 2014, Vol.34
2014, Vol.34文章信息
- 卢训令, 梁国付, 汤茜, 丁圣彦, 李乾玺, 张晓青
- LU Xunling, LIANG Guofu, TANG Qian, DING Shengyan, LI Qianxi, ZHANG Xiaoqing
- 黄河下游平原农业景观中非农生境植物多样性
- Plant species of the non-agricultural habitats in the lower reaches of the Yellow River plain agro-landscape
- 生态学报, 2014, 34(4): 789-797
- Acta Ecologica Sinica, 2014, 34(4): 789-797
- http://dx.doi.org/10.5846/stxb201209251353
-
文章历史
- 收稿日期:2012-9-25
- 修订日期:2013-2-5






2. 河南大学生命科学学院, 开封 475004;
3. 河南大学生态科学与技术研究所, 开封 475004
2. School of Life Sciences, Henan University, Kaifeng 475004, China;
3. Institute of Ecological Science and Technology, Henan University, Kaifeng 475004, China
农业景观是地球陆地表面最主要的景观类型之一,是人类生存生活所需资料的最主要来源地。农业生态系统功能和服务的稳定与持续是人类生存和福祉的基础。生物多样性是影响生态系统功能和服务发挥的最关键因素之一[1, 2]。农业景观中的生物多样性对系统内生态过程、功能、服务和持续具有极其重要的作用,农业系统的可持续发展离不开生物多样性已成为共识[3]。研究表明农业景观中非农生境对生物多样性保护、景观结构和农业产量等具有重要作用[4, 5, 6]。而农业景观中的生物多样性的维持依赖于生境多样性和异质性的存在和维持[7, 8],通过增加农业景观多样性和异质性以提高生物多样性已成为欧盟和美国农业环境保护的重要内容[9, 10],研究者还探讨了一些被认为能提高生物多样性的土地利用和农业管理策略[11, 12, 13]。随着人口的激增,粮食需求、农业用地的增加和生物多样性保护间的权衡已成为全球性的挑战[13]。如何在农业景观中既维持较高的生产力,又进行有效的生物多样性保护就显得尤为重要。为此,国内外研究人员对农业景观中不同土地利用方式、不同景观格局或不同耕作模式与管理方式下,农业生态系统生态过程和功能的变化开展了大量研究[14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24],研究尺度涵盖了区域(景观)、局地和田块等不同水平[4, 5, 9, 22, 23, 24, 25]。研究的生物类群涵盖了动物、植物、土壤微生物等,但以动物类群为主[14, 15, 16, 17, 18, 19, 20]。受“杂草”称呼和对农产品产量负效应的影响,对农业景观中植物多样性保护及其格局、过程和生态效应的研究相对较少[21, 22, 23],而农业景观中的植物群落又是动物的食源地、栖息地、越冬地和庇护所等,对其他生物具有重要的生态意义。如何进行农业景观的构建,才能达到农业生产力的持续与生物多样性保护和生态系统功能维持间的平衡?这一问题尚未能很好的解决。
黄河下游平原是我国黄淮海平原的主体和最重要的粮食主产区之一,耕作历史最为悠久,人类活动对农业景观中生物多样性和生态系统稳定性产生了重要影响,但目前关于区内农业生态系统生物多样性的研究还较少[26, 27],区内农业景观中物种多样性格局如何变化?不同生境中物种多样性变化特征是怎样的?区域内不同生境对植物多样性保护的贡献度如何?这些问题均尚待解决。而对这些的问题的研究又是探讨农业景观生态过程、功能和服务健康可持续的基础。为此,本研究通过对黄河中下游平原农业景观格局、群落结构和物种多样性的调查分析,探讨区内主要非农生境中的物种多样性与群落结构特征,可为未来区域可持续农业景观构建和生态系统服务功能维持的研究提供理论支持,并为区域城乡景观规划和设计实践提供技术支撑。
1 研究区概况

|
| 图 1 研究区地理位置及样地布局示意图 Fig. 1 Schematic diagram of the study area′s location and plots distribution |
研究区位于河南省新乡市封丘县,地处黄河故道,是典型的黄河下游冲积平原区,全国的商品粮基地县之一。地理位置为北纬34°53′—35°14′,东经114°14′—114°45′之间,总面积约1225.6 km2(图 1)。区内地势由西南向东北倾斜,成土母质为第四纪全新世以来的黄河冲积物,土壤类型以潮土为主,属暖温带大陆性季风气候,多年平均降雨量约615 mm,年均温13.9 ℃,地带性植被类型为落叶阔叶林,但目前已几乎消失殆尽,取而代之的是大量的农田和少量的人工植被。
2 研究方法 2.1 样地设置在研究区卫星影像和土地利用现状图的基础上,设置4 km×4 km网格,在每个网格中选取1 km×1 km的网格为调查区域,在每个调查区中选取主要的非农生境(主要包括人工林地、树篱、田间道路和沟渠等)进行调查,每个生境类别5个样方,共布设26个样点,350个调查样方(图 1)。采用典型样地法对各样方的草本植被进行群落调查,主要记录指标为物种、多优度、群集度、物候期等,同时记录各样地的地理坐标。因人工林地和树篱虽然存在乔木层,但其物种绝大多数均为单一的杨树,且在整个研究区内各生境中几乎均不存在灌木层,故整个研究和数据分析中所用的资料均为草本层数据。野外调查时间为2011年5月10日—5月25日,并于2012年4月26日—5月5日进行了复查。
2.2 指数计算(1)景观格局分析 以2007年土地利用现状图和2009年遥感数据为基础,使用ArcGis10.0对区内的景观格局特征进行分析,考虑到行政区域的完整性,景观格局分析数据以整个封丘县行政区界为限。
(2)物种丰富度 以样地中出现的物种数目S表示。
(3)物种多样性指数 选取Shannon-Wiener指数为指标,计算公式为:
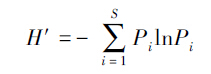
式中,H′为Shannon-Wiener指数;S为样方内物种数目;Pi为物种i在群落中的重要性,为物种i的多优度与总多优度的比值。
(4)均匀度指数
本文选用基于Shannon-Wiener多样性指数的均匀度指数进行研究:

(5)Beta多样性 选取Whittaker Beta多样性指数来反映生境中群落组成变化的幅度和分化程度。计算公式为:
β=γ/α-1
式中,γ为某取样生境中的总物种数,α为某生境类型所有取样点物种数的平均值。
(6)相似性系数
分别选取Jaccard相似性系数、Czekanowski相似性系数(又称Srensen系数)和Gleason群落相似性系数进行计算,其中Gleason系数由Jaccard相似性系数演变而来,选用的计算指标为重要值,具体的计算公式分别如下:
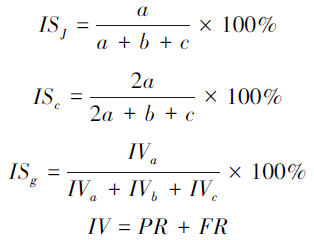
式中,ISJ、ISc和ISg分别为Jaccard、Czekanowski和Gleason相似性系数,a为两个群落共有的物种数,b和c分别为两个群落独有的物种数,IV为物种的重要值,PR为物种的相对显著度,由物种的多优度计算获得,FR为物种的相对频度[28],所有物种的重要值总和为2。
3 结果分析 3.1 景观格局特征分析景观类型分析表明,封丘县为典型的农业区,景观总体为农业景观,农田(包括旱田、水田和设施农业等)是最主要的景观类型,总面积达81367.9 hm2,约占全县总面积的66.4%;居民点景观类型广泛散落在区域内,大小不一,约占总面积的11.6%;林地景观多呈斑块状散布在农田中和居民点周边,以人工种植的杨树林为主,间有极少量的混交林和次生林,总面积约8672.6 hm2;水域景观主要为黄河河道和散布在区内的一些河流与引黄灌渠,面积较小;黄河滩区位于黄河主河道和大堤之间,总面积约9795.3 hm2,现多已被垦为耕地,间有一些砖窑厂(图 2)。

|
| 图 2 封丘县景观类型分布图 Fig. 2 Landscape classification map of Fengqiu County |
为了解农业景观中主要非农植物的生物地理分布态势,对群落调查数据进行统计分析发现:统计中的127种植物隶属于菊科(Compositae)、 禾本科(Gramineae)、十字花科(Cruciferae)、豆科(Leguminosae)、 藜科(Chenopodiaceae)等38科,107属,其中蕨类植物1种。所有植物中分布最多的为菊科(16属23种),其次是禾本科(17属17种),十字花科(7属8种),豆科(7属7种),这4科物种占了总种数的43.31%。
分别参照李锡文和吴征镒对中国种子植物科和属分布区类型的划分[29, 30],对区内的植物区系分布进行研究(图 3),结果表明:区域内植物科的分布主要是以世界分布(15科,70种)和泛热带分布(14科,30种)为主,二者占了总科数的76.3%和总物种数的79.4%,其次是北温带分布(7科,21种),只有极少量的旧世界温带分布(1科,4种)和全温带分布(1科,1种),缺少特有分布科;属的地理成分复杂,区内具有中国15个种子植物属分布区类型中的13个,仅缺少中亚分布与中国特有分布型,以温带分布(33属39种)、世界分布(23属30种)和泛热带分布(15属19种)为主,以上3个分布型占了全区属种数的69.84%和66.36%。区系分析表明,区内的植物物种组成以耐干扰的广布种为主,缺少特有种。

|
| 图 3 植物区系分布特征 Fig. 3 Distributional characteristics of the flora |
对区内不同生境类型物种多样性进行分析(图 4)。结果表明:物种丰富度在林地和树篱中最高(分别为108和106种),其次是田间道路,共86种,沟渠最少,仅59种;Shannon多样性指数呈现林地最高(3.71),其次是树篱(3.57),沟渠(3.24),田间道路最低(2.43),且林地和树篱中的物种多样性显著高于田间道路;各生境中的物种均匀度以沟渠中最高,显著高于树篱和田间道路,林地的物种均匀度也较高;水渠、林地和树篱均具有较高的草本层盖度,田间道路则显著降低,同时不同林地群落间盖度差异较大。
通过计算各生境中的β多样性指数研究了不同生境内部群落组成的变化幅度和分化程度(图 4),结果显示:田间道路各样地中群落组成的变化幅度最大,不同样点间物种组成差异性最高,其次是树篱和林地,沟渠最低。这种变化趋势表明区域内人类活动和生境内生态因子的异质性可能起着重要作用。
3.4 不同生境间群落异质性分析不同生境中群落优势种重要值分析显示,群落主要优势种组成有一定的差异(表 1),结合物种数目——累积重要值变化曲线发现,田间道路中的优势种优势度最明显,沟渠、树篱和林地中顺次降低(图 5),推测在农业景观中,随着人类活动干扰的增强,群落中优势种的地位会更加突出,优势种对整个群落的控制作用显著。群落相似性指数表明,各指数的变化趋势基本一致,只是数值高低稍有不同而已(图 5),三者均显示:林地、树篱和田间道路之间的相似性系数明显较高,远高于沟渠与之的相似性。 表明在黄河下游平原农业景观中,除沟渠外,各生境中物种组成具有极高的相似性,但群落优势种和盖度等级具有较大的差异,尤其田间道路与林地、树篱间差异显著(图 4),推测农业景观中虽然各生境间的物种组成差异不大,但其生态功能和服务具有较大的差距。

|
| 图 4 不同生境植物多样性特征比较 采用单因子方差分析进行显著性检验,α=0.05 Fig. 4 Comparison of plant diversity of different habitats |
| 树篱 Hedge | 林地Forest | 田间道路Road | 沟渠 Ditch | |||||
| 物种名
Species | 重要值
IV | 物种名
Species | 重要值
IV | 物种名
Species | 重要值
IV | 物种名
Species | 重要值
IV | |
| 1 | 狗牙根
Cynodon dactylon | 0.12 | 夏至草
Lagopsis supina | 0.15 | 狗牙根
C. dactylon | 0.29 | 狗牙根
C. dactylon | 0.17 |
| 2 | 刺儿菜
Cirsium setosum | 0.10 | 葎草
H. scandens | 0.09 | 播娘蒿
Descurainia sophia | 0.09 | 稗子
Echinochloa crusgali | 0.13 |
| 3 | 雀麦
Bromus japonicus | 0.09 | 枸杞
L. chinense | 0.08 | 打碗花
Calystegia hederacea | 0.09 | 芦苇
Phragmites australis | 0.12 |
| 4 | 葎草
Humulus scandens | 0.09 | 刺儿菜
C. setosum | 0.08 | 萹蓄
Polygonum aviculare | 0.09 | 风花菜
Rorippa globosa | 0.08 |
| 5 | 枸杞
Lycium chinense | 0.08 | 婆婆纳
Veronica didyma | 0.06 | 婆婆纳
V. didyma | 0.08 | 委陵菜
Potentilla chinensis | 0.07 |
| 合计Total | 0.48 | 0.46 | 0.65 | 0.57 | ||||
| 比例Ratio/% | 24.03 | 22.88 | 32.54 | 28.36 | ||||

|
| 图 5 不同生境间群落异质性比较 Fig. 5 Comparison of community heterogeneity of different habitats |
本研究发现,黄河下游平原区农业景观不同生境中的物种丰富度、多样性、均匀度和群落盖度均有一定的差异,以林地和树篱中的物种多样性和丰富度最高,林地在物种多样性保护方面的优势并不甚明显;沟渠是区域内一个较为特殊的生境类型,群落组成分化程度很低,群落相似性系数分析也表明沟渠景观中的植物群落是一个独立的类别,其群落组成与其他群落差异较大;林地、树篱、田间道路等生境间的群落相似性极高,建议在更大尺度上(如景观或区域尺度)进行物种多样性研究时,可将区内农业景观中的这些生境视为同一个类别看待,不需再单独分类,但群落盖度和优势种分析表明,田间道路与林地、树篱间存在显著差异,据此这些生境间极高的相似性仅是组成物种名录的相似性,而生态功能的差异则是明显的。农业景观中人类活动和干扰强度对群落组成意义重大,随着人类干扰强度的增加,群落物种组成的变异和分化程度迅速增加,种群更多的呈群集和斑块状分布,如在田间道路上,多形成以狗牙根(C. dactylon)或萹蓄(P. aviculare)等耐践踏,强阳性物种为优势种的单优势群落,群落物种重要值排序也显示田间道路等干扰强烈的生境中优势种群更明显。区系分析的结果表明,在科的分布型中,优势分布型明显,属的地理成分十分复杂,优势属分布型明显。总之区内植物物种以广布种为主,缺少特有种,优势科属分布型明显。在区域内进行的生物多样性保护重在对其生态服务功能的维持和提高,而非濒危、特有物种的保护。
植物群落组成、结构、动态和物种多样性对区域生态系统结构、功能和服务的发挥具有至关重要的作用,非农生境中的植物群落在农业景观中具有重要的生态服务功能,如为动物提供食物、栖息地。Robinson等的研究表明,战后随着农业景观的变化,树篱的清除,鸟类因非繁殖季节的食物供应下降而降低[31];同时对农药、化肥、降水等具有过滤、沉降和缓冲效应,从而起到水源保障的作用,也是濒危物种和天敌的避难所[8]。为了提高农业地区的生物多样性,欧盟提出了农业环境计划(Agri-Environment Schemes),尽管后来有些研究人员认为该计划对区域生物多样性提升的作用很有限,尤其是鸟类和哺乳动物[32, 33],但非农生境的增加对植物多样性具有强烈的促进作用[34, 35]。国内的一些研究也表明农业生态系统中的非农生境在动物多样性保护方面具有重要作用[36],但关于植物群落方面的研究还较少[37, 38],对我国东部区域典型平原农业景观植物群落结构、动态过程依然不明了。目前农业生态系统的可持续发展已经日益引起大家的关注,农业生态系统的健康与持续离不开系统内非农生境及其上的生物群落,在未来的区域农业景观的规划和管理过程中,如何布局和管理这些非农生境就显得尤为重要。
黄河下游平原是我国最主要的粮食主产区和核心区,该区域的农业生态系统的健康可持续发展是当前我们必须面对和亟待解决的科学和社会难题,在未来的农业景观规划和布局中必须充分重视非农生境的重要性,尽管研究人员对于各种不同景观斑块在农业景观中的作用已有众多的探讨[4, 5, 6],已经证实这些非农生境对于区域内的生态系统结构和功能具有重要作用,但在农业生产、粮食安全和生态功能间进行权衡和取舍中,如何进行农业景观的优化,何种景观结构和构型才更合理、更高效、更稳定、更持续?关于这个问题目前尚无定论[24]。景观生态学、岛屿生物地理学中关于大面积林地斑块和廊道(树篱)的研究表明它们对生物多样性保护和生态系统功能具有重要作用[39, 40],但在农业景观中,在当前农业生态系统的第一要务依然是粮食生产和粮食安全保障的情况下,如何进行更合理的景观要素配置,以提高农业景观生态系统的稳定性和持续性,这种景观格局的优化和合理的空间布局仍是目前景观生态学研究中的一个重要论题[41]。结合已有的研究成果认为,在农业景观中,在保证农业用地的同时,构建更多的树篱,提高景观连通性,能够有效的提高农业景观中的生物多样性[42, 43]。本研究也显示树篱具有相当高的物种丰富度、多样性和群落盖度。另外需改造目前已有的物种单一的人工林地,因为人工纯林是不存在乔木和灌木多样性的,在生态功能方面的弱势和危害是显而易见的[44],而更应因地发展混交林或“近自然林”,使之成为自然、半自然斑块,以提供更高的、更稳定的生态功能和服务[45]。同时,沟渠作为一种特殊生境,一方面是区域内一些湿生物种的庇护所,同时沟渠上的植物群落能提供净化水质、改善小气候等的作用,在规划沟渠的过程中应减少水泥护坡的使用。但如何在农业景观中进行更有效的生物多样性保护和生态系统功能维持与提高依然需要进一步研究,需要政府决策人员、农民和研究人员的共同努力。
| [1] | Mace G M, Norris K, Fitter A H. Biodiversity and ecosystem services: a multilayered relationship. Trends in Ecology and Evolution, 2012, 27(1): 19-26. |
| [2] | Midgley G F. Biodiversity and Ecosystem function. Science, 2012, 335(6065): 174-175. |
| [3] | Altieri M A. The ecological role of biodiversity in agroecosystems. Agriculture, Ecosystems and Environment, 1999, 74(1/3): 19-31. |
| [4] | Ernoult A, Alard D. Species richness of hedgerow habitats in changing agricultural landscapes: are α and β diversity shaped by the same factors? Landscape Ecology, 2011, 26(5): 683-696. |
| [5] | Groot J C J, Jellema A, Rossing W A H. Designing a hedgerow network in a multifunctional agricultural landscape: balancing trade-offs among ecological quality, landscape character and implementation costs. European Journal of Agronomy, 2010, 32(1): 112-119. |
| [6] | Marshall E J P, Moonen A C. Field margins in northern Europe: their functions and interactions with agriculture. Agriculture, Ecosystems and Environment, 2002, 89(1/2): 5-21. |
| [7] | Bengtsson J, Ahnström J, Weibull A C. The effects of organic agriculture on biodiversity and abundance: a meta-analysis. Journal of Applied Ecology, 2005, 42(2): 261-269. |
| [8] | Duelli P, Obrist M K. Regional biodiversity in an agricultural landscape: the contribution of seminatural habitat islands. Basic and Applied Ecology, 2003, 4(2): 129-138. |
| [9] | Jackson L, van Noordwijk M, Bengtsson J, Foster W, Lipper L, Pulleman M, Said M, Snaddon J, Vodouhe R. Biodiversity and agricultural sustainagility: from assessment to adaptive management. Current Opinion in Environmental Sustainability, 2010, 2(1/2): 80-87. |
| [10] | Whittingham M J. The future of agri-environment schemes: biodiversity gains and ecosystem service delivery? Journal of Applied Ecology, 2011, 48(3): 509-513. |
| [11] | Green R E, Cornell S J, Scharlemann J P W, Balmford A. Farming and the fate of wild nature. Science, 2005, 307(5709): 550-555. |
| [12] | Mattison E H A, Norris K. Bridging the gaps between agricultural policy, land-use and biodiversity. Trends in Ecology and Evolution, 2005, 20(1): 610-616. |
| [13] | Tscharntke T, Clough Y, Wanger T C, Jackson L, Motzke I, Perfecto I, Vandermeer J, Whitbread A. Global food security, biodiversity conservation and the future of agricultural intensification. Biological Conservation, 2012, 151(1): 53-59. |
| [14] | Meek B, Loxton D, Sparks T H, Pywell R F, Pickett H, Nowakowski M. The effect of arable field margin composition on invertebrate biodiversity. Biological Conservation, 2002, 106(2): 259-271. |
| [15] | Douglas D J T, Vickery J A, Benton T G. Improving the value of field margins as foraging habitat for farmland birds. Journal of Applied Ecology, 2009, 46(2): 353-362. |
| [16] | Merckx T, Feber R E, Mclaughlan C, Bourn N A D, Parsons M S, Townsend M C, Riordan P, Macdonald D W. Shelter benefits less mobile moth species: the field-scale effect of hedgerow trees. Agriculture, Ecosystems and Environment, 2010, 138(3/4): 147-151. |
| [17] | Diekötter T, Wamser S, Wolters V, Birkhofer K. Landscape and management effects on structure and function of soil arthropod communities in winter wheat. Agriculture, Ecosystems and Environment, 2010, 137(1/2): 108-112. |
| [18] | Liu Y H, Duan M C, Yu Z R. Agricultural landscapes and biodiversity in China. Agriculture, Ecosystems and Environment, 2013, 166: 46-54. |
| [19] | Liu Y H, Axmacher J C, Wang C L, Li L T, Yu Z R. Ground beetles (Coleoptera: Carabidae) in the intensively cultivated agricultural landscape of Northern China-implications for biodiversity conservation. Insect Conservation and Diversity, 2010, 3(1): 34-43. |
| [20] | Liu Y H, Yu Z R, Gu W B, Axmacher J C. Diversity of carabids (Coleoptera, Carabidae) in the desalinized agricultural landscape of Quzhou county, China. Agriculture, Ecosystems and Environment, 2006, 113(1/4): 45-50. |
| [21] | José-María L, Armengot L, Blanco-Moreno J M, Bassa M, Sans F X. Effects of agricultural intensification on plant diversity in Mediterranean dryland cereal fields. Journal of Applied Ecology, 2010, 344: 832-840. |
| [22] | Gabriel D, Roschewitz I, Tscharntke T, Thies C. Beta diversity at different spatial scales: plant communities in organic and conventional agriculture. Ecological Applications, 2006, 16(5): 2011-2021. |
| [23] | Rundlöf M, Edlund M, Smith H G. Organic farming at local and landscape scales benefits plant diversity. Ecography, 2010, 33(3): 514-522. |
| [24] | Fahrig L, Baudry J, Brotons L, Burel F G, Crist T O, Fuller R J, Sirami C, Siriwardena G M, Martin J L. Functional landscape heterogeneity and animal biodiversity in agricultural landscapes. Ecology Letters, 2011, 14(2): 101-112. |
| [25] | Tscharntke T, Klein A M, Kruess A, Steffan-Dewenter I, Thies C. Landscape perspectives on agricultural intensification and biodiversity-ecosystem service management. Ecology Letters, 2005, 8(8): 857-874. |
| [26] | Gu Y F, Hu N, Ding S Y, Zhang L X. Community structure and dynamics of agricultural weeds under different land utilization regimes in Kaifeng area. Acta Ecologica Sinica, 2007, 344: 1359-1367. |
| [27] | Wang Y, Wang G, Du Y Q, Chen Q M, Gu Y F. Influence of edge effects on arthropods communities in agroforestry ecological systems. Acta Ecologica Sinica, 2011, 31(20): 6186-6193. |
| [28] | Song Y C. Vegetation Ecology. Shanghai: East China Normal University Press, 2001: 580-588. |
| [29] | Li X W. Floristic statistics and analyses of seed plants from China. Acta Botanica Yunnanica, 1996, 344: 363-384. |
| [30] | Wu Z Y. The areal-types of Chinese genera of seed plants. Acta Botanica Yunnanica, 1981, 5(S1): 1-139. |
| [31] | Robinson R A, Sutherland W J. Post-war changes in arable farming and biodiversity in Great Britain. Journal of Applied Ecology, 2002,39(1): 157-176. |
| [32] | Birrer S, Spiess M, Herzog F, Jenny M, Kohli L, Lugrin B. The Swiss agri-environment scheme promotes farmland birds: but only moderately. Journal of Ornithology, 2007, 148(S2): 295-303. |
| [33] | Kleijn D, Baquero R A, Clough Y, Díaz M, De Esteban J, Fernández F, Gabriel D, Herzog F, Holzschuh A, Jöhl R, Knop E, Kruess A, Marshall E J P, Steffan-Dewenter I, Tscharntke T, Verhulst J, West T M, Yela, J L. Mixed biodiversity benefits of agri-environment schemes in five European countries. Ecology Letters, 2006, 9(3): 243-254. |
| [34] | Jonason D, Andersson G K S, Öckinger E, Rundlöf M, Smith H G, Bengtsson J. Assessing the effect of the time since transition to organic farming on plants and butterflies. Journal of Applied Ecology, 2011, 48(3): 543-550. |
| [35] | Gabriel D, Sait S M, Hodgson J A, Schmutz U, Kunin W E, Benton T G. Scale matters: the impact of organic farming on biodiversity at different spatial scales. Ecology Letters, 2010, 13(7): 858-869. |
| [36] | Liu Y H, Yu Z R, Wang C L, Li L T, Chang H. The diversity of ground-dwelling beetles at cultivated land and restored habitats on the Bashang plateau. Acta Ecologica Sinica, 2011, 31(2): 465-473. |
| [37] | Yu Z R, Gu W B, Hu D X. On landscape pattern and biodiversity in rural areas of Jianghan plain-taking two villages as a case study. Resources Science, 2000, 22(2): 19-23. |
| [38] | Ma S C, Yuan D F, Yang S J, Wang H Z. Plant diversity in field margin systems in the hilly regions of northern Henan. Chinese Journal of Eco-Agriculture, 2010, 344: 815-819. |
| [39] | Wu J G. Landscape ecology-concepts and theories. Chinese Journal of Ecology, 2000, 19(1): 42-52. |
| [40] | Damschen E I, Haddad N M, Orrock J L, Tewksbury J J, Levey D J. Corridors increase plant species richness at large scales. Science, 2006, 313(5791): 1284-1286. |
| [41] | Wu J G. The key research topics in landscape ecology. Acta Ecologica Sinica, 2004, 24(9): 2074-2076. |
| [42] | Delattre T, Vernon P, Burel F. An agri-environmental scheme enhances butterfly dispersal in European agricultural landscapes. Agriculture, Ecosystems and Environment, 2013, 166: 102-109. |
| [43] | Donald P F, Evans A D. Habitat connectivity and matrix restoration: the wider implications of agri-environment schemes. Journal of Applied Ecology, 2006, 43(2): 209-218. |
| [44] | Cai N H, L G Q, Zhu C F, Huang Y X, Li J N, Zhao W D. A comparison study on the community structure between artificial and natural forests of Pinus yunnanensis. Journal of Northwest Forestry University, 2007, 22(2): 1-4. |
| [45] | Oria-de-Rueda J A, Hernández-Rodríguez M, Martín-Pinto P, Pando V, Olaizola J. Could artificial reforestations provide as much production and diversity of fungal species as natural forest stands in marginal Mediterranean areas? Forest Ecology and Management, 2010, 260(2): 171-180. |
| [26] | 谷艳芳, 胡楠, 丁圣彦, 张丽霞. 开封地区不同土地利用方式农田杂草群落结构及动态. 生态学报, 2007, 344: 1359-1367. |
| [27] | 汪洋, 王刚, 杜英琪, 陈绮铭, 谷艳芳. 农林复合生态系统防护林斑块边缘效应对节肢动物的影响. 生态学报, 2011, 31(20): 6186-6193. |
| [28] | 宋永昌. 植被生态学. 上海:华东师范大学出版社, 2001: 580-588. |
| [29] | 李锡文. 中国种子植物区系统计分析. 云南植物研究, 1996, 344: 363-384. |
| [30] | 吴征镒. 中国种子植物属的分布区类型. 云南植物研究, 1981, 5(增刊): 1-139. |
| [36] | 刘云慧, 宇振荣, 王长柳, 李良涛, 常虹. 坝上地区农田和恢复生境地表甲虫多样性. 生态学报, 2011, 31(2): 465-473. |
| [37] | 宇振荣, 谷卫彬, 胡敦孝. 江汉平原农业景观格局及生物多样性研究——以两个村为例. 资源科学, 2000, 22(2): 19-23. |
| [38] | 马守臣, 原东方, 杨慎骄, 王和洲. 豫北低山丘陵区农田边界系统植物多样性的研究. 中国生态农业学报, 2010, 344: 815-819. |
| [39] | 邬建国. 景观生态学——概念与理论. 生态学杂志, 2000, 19(1): 42-52. |
| [41] | 邬建国. 景观生态学中的十大研究论题. 生态学报, 2004, 24(9): 2074-2076. |
| [44] | 蔡年辉, 李根前, 朱存福, 黄永祥, 李俊楠, 赵文东. 云南松人工林与天然林群落结构的比较研究. 西北林学院学报, 2007, 22(2): 1-4. |