统计的总面积为11.73×105 km2,1961—2010东北地区年平均温度变化在2.49—6.02℃之间,从南到北随着纬度升高,年均温逐渐降低,年降水量变化在427—680mm,从东南向西北,降水量逐渐减少。冬季寒冷干燥,夏季炎热多雨,特殊的地理位置和气候特点,形成了东北地区独特的植被分布格局。从南向北随着热量的变化,出现暖温带、温带和寒温带,植被也相应形成了暖温带落叶阔叶林带,温带针阔混交林带和寒温带针叶林带;从东南向西北随着湿度的分异,出现湿润地区、半湿润地区和半干旱地区,植被则有森林、森林草原、典型草原植被[44]。
2 结果分析 2.1 模型验证CEVSA模型所应用的生物和生态学原理、计算方程和参数均取自于大量的实验室和野外试验观测,曾应用叶片和植株生理试验、样点初级生产力观测和遥感反演数据进行了验证,模型估算的NPP、叶面积指数及植被和土壤中的碳储量与实地调查和测定所得的结果有很好的一致性[28, 30]。但CEVSA模型自建立后一直缺乏在生态系统尺度上的验证。20世纪90年代后大型环境控制试验和涡度相关技术的应用提供了连续、长期的生态系统尺度碳通量观测数据[45, 46, 47]。它们为研究生态系统尺度的碳交换[48, 49]以及模型的检验测试[50]提供了大量有用的数据。为检验该CEVSA模型对生态系统碳通量季节动态的模拟效果,评价模型对生态系统碳循环过程机理定量描述的准确性,顾峰雪等[51]在2007年曾应用多个站点的涡度相关观测数据对模型的模拟结果进行了验证和评价,包括位于东北地区的长白山站,结果表明CEVSA模型能够较好地模拟长白山针阔混交林碳交换的季节和年际动态(图 1)。

|
| 图 1 长白山站NEP模拟值与通量观测值的比较NEP观测值与模拟值的季节动态;NEP观测值与模拟值的1 ∶ 1线 Fig.1 Comparison between modeled and measured monthly net ecosystem productivity (NEP) in Changbaishan station from 2003 to 2005 simulated and observed monthly NEP; correlation of simulated and observed monthly NEP |
另外,为了进一步验证模型在区域模拟时对碳收支的模拟效果,本研究应用东北地区81个实测站点地面NPP数据[52, 53, 54]对模型模拟的NPP进行了验证,这81个实测站点包含了落叶阔叶林、落叶松林、针阔混交林等东北地区的多种森林类型。原数据中以生物量表示的NPP,本文中取碳转化率0.45,得到以碳含量表示的实测NPP。81个实测站点NPP数据的平均值为491 gC · m-2 · a-1,根据样点坐标提取的1961—2010年模拟NPP年均值为479 gC · m-2 · a-1,平均相对误差为2.2%。NPP年均模拟值与对应样点NPP实测值间的相关系数为r=0.584 (P<0.001),NPP实测值与模拟值基本吻合(图 2)。同时,本研究将CEVSA模型模拟的NPP与已有研究进行了比较,结果表明,本研究模拟的东北地区平均的NPP为439 gC · m-2 · a-1,在以往其他研究结果的范围内(400.85—445 gC · m-2 · a-1)[23, 55, 56]。另外,本研究收集了东北地区17个实测土壤异养呼吸数据[57, 58, 59, 60, 61, 62, 63, 64, 65, 66],这17个实测数据包含了农田、森林、草地等多种植被类型,模拟值与对应经纬度土壤异养呼吸实测值间存在显著相关性,相关系数为r=0.880 (P<0.01)(图 2)。

|
| 图 2 NPP,HR 模拟值与地面实测值的比较 Fig.2 Comparison of modeled NPP and HR with the observed values in the Northeast |
近50年中,东北地区陆地生态系统是大气CO2的汇,但碳吸收的年际间变异很大。1961—2010年,东北地区年NEP总量最低值为-0.094PgC/a,最高值为0.117PgC/a,年平均为0.026PgC/a。年NEP单位平均值变化范围为-79.0—99.8 gC · m-2 · a-1,平均为22.5 gC · m-2 · a-1,变异系数为206%。研究时段内年NEP总量具有下降趋势,但线性变化趋势不显著(P=0.240)。在年代际总量上,20世纪80年代的碳汇强度最大(52.47TgC/a),占研究时段内总碳吸收量的40%。60年代和70年代的碳吸收处于较高水平(27.73TgC/a,26.11TgC/a),分别占研究时段总量的21%和20%。90年代和2000—2010年后碳吸收量急剧减少(13.02TgC/a和10.51TgC/a),分别占研究时段总量的10%和8%。研究时段内,仅有30%的年份表现为碳源,且碳源强度较弱,除20世纪90年代后期以及21世纪初期向大气释放了少量碳外,中国东北地区陆地生态系统在其他时期主要表现为碳汇,尤其是80年代(图 3)。

|
| 图 3 研究时段内东北地区净生态系统生产力NEP的年际变化及年代际变化 Fig.3 Interannual variations and decadal variations in NEP during 1961—2010 in the Northeast |
研究时段内,东北地区的年平均温度变化在2.49—6.02℃之间,温度升高明显(P<0.05),年降水量427.26—680.72mm,趋势倾向率为-5.4mm/10a,变化趋势不显著,降水的年际变化较大,变异系数为9.62(图 4)。NEP的年际变化与温度呈显著负相关(r=-0.343,P<0.05),与降水相关性极显著(r=0.859,P<0.01),同时东北地区NEP和年降水量的变化规律基本一致,即同期上升或达到最高值。因此,温度和降水共同作用导致东北地区NEP的年际变化,而年降水量的变化对NEP年际变化起主要作用。

|
| 图 4 1961—2010年东北地区年平均温度、降水量的年际变化及年代际变化 Fig.4 Interannual variations and decadal variations in mean annual temperature and mean annual precipitation during 1961—2010 in the Northeast |
东北地区NEP呈现出东部高,西部和中部低,北部高,南部低的空间分布特点。NEP正值代表碳汇,NEP负值代表碳源。东北平原的部分地区,内蒙古高原部分地区以及三江平原小部分地区为碳源区,碳源区仅占全区总面积的15.7%。碳汇值较高的地区分布在大、小兴安岭和长白山的森林分布区,约为45—60gC · m-2 · a-1,部分地区达到60—100 gC · m-2 · a-1。碳汇较低的地区主要分布在辽宁,三江平原,内蒙古与辽宁、吉林交界处的农田分布区,碳汇强度约为0—30gC · m-2 · a-1(图 5 a)。

|
| 图 5 1961—2010年东北地区NEP的空间分布 (a) 及其变化趋势 (b) Fig.5 Spatial distribution (a) and trend (b) of annual NEP during 1961—2010 in the Northeast |
从NEP的变化趋势来看,东北地区NEP的变化趋势与其空间分布有良好的一致性。NEP增加地区的面积占研究区域总面积的31.8%,而显著增加的面积仅占总面积29.3%,主要集中在三江平原地区,内蒙古高原等碳源区。NEP减少地区的面积占研究区域总面积的66.8%,其中显著减少的面积占研究区域总面积的55%,主要在长白山西麓,大、小兴安岭等碳汇区(图 5 b)。总体来看,碳源区向大气释放的碳量在减少,碳汇区从大气吸收的碳也在减少。
东北地区NEP与温度呈显著负相关(P<0.05)的面积占31.6%,主要分布在大兴安岭东麓,小兴安岭以及长白山地区(图 6)。NEP与降水呈显著正相关(P<0.05)的面积占研究区域总面积的91.5%,只有大、小兴安岭和长白山的部分地区与降水的相关性不显著(图 6)。因此,在空间上,东北地区NEP空间分布主要由降水决定,只有一小部分地区的NEP受温度控制。

|
| 图 6 1961—2010年东北地区NEP与温度、降水的相关关系 Fig.6 Correlation coefficient between annual NEP and mean annual temperature and mean annual precipitation during 1961—2010 in the Northeast |
1961—2010年,东北地区净生态系统生产力(NEP)单位平均值为22.5gC · m-2 · a-1,低于在西南高山地区NEP值的29.7gC · m-2 · a-1[36]。东北地区固定的总碳量为0.026PgC/a,占全国NEP总量的15%—37%[6, 67, 68]。在年代际变化量上,Cao在中国区域内的研究结果表明,与20世纪80年代相比,东北地区NEP总量在90年代的减少,抵消了南部地区NEP的增加,本研究中,80年代到90年代,东北地区NEP总量减少了39.45TgC/a,与Cao的研究结果一致[6],而且在2000年以后,东北地区NEP总量仍在减少,但变化量很小,为2.51TgC/a。
NEP的年际变化与气候所导致的净初级生产力(NPP)和异养呼吸(HR)的变化密切相关。研究时段内,东北地区增温明显,年平均NPP与温度不相关,HR与温度有极显著的正相关关系(r=0.505,P<0.01),温度每升高1℃,东北地区由土壤异养呼吸向大气释放的碳增加14.85Tg(图 7)。NPP与降水有极显著的正相关关系(r=0.942,P<0.01),降水量每增加100mm,东北地区的植被净初级生产力增加89.4Tg,而HR与降水的相关关系不显著(图 7)。这表明东北地区降水不足,是植被生长的限制因子,这与国志兴等基于EOS/MOS卫星遥感资料的研究,分析东北地区年均NPP与温度和降水量的相关性结论一致[23]。

|
| 图 7 东北地区NPP、HR与温度、降水的相关关系 Fig.7 Sensitivity to temperature and precipitation of NPP and HR in the Northeast |
NPP和HR共同决定了NEP的年际变化,年平均NEP与NPP和HR的相关系数分别为0.867(P<0.05)和-0.338(P<0.05)。这表明东北地区的陆地碳汇主要是由于降水量的变化影响植被生产力引起的,与之相比,温度对土壤呼吸作用对陆地碳汇的贡献相对较小,这与在东亚地区的研究存在差异[12],在东亚地区,温度和太阳辐射的年际变化是导致区域碳收支年际变化的主要原因,可能原因是东亚大部分地区处于湿润的季风气候区,降水量的变化和水分胁迫没有显著影响,NPP对降水的敏感性很低,而东北地区处于欧亚大陆的东缘,自东向西,气候上从湿润区、半湿润区过渡到半干旱区,降水是植被生长的限制因子。
东北地区气候变暖导致HR增加,而NPP随着降水量的年际变化呈波动趋势(图 8),两者的综合作用导致东北地区NEP的年际变化没有明显的线性变化趋势,而是与降水量的年际波动趋势相一致。20世纪60和70年代,东北地区碳吸收量分别占研究时段总量的21%和20%,80年代的碳吸收量显著增加,这与80年代东北地区降水量增加导致的NPP增加量大于温度升高造成土壤呼吸的增加量有关,最终导致NEP呈增长趋势(图 8)。进入90年代,尤其是在90年代后期,东北地区由于降水量下降NPP减少趋势明显,同期的土壤呼吸量却显著增加,碳吸收强度有所下降。21世纪的前10a,东北地区的温度继续升高,降水显著减少,碳吸收总量较90年代减少了2.51TgC/a,这一时期中的2000年和2007年,是研究时段内净生态系统生产力(NEP)最低的两年,降水量分别为465mm和411mm,低于降水量的多年平均值,因而NPP水平很低,而且由于温度的升高伴随着土壤呼吸的增加,导致这两年东北地区净碳释放量达-0.094PgC和-0.093PgC,约是1961—2010年年平均碳吸收的3.6倍,但这期间东北地区陆地生态系统仍起着碳汇作用。因此,近20年,升温伴随降水减少是东北区域碳汇能力减弱的重要原因。

|
| 图 8 1961—2010年东北地区NPP和HR的年际变化 Fig.8 Interannual variations in NPP and HR in the Northeast |
NEP的空间分布和变化趋势与温度和降水也有密切关系。东北平原、内蒙古高原以及三江平原的部分区域,是东北地区主要的碳源区,在过去50a,由于降水与净初级生产力呈正相关关系,降水的减少造成NPP减少,同时温度的降低使得土壤呼吸作用减弱,但降水引起的NPP减少量小于温度降低造成的土壤呼吸减少量,使得过去50a这些地区的NEP呈增加趋势,即碳源区向大气释放的碳量在减少。大、小兴安岭和长白山等主要碳汇区,大部分地区NEP与温度呈极显著负相关,与降水正相关,低温或多雨是这些地区形成碳汇的主要原因。研究时段内,碳汇区温度升高,降水减少,导致土壤呼吸增高,NPP减少,NEP呈减少趋势,即碳汇区从大气吸收的碳量在减少。
4 结论(1)过去50a东北区域NEP没有明显的线性变化趋势,NEP和年降水量的变化规律基本一致,即同期上升或达到最高值。东北地区的陆地碳汇主要是由于降水量的变化影响植被生产力引起的,与之相比,温度对土壤呼吸作用对陆地碳汇的贡献相对较小。
(2)NEP的空间分布呈现出东部高,西部和中部低,北部高,南部低的空间趋势,并随降水量和温度分布呈现区域异质性。降水是决定NEP空间分布的最主要因子。
(3)升温伴随降水增加导致1961—1990年NEP呈增加趋势,而其后升温伴随降水减少则是近20年东北区域碳汇能力减弱的重要原因。
| [1] | Canadell J G, Mooney H A, Baldocchi D D, Berry J A, Ehleringer J R, Field C B, Gower S T, Hollinger D Y, Hunt J E, Jackson R B, Running S W, Shaver G R, Steffen W, Trumbore S E, Valentini R, Bond B Y. Carbon metabolism of the terrestrial biosphere: A multitechnique approach for improved understanding. Ecosystems, 2000, 3(2):115-130. |
| [2] | Schimel D S, House I J, Hibbard K A, Bousquet P, Ciais P, Peylin P, Braswell B H, Apps M J, Baker D, Bondeau A, Canadell J, Churkina G, Cramer W, Denning A S, Field C B, Friedlingstein P, Goodale C, Heimann M, Houghton R A, Melillo J M, Moore Ⅲ B, Murdiyarso D, Noble I, Pacala S W, Prentice I C, Raupach M R, Rayner P J, Scholes R J, Steffen W L, Wirth C. Recent patterns and mechanisms of carbon exchange by terrestrial ecosystems. Nature, 2001, 414(6860):169-172. |
| [3] | IGBP Terrestrial Carbon Working Group. The terrestrial carbon cycle: implications for the Kyoto Protocol. Science, 1998, 280(5368):1393-1394. |
| [4] | Take K R, Scott N A, Parshotam A, Brown L, Wilde R H, Giltrap D J, Trustrum N A, Gomez B, Ross D J. A multi-scale analysis of a terrestrial carbon budget: Is New Zealand a source or sink of carbon?. Agriculture, Ecosystems & Environment, 2000, 82(1/3):229-246. |
| [5] | Cao M K, Prince S D, Tao B, Small J, Li K R. Regional pattern and interannual variations in global terrestrial carbon uptake in response to changes in climate and atmospheric CO2. Tellus B, 2005, 57(3):210-217. |
| [6] | Cao M K, Prince S D, Li K R, Tao B, Small J, Shao X M. Response of terrestrial carbon uptake to climate interannual variability in China. Global Change Biology, 2003, 9(4):536-546. |
| [7] | Mohamed M A A, Babiker I S, Chen Z M, Ikeda K, Ohta K, Kato K. The role of climate variability in the inter-annual variation of terrestrial net primary production (NPP). Science of the Total Environment, 2004, 332(1):123-137. |
| [8] | Lu X L,Zhuang Q L. Evaluating climate impacts on carbon balance of the terrestrial ecosystems in the Midwest of the United States with a process-based ecosystem model. Mitigation and Adaptation strategies Global Change, 2010, 15(5):467-487. |
| [9] | Xu X F, Tian H Q, Wan S Q. Climate warming impacts on carbon cycling in terrestrial ecosystems. Journal of Plant Ecology(Chinese Version), 2007, 31(2): 175-188. |
| [10] | Woodwell G M, Whitaker R H, Reiners W A, Likens G E, Delwich C C, Botkin D B. Biota and the world carbon budget. Science; (United States), 1978, 199(4325):141-146. |
| [11] | Tian H Q, Melillo J M, Kicklighter D W, McGuire A D, Helfrich Ⅲ J V K, Moore Ⅲ B, Vorosmarty C J. Effect of interannual climate variability on carbon storage in Amazonian ecosystems. Nature, 1998, 396(6712):664-667. |
| [12] | Ito A. The regional carbon budget of East Asia simulated with a terrestrial ecosystem model and validated using AsiaFlux data. Agricultural and Forest Meteorology, 2008, 148(5):738-747. |
| [13] | Piao S L, Tan K, Nan H J, Ciais P, Fang J Y, Wang T. Impacts of climate and CO2 changes on the vegetation growth and carbon balance of Qinghai-Tibetan grasslands over the past five decades. Global and Planetary Change, 2012, 98-99:73-80. |
| [14] | Gurney K R, Law R M, Denning A S, Rayner P J, Baker D, Bousquet P, Bruhwiler L, Chen Y H, Ciais P, Fan S M, Fung I Y, Gloor M, Heimann M, Higuchi K, John J, Maki T, Maksyutov S, Masarie K, Peylin P, Prather M, Pak B C, Randerson J, Sarmiento J, Taguchi S, Takahashi T, Yuen C W. Towards robust regional estimates of CO2 sources and sinks using atmospheric transport models. Nature, 2002, 415(6872):626-630. |
| [15] | Houghton R A. Why are the estimates of the terrestrial carbon balance so different?. Global Change Biology, 2003, 9(4):500-509. |
| [16] | Pacala S W, Hurtt G C, Baker D, Peylin P, Houghton R A, Birdsey R A, Heath L, Sundquist E T, Stallard R F, Ciais P, Moorcroft P, Caspersen J P, Shevliakova E, Moore B, Kohlmaier G, Holland E, Gloor M, Harmon M E, Fan S M, Sarmiento J L, Goodale C L, Schimel D, Field C B. Consistent land- and atmosphere-based U.S. carbon sink estimates. Science, 2001, 292(5525):2316-2320. |
| [17] | Tao B,Cao M K, Li K R, Gu F X, Ji J J, Huang M, Zhang L M. Spatial patterns of terrestrial net ecosystem productivity in China during 1981-2000. Science in China (Series D:Earth Sciences), 2007, 50(5):745-753. |
| [18] | Liu W G. Research of carbon and NPP changes of the land ecosystem in XinJing . Wulumuqi: Xinjiang University, 2007. |
| [19] | Yu L, Zhang B. The degradation situations of black soil in China and its prevention and counter measures. Journal of Arid Land Resources and Environment, 2004, 18(1):99-103. |
| [20] | Zhu F, LiuZ M, Wang Z M, Song K S. Temporal-spatial characteristics and factors influencing crop NPP across Northeastern China. Resources Science, 2010, 32(11):2079-2084. |
| [21] | Yu Y, Fan W Y, Li M Z. Forest carbon rates at different scales in Northeast China forest area. Chinese Journal of Applied Ecology, 2012, 23(2):341-346. |
| [22] | Wang S Q, Zhou C H, Liu J Y, Li K R, Yang X M. Stimulation analyses of terrestrial carbon cycle balance model in Northeast China. Acta Geographica Sinica, 2001, 56(4):390-400. |
| [23] | Guo Z X, Wang Z M, Zhang B, Liu D W, Yang G, Song K S, Li F. Analysis of temporal-spatial characteristics and factors influencing vegetation NPP in Northeast China from 2000 to 2006. Resources Science, 2008, 30(8):1226-1235. |
| [24] | Liu X,Guo Q X, Liu J W. Simulation of spatial pattern of soil respiration in forests in the eastern part of Northeast China. Journal of Northeast Forestry University. 2010, 28(12):58-64. |
| [25] | Liu X,Guo Q X, Liu J W. Sensitivity analysis for main factors influencing NPP of forests simulated by IBIS in the eastern area of Northeast China. Acta Ecologica Sinica, 2011, 31(7):1772-1782. |
| [26] | Wang J B. Chinese terrestrial net ecosystem productive model applied remote sensing data . Hangzhou: Zhejiang University,2004. |
| [27] | Zhao J F, Yan X D,Jia G S. Simulating the responses of forest net primary productivity and carbon budget to climate change in Northeast China. Acta Ecologica Sinica, 2008,28(1):92-102. |
| [28] | Cao M K, Woodward F I. Net primary and ecosystem production and carbon stocks of terrestrial ecosystems and their responses to climate change. Global Change Biology, 1998, 4(2):185-198. |
| [29] | Cao M K, Woodward F I. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature, 1998, 393(6682):249-252. |
| [30] | Woodward F I, Smith T M, Emanuel W R. A global land primary productivity andphytogeography model. Global biogeochemical cycles, 1995, 9(4):471-490. |
| [31] | Cao M K, Prince S D,Shugart H H. Increasing terrestrial carbon uptake from the 1980s to the 1990s with changes in climate and atmospheric CO2. Global Biogeochemical cycles, 2002, 16(4):1069. |
| [32] | Cao M K, Zhang Q F, Shugart H H. Dynamic responses of African ecosystem carbon cycling to climate change. Climate Research, 2001, 17(2):183-193. |
| [33] | Cao M K, Tao B, Li K R, Shao X M, Prience S D. Interannual variation in terrestrial ecosystem carbon fluxes in China from 1981-1998. Acta Botanica Sinica(Chinese Edition), 2003, 45(5):552-560. |
| [34] | Tao B. Modeling research on terrestrial net primary productivity & net ecosystem productivity in China . Beijing: Graduate University of Chinese Academy of Sciences, 2003. |
| [35] | Li K R, Wang S Q,Cao M K. Carbon storage of vegetation and soil in china. Science in China (Series D), 2003, 33(1):72-80. |
| [36] | Pang R,Gu F X, Zhang Y D, Hou Z H, Liu S R. Temporal-spatial variations of net ecosystem productivity in alpine area of southwestern China. Acta Ecologica Sinica, 2012, 32(24):7844-7856. |
| [37] | Gu F X, Cao M K, Wen X F, Liu Y F, Tao B. A comparison between simulated and measured CO2 and water flux in a subtropical coniferous forest. Science in China (Series D:Earth Sciences), 2006, 49( Suppl Ⅱ): 241-251. |
| [38] | Zhang L, Yu G R,Gu F X, He H L, Zhang L M, Han S J. Uncertainty analysis of modeled carbon fluxes for a broad-leaved Korean pine mixed forest using a process-based ecosystem model. Journal of Forest Research, 2012, 17(3): 268-282. |
| [39] | Harley P C, Thomas R B, Reynolds J F, Strain B R. Modelling photosynthesis of cotton grown in elevated CO2. Plant, Cell & Environment, 1992, 15(3): 271-282. |
| [40] | Raich J W, Rastetter E B, Melillo J M, Kicklighter D W, Steudler P A, Peterson B J, Grace A L, Moore Ⅲ B, Vorosmarty C J. Potential net primary productivity in south America: Application of a global model. Ecological Applications, 1991, 1(4): 399-429. |
| [41] | McGuire A D,Melillo J M, Joyce L A, Kicklighter D W, Grace A L, Moore Ⅲ B, Vorosmarty C J. Interactions between carbon and nitrogen dynamics in estimating net primary productivity for potential vegetation in North America. Global Biogeochemical Cycles, 1992, 6(2): 101-124. |
| [42] | Gu F X, Zhang Y D, Tao B, Wang Q F, Yu G R, Zhang L M, Li K R. Modeling the effects of nitrogen deposition on carbon budget in two temperate forests. Ecological Complexity, 2010, 7(2): 139-148. |
| [43] | Mao D H, Wang Z M,Luo L, Han J X. Spatio-temporal pattern of net primary productivity and its driven factors in Northeast China in 1982-2010. Scientia Geographica Sinica, 2012, 32(9):1106-1111. |
| [44] | Xu W D, He X Y, Chen W, Liu C F, Zhao G L, Zhou Y. Ecological division of vegetation in Northeast China. Chinese Journal of Ecology, 2008, 27(11):1853-1860. |
| [45] | Lloyd J, Grace J, Miranda A C, Meir P, Wong S C,Miranda H S, Wright I R, Gash J H C, McIntyre J. A simple calibrated model of Amazon rainforest productivity based on leaf biochemical properties. Plant Cell and Environment, 1995, 18(10): 1129-1145. |
| [46] | Baldocchi D D. Assessing the eddy covariance technique for evaluating carbon dioxide exchange rates of ecosystems: past, present and future. Global Change Biology, 2003, 9(4): 479-492. |
| [47] | Yu G R, Zhang L M, Sun X M, Fu Y L, Li Z G. Advances in carbon flux observation and research in Asia. Science in China(Series D: Earth Sciences), 2005, 48( SupplⅠ): 1-16. |
| [48] | Baldocchi D, Falge E, Gu L H, Olson R, Hollinger D, Running S, Anthoni P. FLUXNET: A new tool to study the temporal and spatial variability of ecosystem-scale carbon dioxide, water vapor, and energy flux densities. Bulletin of the American Meteorological Society, 2001, 82(11): 2415-2434. |
| [49] | Falge E, Baldocchi D, Tenhunen J, Aubinet M, Bakwin P, Berbigier P, Bernhofer C. Seasonality of ecosystem respiration and gross primary production as derived from FLUXNET measurements. Agricultural and Forest Meteorology, 2002, 113(1): 53-74. |
| [50] | Baldocchi D D, Wilson K B. Modeling CO2 and water vapor exchange of a temperate broadleaved forest across hourly to decadal time scales. Ecological Modelling, 2001, 142(1): 155-184. |
| [51] | Gu F X, Cao M K, Yu G R, Tao B, Wen X F, Liu Y F, Zhang L M. Modeling carbon exchange in different forest ecosystems by CEVSA model: Comparison with eddy covariance measurements. Advances in Earth Science, 2007, 22(3):223-234. |
| [52] | Luo T X. Patterns of net primary productivity for Chinese major forest types and their mathematical models . Beijing: Chinese Academy of Sciences,1996. |
| [53] | Zhao M. Zhou G S. A new methodology for estimating forest NPP based on forest inventory data A case study of Chinese pine forest. Journal of Forestry Research, 2004, 15(2):93-100. |
| [54] | Wang Y H, Zhou G S, Jiang Y L, Yang Z Y. Estimating biomass and NPP ofLarix forests using forest inventory data (FID). Acta Phytoecologica Sinica. 2001, 25(4): 420-425. |
| [55] | Zhang F, Zhou G S. Spatial-temporal variation in net primary productivity along Northeast China Transect (NECT) from 1982 to 1999. Journal of Plant Ecology (Chinese Version). 2008, 32(4):798-809. |
| [56] | Zhao G S, Wang J B, Fan W Y, Ying T Y. Vegetation net primary productivity in Northeast China in 2000-2008: Simulation and seasonal change. Chinese Journal of Applied Ecology, 2011, 22(3):621-630. |
| [57] | Zhu J J, Yan Q L, Fan A N. Soil carbon flux for secondary forest ecosystems in the northeast mountains, China//Proceeding of the Fifth Youth Ecologists Conference. Guangdong: Guangdong Province Association for Science and Technology, 2008: 112-120. |
| [58] | Jiang C S,Hao Q J, Song C C, Hu B Q. Effects of marsh reclamation on soil respiration in the Sanjiang Plain. Acta Ecologica Sinica. 2010, 30(17): 4539-4548. |
| [59] | Liu Y. Soil carbon efflux in the typical forest ecosystems of Changbai Mountains . Beijing: Graduate University of Chinese Academy of Sciences, 2007. |
| [60] | Liu Q J, Zhang G C,Xu Q Q, Wang Y D, Wang H M. Soil respiration of two dominant tundra communities under controlled temperatures in Changbai Mountain, Northeast China. Acta Ecologica Sinica, 2011,31: 137-44. |
| [61] | Han G X, Zhou G S,Xu Z Z. Seasonal dynamics of soil respiration and carbon budget of maize (Zea mays L.) farmland ecosystem. Chinese Journal of Eco-Agriculture, 2009, 17(5): 874-879. |
| [62] | Wang X, Zhou G S, Jiang Y L, Li F. Comparison of soil respiration in broad-leaved Korean pine forest and reclaimed cropland in Changbai Mountains, China. Journal of Plant Ecology. 2006, 30(6):887-893. |
| [63] | Yang J Y, Wang C K. Soil carbon storage and flux of temperate forest ecosystems in northeastern China. Acta Ecologica Sinica, 2005, 25(11): 2875-2881. |
| [64] | Lu B, Wang S H, Mao Z J, Sun T,Jia G M, Jin S B, Sun P F, Cheng C X. Soil respiration characteristics of four primary Korean pine communities in growing season at Xiaoxing'an Mountain, China. Acta Ecologica Sinica, 2010, 30(15):4065-4074. |
| [65] | Qi Y C, Dong Y S, Manfred D, Geng Y B, Liu L X, Liu X R. Comparison of CO2 effluxes and their driving factors between two temperate steppes in Inner Mongolia, China. Advances in Atmospheric Sciences, 2006, 23(5): 726-736. |
| [66] | Qi Y C, Dong Y S, Liu L X, Liu X R, Peng Q, Xiao S S, He Y T. Spatial-temporal variation in soil respiration and its controlling factors in three steppes of Stipa L. in Inner Mongolia, China. Science China (Series D:Earth Sciences), 2010, 53(5): 683-693. |
| [67] | Mu Q Z, Zhao M S, Running S W, Liu M L,Tian H Q. Contribution of increasing CO2 and climate change to the carbon cycle in China's ecosystems. Journal of Geophysical Research: Biogeosciences (2005-2012), 2008, 113(G01018):1-15. |
| [68] | Fang J Y,Guo Z D, Piao S L, Chen A P. Terrestrial vegetation carbon sinks in China from 1981 to 2000. Science in China (Series D: Earth Sciences), 2007, 37(6): 804-812. |
| [9] | 徐小锋, 田汉勤, 万师强. 气候变化对生态系统碳循环的影响. 植物生态学报, 2007, 31(2): 175-188. |
| [17] | 陶波, 曹明奎, 李克让, 顾峰雪, 季劲钧, 黄玫, 张雷明. 1981—2000年中国陆地净生态系统生产力空间格局及其变化. 中国科学(D辑), 2006, 36 (12): 1131-1139. |
| [18] | 刘卫国. 新疆陆地生态系统净初级生产力和碳时空变化研究[D]. 乌鲁木齐: 新疆大学, 2007. |
| [19] | 于磊, 张柏. 中国黑土退化现状与防治对策. 干旱区资源与环境, 2004, 18(1): 99-103. |
| [20] | 朱锋, 刘志明, 王宗明, 宋开山. 东北地区农田净初级生产力时空特征及其影响因素分析. 资源科学, 2010, 32(11): 2079-2084. |
| [21] | 于颖, 范文义, 李明泽. 东北林区不同尺度森林的含碳率. 应用生态学报, 2012, 23(2): 341-346. |
| [22] | 王绍强, 周成虎, 刘纪远, 李克让, 杨晓梅. 东北地区陆地碳循环平衡模拟分析. 地理学报, 2001, 56(4): 390-400. |
| [23] | 国志兴, 王宗明, 张柏, 刘殿伟, 杨桄, 宋开山, 李方. 2000年— 2006年东北地区植被NPP的时空特征及影响因素分析. 资源科学, 2008, 30(8): 1226-1235. |
| [24] | 刘曦, 国庆喜, 刘经伟. 东北东部森林土壤呼吸空间分布格局的模拟. 东北林业大学学报. 2010, 28(12):58-64. |
| [25] | 刘曦, 国庆喜, 刘经伟. IBIS 模拟东北东部森林NPP 主要影响因子的敏感性. 生态学报, 2011, 31(7): 1772-1782. |
| [26] | 王军邦. 中国陆地净生态系统生产力遥感模型研究[D]. 杭州: 浙江大学, 2004. |
| [27] | 赵俊芳, 延晓冬, 贾根锁. 东北森林净第一性生产力与碳收支对气候变化的响应. 生态学报, 2008, 28(1): 92-102. |
| [34] | 陶波. 中国陆地生态系统净初级生产力和净生态系统生产力模拟研究[D]. 北京:中国科学院研究生院, 2003. |
| [35] | 李克让, 王绍强, 曹明奎. 中国植被和土壤碳储量. 中国科学(D辑), 2003, 33(1): 72-80. |
| [36] | 庞瑞, 顾峰雪, 张远东, 侯振宏, 刘世荣. 西南高山地区净生态系统生产力时空动态. 生态学报, 2012, 32(24): 7844-7856. |
| [37] | 顾峰雪, 曹明奎, 温学发, 刘允芬, 陶波. 亚热带针叶林水碳通量的模拟及其与观测的对比研究. 中国科学(D辑), 2006, 36(增刊I): 224-233. |
| [43] | 毛德华, 王宗明, 罗玲, 韩佶兴.1982—2010年中国东北地区植被NPP时空格局及驱动因子分析. 地理科学, 2012, 32(9): 1106-1111. |
| [44] | 徐文铎, 何兴元, 陈玮, 刘常富, 赵桂玲, 周园. 中国东北植被生态区划. 生态学杂志, 2008, 27(11): 1853-1860. |
| [51] | 顾峰雪, 曹明奎, 于贵瑞, 陶波, 温学发, 刘允芬, 张雷明. 典型森林生态系统碳交换的机理模拟及其与观测的比较研究. 地理科学进展, 2007, 22(3): 223-234. |
| [52] | 罗天祥. 中国主要森林类型生物生产力格局及其数学模型[D]. 北京: 中国科学院地理科学与资源研究所, 1996. |
| [54] | 王玉辉, 周广胜, 蒋延玲, 杨正宇. 基于森林资源清查资料的落叶松林生物量和净生长量估算模式.植物生态学报, 2001, 25(4): 420-425. |
| [55] | 张峰, 周广胜. 中国东北样带植被净初级生产力时空动态遥感模拟. 植物生态学报, 2008, 32(4): 798-809. |
| [56] | 赵国帅, 王军邦, 范文义, 应天玉. 2000—2008年中国东北地区植被净初级生产力的模拟及季节变化. 应用生态学报, 2011, 22(3): 621-630. |
| [57] | 朱教君, 闫巧玲, 范阿南. 东北山区次生林生态系统土壤碳通量研究//第五届中国青年生态学工作者学术研讨会论文集. 广东:广东省科学技术协会,2008: 112-120. |
| [58] | 江长胜, 郝庆菊, 宋长春, 胡必琴. 垦殖对沼泽湿地土壤呼吸速率的影响. 生态学报, 2010, 30(17): 4539-4548. |
| [59] | 刘颖. 长白山典型森林生态系统土壤碳通量研究[D]. 北京:中国科学院研究生院, 2007. |
| [61] | 韩广轩, 周广胜, 许振柱. 玉米农田生态系统土壤呼吸作用季节动态与碳收支初步估算. 中国生态农业学报, 2009, 17(5): 874-879. |
| [62] | 王旭, 周广胜, 蒋延玲, 李峰. 长白山红松针阔混交林与开垦农田土壤呼吸作用比较. 植物生态学报, 2006, 30(6): 887-893. |
| [63] | 杨金艳, 王传宽. 东北东部森林生态系统土壤碳贮量和碳通量. 生态学报, 2005, 25(11): 2875-2881. |
| [64] | 陆斌, 王淑华, 毛子军, 孙涛, 贾桂梅, 靳世波, 孙鹏飞, 程春香.小兴安岭4种原始红松林群落类型生长季土壤呼吸特征. 生态学报, 2010, 30(15): 4065-4074. |
| [66] | 齐玉春, 董云社, 刘立新, 刘杏认, 彭琴, 肖胜生, 何亚婷. 内蒙古锡林河流域主要针茅属草地土壤呼吸变化及其主导因子. 中国科学(D辑), 2010, 40(3): 341-351. |
| [68] | 方精云, 郭兆迪, 朴世龙, 陈安平. 1981—2000 年中国陆地植被碳汇的估算. 中国科学(D辑), 2007,37(6): 804-812. |