 2014, Vol. 34
2014, Vol. 34文章信息
- 王振海, 殷秀琴, 蒋云峰
- WANG Zhenhai, YIN Xiuqin, JIANG Yunfeng
- 长白山苔原带土壤动物群落结构及多样性
- Structure and diversity of soil fauna communities in the tundra of the Changbai Mountains, China
- 生态学报, 2014, 34(3): 755-765
- Acta Ecologica Sinica, 2014, 34(3): 755-765
- http://dx.doi.org/10.5846/stxb201306081438
-
文章历史
- 收稿日期:2013-6-8
- 修订日期:2013-11-11






2. 吉林省动物资源保护与利用重点实验室, 长春 130024
2. Jilin Key Laboratory of Animal Resource Conservation and Utilization, Changchun 130024, China
土壤动物是陆地生态系统的重要组成部分,联系着地上和地下生态系统[1, 2],在物质循环和能量流动等方面起着重要的作用[3, 4],对植被、土壤等环境因子的变化有一定的响应[5, 6]。近年来,土壤动物的生态学研究越来越受到学者们的关注[7, 8, 9]。
长白山属于温带大陆性季风气候,是欧亚大陆北半部最具代表性的典型综合自然体和物种基因库,是我国重要的地理单元。然而,现在的长白山生态系统是火山喷发后自然演替而成,土壤层较薄,生态环境比较脆弱。近年来学者们加强了对长白山生态系统的研究,在火山遥感监测、植被与环境的关系等方面都进行了比较全面的研究;土壤动物方面,张荣祖、陈鹏等最早对长白山土壤动物进行了调查研究[10, 11],殷秀琴等对长白山红松阔叶混交林土壤动物生态分布进行了研究[12],佟富春等对长白山土壤动物群落随海拔的变化进行研究[13, 14]。 长白山苔原带植被以小灌木和苔藓、地衣为主,生态环境更为脆弱,对于土壤动物的研究较少,仅陈鹏等在1983年进行了研究[15, 16]。本文通过对上部和下部苔原带土壤动物的研究,旨在较深入的探讨长白山苔原带土壤动物群落的组成、结构和多样性特征,为长白山生态系统的保护和可持续发展提供科学依据。因此具有重要的生态学意义。
1 研究区概况长白山苔原带位于海拔2000 m以上,属于火山椎体的上部,寒冷而风大。每年至少有6个月的时间被积雪覆盖。根据植被情况将苔原带分上下两个亚带。
上部苔原亚带位于海拔2300 m以上,主要特征为寒冷多雾、降水多以及风大。1月平均气温低于-20 ℃,7月为8—10 ℃,无霜期不足60 d。土壤为高山寒漠土[16],植被总覆盖度为40%[17],以苔藓地衣为主, 伴有珠芽拳蓼(Bistorta vivipara)、萝蒂草(Lloydia serotina)和轮叶马先嵩(Pedicularsi verticillat)等。
下部苔原亚带位于2000—2300 m,寒冷而风大,≥10 ℃活动积温为300—500 ℃,年降水量为1100—1300 mm,土壤为高山苔原土[16], 植被总覆盖度为100%[17],主要植物有仙女木(Dryas octopetala)、牛皮杜鹃(Rhondodendron aureum)、笃斯越桔(Vaccinium uliginosum)、毛毡杜鹃(Rhondodendron confertissimum) 等。
2 研究方法 2.1 土壤动物的采集与鉴定2009年5月、7月、9月在长白山苔原带上、下两个亚带分别设置样地进行取样。大型土壤动物按25 cm×25 cm的样方进行取样,中小型土壤动物按10 cm×10 cm样方取样,分别设6个重复样方;每个样方分别取凋落物层、0—5 cm 、5—10 cm、10—15 cm土层;共取288个样品。大型土壤动物采用手拣法就地获取,中小型土壤动物带回室内采用Tullgren分离。收集到所有的土壤动物放到75%的酒精中保存。
土壤动物在体视镜( OLYMPUS SZX16)下的鉴定,一般鉴定到科,少数鉴定到目水平。
2.2 数据处理土壤动物群落多度等级的划分:个体数占总捕获量10%以上的为优势类群,个体数占总捕获量1%—10%之间的为常见类群,个体数占总捕获量的1%以下的为稀有类群。
土壤动物多样性特征主要采用Shannon-Wiener多样性指数H′、Margalef丰富度指数D、Pieluo均匀度指数J和Simpson优势度指数C来计算。
Shannon-Wiener多样性指数H′
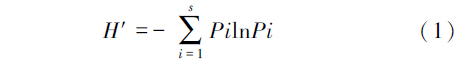
Margalef 丰富度指数D

Pielou 均匀度指数J

Simpson优势度指数C
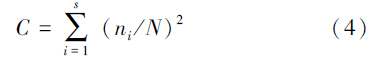
式中,s代表土壤动物的总类群数,Pi代表第i类土壤动物的个体数与土壤动物总个体数的比,ni为第i类群个体数,N为群落的总个体数。
土壤动物群落的相似性采用Sorenson相似性系数S1(定性分析群落相似性,考虑物种组成的差异性)和Morisita-Horn相似性系数S2(定量分析群落相似性,考虑到每个物种的丰度)来表示。
Sorenson相似性系数S1:
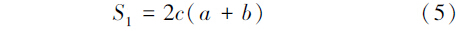
式中,a为A生境全部类群数,b为B生境全部类群数,c为A、B两生境共有的类群数。计算值在0.75—1.00之间为极相似,在0.50—0.74之间为中等相似,在0.25—0.49之间为中等不相似,在0—0.24之间为极不相似。
Morisita-Horn相似性系数S2
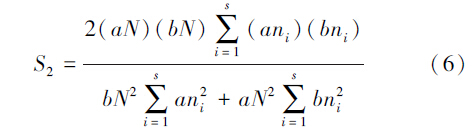
式中,s为A、B两生境所有的类群数;aN和bN分别为A、B生境中所有类群的总个体数;ani和bni分别A、B生境中第i个类群的个体数。
采用EXCEL2003进行图表绘制和计算土壤动物多样性指数,用SPSS18.0进行方差分析和均值比较。
3 研究结果 3.1 土壤动物群落组成特征共获得土壤动物10286只,计37类(表1),隶属2门6纲16目26科。优势类群为甲螨亚目、节跳虫科和球角跳虫科3类,占苔原带总捕获量的69.86%,常见类群为革螨亚目、辐螨亚目、山跳虫科和线蚓科4类,占总数的27.57%,其余30类为稀有类群,占2.57%。
上部苔原亚带捕获4535只,占44.09%,24类;优势类群为节跳虫科,占上部苔原带总数的57.55%,常见类群为甲螨亚目、革螨亚目、辐螨亚目、山跳虫科、球角跳虫科和线蚓科6类,占39.43%,其余17类为稀有类群,占3.02%。下部苔原亚带共捕获土壤动物32类,5751只,占土壤动物总数的55.91%。优势类群为甲螨亚目、辐螨亚目、节跳虫科和球角跳虫科4类,占该带的77.12%,常见类群为革螨亚目、山跳虫科和线蚓科3类,占20.67%,其余25类为稀有类群,占2.21%。说明下部苔原亚带生境更适宜土壤动物生存,土壤动物群落组成较复杂。
两个亚带土壤动物的优势类群和常见类群存在一定差异,并且上部苔原亚带不同类群所占比例差异较大,仅节跳虫科占上部苔原亚带的一半以上,远大于其在下部苔原亚带所占的比例;两个亚带的稀有类群在组成上存在一定的差异,鳞跳虫科、花萤科幼虫、地幺蚣科、啮虫科、夜蛾科幼虫、地蜈蚣科、象甲科、叩甲科幼虫、萤科幼虫、拟态蛛科、叶甲科幼虫、马陆目、幺蚣科土壤动物仅出现在下部苔原亚带,这对该生境有一定的指示作用。
|
上部苔原亚带
The upper tundra | % |
下部苔原亚带
The lower tundra | % |
合计
Total | % | |
| 节跳虫科Isotomidae | 2610 | 57.55 | 2075 | 36.08 | 4685 | 45.55 |
| 甲螨亚目Oribatida | 431 | 9.5 | 1038 | 18.05 | 1469 | 14.28 |
| 球角跳虫科Hypogastruridae | 300 | 6.62 | 732 | 12.73 | 1032 | 10.03 |
| 辐螨亚目Actinedida | 396 | 8.73 | 590 | 10.26 | 986 | 9.59 |
| 山跳虫科Pseudachorutidae | 382 | 8.42 | 522 | 9.08 | 904 | 8.79 |
| 线蚓科Enchytraeidae | 184 | 4.08 | 491 | 8.54 | 675 | 6.56 |
| 革螨亚目Gamasida | 95 | 2.09 | 176 | 3.06 | 271 | 2.63 |
| 短角亚目幼虫Brachycera larvae | 12 | 0.26 | 28 | 0.49 | 40 | 0.39 |
| 芒角亚目幼虫Aristocera larvae | 23 | 0.51 | 16 | 0.28 | 39 | 0.38 |
| 长角亚目幼虫Nematocera larvae | 21 | 0.46 | 1 | 0.02 | 22 | 0.21 |
| 石蜈蚣科Lithobiidae | 13 | 0.29 | 8 | 0.14 | 21 | 0.2 |
| 管蓟马科Phloeothripidae | 16 | 0.35 | 4 | 0.07 | 20 | 0.19 |
| 圆跳虫科Sminthuridae | 14 | 0.31 | 14 | 0.14 | ||
| 长角跳虫科Entomobryidae | 8 | 0.18 | 6 | 0.1 | 14 | 0.14 |
| 花萤科幼虫Cantharidae larvae | 13 | 0.23 | 13 | 0.13 | ||
| 隐翅虫科Staphyinidae | 1 | 0.02 | 10 | 0.17 | 11 | 0.11 |
| 隐翅虫科幼虫Staphyinidae laevae | 8 | 0.18 | 1 | 0.02 | 9 | 0.09 |
| 蟹蛛科Thomisidae | 6 | 0.13 | 1 | 0.02 | 7 | 0.07 |
| 地蜈蚣科Geophilidae | 6 | 0.1 | 6 | 0.06 | ||
| 步甲科Carabidae | 3 | 0.07 | 2 | 0.03 | 5 | 0.05 |
| 缨翅目Thysanoptera | 1 | 0.02 | 4 | 0.07 | 5 | 0.05 |
| 鳞翅目幼虫Lepidoptera larvae | 3 | 0.07 | 1 | 0.02 | 4 | 0.04 |
| 象甲科Curculionidae | 4 | 0.07 | 4 | 0.04 | ||
| 啮虫科Psocidae | 4 | 0.07 | 4 | 0.04 | ||
| 地幺蚣科Geophilellidae | 4 | 0.07 | 4 | 0.04 | ||
| 鳞跳虫科Tomoceridae | 3 | 0.05 | 3 | 0.03 | ||
| 萤科幼虫 Lampyridae larvae | 3 | 0.05 | 3 | 0.03 | ||
| 摇蚊科Chironomidae | 3 | 0.07 | 3 | 0.03 | ||
| 步甲科幼虫Carabidae larvae | 2 | 0.04 | 2 | 0.02 | ||
| 叩甲科幼虫Elateridae larvae | 2 | 0.03 | 2 | 0.02 | ||
| 幺蚣科Scolopendrellidae | 2 | 0.03 | 2 | 0.02 | ||
| 拟态蛛科Mimetidae | 1 | 0.02 | 1 | 0.01 | ||
| 马陆目Juliformia | 1 | 0.02 | 1 | 0.01 | ||
| 叶甲科幼虫 Chrysomelidae larvae | 1 | 0.02 | 1 | 0.01 | ||
| 夜蛾科幼虫Noctuidae larvae | 1 | 0.02 | 1 | 0.01 | ||
| 长蝽科Lygaeidae | 1 | 0.02 | 1 | 0.01 | ||
| 总个体数 Total individual | 4535 | 5751 | 10286 | |||
| 类群数 Group number | 24 | 32 | 37 |
由图1可以看出,无论在哪个月份下部苔原亚带的个体数均多于上部苔原亚带,除7月外,下部苔原亚带的类群数也多于上部苔原亚带。两个亚带土壤动物的个体数都是先减后增,上部苔原亚带从5月的2440个降到7月的796个,9月时上升到1299个,而下部苔原亚带各月份土壤动物个体数分别为2696个、1086个和1969个;两个亚带类群数均呈现减少的趋势,上部苔原亚带在5月和7月保持不变,之后减少,下部苔原亚带在5月相对多,7月和9月没有变化。经多因素方差分析(表3)和LSD多重比较分析,5月土壤动物的个体数显著大于7月(P﹤0.001)和9月(P=0.012),而两个亚带间没有显著差异。表明土壤动物的个体数随生境和时间变化有很大波动,而类群数变化较小。

|
| 图 1 土壤动物的个体数和类群数 (平均值±标准误) Fig. 1 Individual number and group number of soil fauna (Mean±SE) |
为了更好地了解不同类群土壤动物的水平结构,选取研究区甲螨亚目、革螨亚目、辐螨亚目、节跳虫科、山跳虫科、球角跳虫科和线蚓科7个主要类群进行比较(图2)。由图2可以看出,下部苔原亚带各土壤动物类群的个体数要大于上部苔原亚带,特别是在5月份下部苔原亚带球角跳虫科和山跳虫科的个体数显著大于上部苔原亚带(P=0.038,P=0.003)。在土壤动物的时间变化方面,螨类在7月份数量最少,9月份数量最多,特别是甲螨在上部苔原亚带和下部苔原亚带两个生境都有显著的变化(P=0.002,P=0.014);而弹尾目在5月份个体数最多,尤其是在下部苔原亚带,节跳虫科、球角跳虫科和山跳虫科5月份都显著大于其它两个月份(P=0.029,P=0.031,P=0.009);线蚓科在7月份个体数最多。说明不同类群土壤动物的个体数存在生境和时间上的波动。
3.3 土壤动物的垂直结构及时间动态土壤动物的个体数和类群数总体上随土壤深度的增加而降低,不同月份的垂直分布情况有一定差异(表2)。从个体数来看,上部苔原亚带在5月凋落物层中最多,下部苔原亚带也同样是在5月个体数最多,但出现在0—5 cm层。从类群数来看,上部苔原亚带在7月凋落物层中最多(18类),下部苔原亚带在7和9月最多(15类),但出现在0—5 cm层。经多因素方差分析(表3)表明土壤动物个体数在各层和月份之间存在显著差异,LSD多重比较分析显示0—5 cm层显著多于其它3层(P﹤0.001),并且取样层与时间、取样层与生境以及三者的交互作用都对土壤动物个体数有显著的影响;LSD多重比较分析显示0—5 cm层土壤动物类群数也显著多于凋落物层、5—10 cm层、10—15 cm层(P=0.024,P﹤0.001,P﹤0.001),取样层与时间、取样层与生境的交互作用都对土壤动物类群数有显著的影响。
| 月份
Month | 个体数 Individual number | 类群数 Group number | |||||||
| 凋落物层
Litter layer | 0—5 cm | 5—10 cm | 10—15 cm | 凋落物层
Litter layer | 0—5 cm | 5—10 cm | 10—15 cm | ||
| 上部苔原亚带 | 5 | 1132 | 864 | 148 | 296 | 17 | 13 | 9 | 9 |
| The upper tundra | 7 | 347 | 350 | 70 | 29 | 18 | 14 | 7 | 5 |
| 9 | 165 | 677 | 262 | 195 | 9 | 15 | 7 | 6 | |
| 下部苔原亚带 | 5 | 200 | 2051 | 279 | 166 | 12 | 14 | 11 | 11 |
| The lower tundra | 7 | 438 | 391 | 222 | 35 | 14 | 15 | 10 | 6 |
| 9 | 282 | 1120 | 283 | 284 | 9 | 15 | 14 | 12 | |

|
| 图 2 主要土壤动物类群的水平结构特征 (平均值±标准误) Fig. 2 Horizontal structure of main soil fauna (Mean±SE) |
| 个体数Individual number | 类群数Group number | |||||
| df | F | P | df | F | P | |
| 土层 Layer | 3 | 20.612 | ﹤0.001 | 3 | 18.113 | ﹤0.001 |
| 月份 Month | 2 | 9.92 | ﹤0.001 | 2 | 2.806 | 0.064 |
| 生境 Habitat | 1 | 1.834 | 0.178 | 1 | 9.464 | 0.003 |
| 土层×月份 Layer×Month | 6 | 3.898 | 0.001 | 6 | 6.23 | ﹤0.001 |
| 土层×生境 Layer×Habitat | 3 | 5.027 | 0.003 | 3 | 3.23 | 0.025 |
| 月份×生境 Month×Habitat | 2 | 0.197 | 0.822 | 2 | 0.193 | 0.825 |
| 土层×月份×生境 Layer×Month×Habitat | 6 | 3.472 | 0.003 | 6 | 0.22 | 0.97 |
土壤动物主要类群在各层之间也存在较大差异 (表4)。对于螨类,上部苔原亚带9月甲螨亚目(P﹤0.01)和革螨亚目(P﹤0.05)在0—5 cm土层显著高于其它几层,下部苔原亚带5月甲螨亚目、革螨亚目、辐螨亚目以及9月的甲螨亚目也有同样的表现(P=0.03,P=0.011,P=0.027,P=0.022),其它月份各层之间没有显著差异(P﹥0.05)。弹尾目只有下部苔原亚带存在显著差异,5月的球角跳虫科以及各月的节跳虫科和山跳虫科在各层之间存在显著差异(P=0.001,P=0.001,P=0.036,P=0.005,P=0.001,P=0.04,P=0.009);两个亚带中线蚓科均在0—5 cm土层个体数量最多。从总体上来看,上部苔原亚带土壤动物的垂直分布比下部苔原亚带容易受时间变化的影响,不同土壤动物的垂直分布随时间和生境的不同而有所不同。
| 月份
Month | 上部苔原亚带 The upper tundra | 下部苔原亚带 The lower tundra | |||||||
| 凋落物层
Litter layer | 0—5 cm | 5—10 cm | 10—15 cm | 凋落物层
Litter layer | 0—5 cm | 5—10 cm | 10—15 cm | ||
| 同行内不同小写字母表示样地间差异显著(P﹤0.05) | |||||||||
| 甲螨亚目 | 5 | 14.83±5.78 | 11±3.44 | 5.83±3.03 | 1.67±1.17 | 7.67±2.46b | 24.17±6.68a | 4.50±1.20b | 4±2.25b |
| Oribatida | 7 | 2.67±1.15 | 1.50±1.15 | 0±0 | 0±0 | 8.17±4.56 | 2.80±0.97 | 0.80±0.80 | 0±0 |
| 9 | 60±3.36b | 265±3.57a | 1.83±0.91b | 0±0b | 32±14.25ab | 72.67±26.10a | 8.67±3.29b | 8.17±4.32b | |
| 革螨亚目 | 5 | 5.67±3.43 | 0.33±0.33 | 0.50±0.50 | 0.17±0.17 | 3±1.13b | 7.80±2.27a | 1.60±1.17b | 1±0.63b |
| Gamasida | 7 | 1.50±0.76 | 2.67±1.86 | 0±0 | 0±0 | 1.67±0.67 | 2.80±1.39 | 0.60±0.25 | 0±0 |
| 9 | 0.50±0.34b | 4.50±1.95a | 0±0b | 0±0b | 0.50±0.50 | 7.50±3.13 | 2.17±1.28 | 2.83±1.05 | |
| 辐螨亚目 | 5 | 3±1.75 | 3.83±0.98 | 3.83±2.12 | 1±0.68 | 3.50±2.43b | 15.67±6.09a | 3.50±1.61b | 1.17±0.48b |
| Actinedida | 7 | 7.50±5.03 | 10.83±7.39 | 0.67±0.67 | 0.33±0.33 | 2.83±1.14 | 8±3.26 | 8.80±4.59 | 0.67±0.33 |
| 9 | 7.83±6.05 | 7.33±4.04 | 11.33±7.01 | 8.50±5.41 | 6.83±3.61 | 23.83±6.37 | 11.67±8.12 | 13.33±4.81 | |
| 节跳虫科 | 5 | 140.67±70.57 | 108.83±65.99 | 5.33±1.23 | 43±40.42 | 6.17±1.78b | 195.50±66.83a | 11±3.10b | 4.50±1.23b |
| Isotomidae | 7 | 15.67±6.38 | 21.50±17 | 3.17±0.95 | 0±0 | 35±10.38a | 10.33±3.60b | 19.40±10.60ab | 1.40±0.51b |
| 9 | 6.67±2.43 | 55.67±24.52 | 23.17±7.17 | 11.33±3.04 | 2.67±1.67b | 50±17.60a | 6.33±4.23b | 7±4.51b | |
| 山跳虫科 | 5 | 6.67±2.82 | 8.33±3.06 | 2.67±1.89 | 0.67±0.33 | 6.50±1.80b | 38.67±11.06a | 7.17±2.65b | 1.17±0.40b |
| Pseudachorutinae | 7 | 15±5.24 | 8.17±2.52 | 6.33±4.62 | 3.67±3.67 | 10.50±2.83a | 6±2.22ab | 3.50±2.20b | 0.80±0.37b |
| 9 | 2.17±0.98 | 4.17±0.7 | 2.50±1.02 | 3.33±2.58 | 1.17±0.65b | 8.83±3.03a | 1.33±0.49b | 1.50±1.12b | |
| 球角跳虫科 | 5 | 12±5.03 | 7.33±3.11 | 3±1.03 | 2±1.23 | 4±1.39b | 46.67±11.38a | 9.67±3.20b | 11.83±5.24b |
| Hypogastruridae | 7 | 5.67±2.95 | 4.5±2.94 | 0.33±0.21 | 0.17±0.17 | 6.33±2.63 | 8±2.89 | 4.67±1.89 | 2.20±1.07 |
| 9 | 1.33±0.49 | 6.50±1.73 | 1.50±1.12 | 5.67±2.29 | 2.83±2.64 | 10.33±5.46 | 63.33±5.16 | 9.50±3.27 | |
| 线蚓科 | 5 | 2.33±1.38 | 2±0.93 | 3±1.31 | 0.5±0.25 | 0.33±0.21 | 10±4.94 | 8.67±6.89 | 2.83±1.08 |
| Enchytraeidae | 7 | 5.33±2.65 | 7±3.93 | 0.67±0.33 | 0.5±0.25 | 6.81±3 | 25.81±12 | 3.16±1.35 | 1.2±0.5 |
| 9 | 0±0 | 3.45±1.28 | 2.83±1.14 | 2.83±1.20 | 0.33±0.21 | 10.15±5.33 | 8.83±2.27 | 3.83±1.22 | |
研究表明,下部苔原亚带土壤动物多样性高于上部苔原亚带,在下部苔原亚带共获得土壤动物32类,而在上部苔原亚带获得24类;上、下两个亚带土壤动物多样性随月份变化都有递减的趋势。
图3显示的是两个亚带土壤动物多样性特征指数的情况。各月份,上部苔原亚带土壤动物的多样性指数和均匀度指数均低于下部苔原亚带。7月,上部苔原亚带土壤动物的丰富度指数高于下部苔原亚带,5月和9月低于下部苔原亚带。各月份,上部苔原亚带土壤动物的优势度指数高于下部。下部苔原亚带的植被组成较复杂,能为土壤动物提供更多的食物和栖息环境,土壤动物组成较丰富,所以多样性指数较高;并且上部苔原亚带各土壤动物的所占比例差异较大,仅优势类群节跳虫科就占该亚带的57.55%,导致优势度指数较高。
上部和下部苔原亚带土壤动物的多样性指数、均匀度指数和优势度指数均为7月﹥9月﹥5月,而优势度指数在5月最高。虽然7月土壤动物类群数不是最多,但是各类群之间个体数量差异最小,群落组成更加均匀,所以多样性指数最高;而5月土壤动物类群数虽然最多,但土壤动物的个体数量差异很大,尤其是节跳虫科和甲螨亚目占有很大的比例,多样性指数最低,优势度指数最高。说明生境差异和时间变化对土壤动物的多样性有一定的影响。

|
| 图 3 土壤动物的多样性特征 (平均值±标准误) Fig. 3 Diversity characteristics of soil fauna (Mean±SE) |
苔原带土壤动物群落的相似性特征如表5所示。各月份苔原带两个亚带之间土壤动物的Sorenson相似性均处于中等相似水平,表明两个亚带之间土壤动物群落物种组成存在一定的相似性;5月和7月两个亚带之间土壤动物群落Morisita-Horn系数均值在0.9以上,9月为0.78,在5月和7月下部苔原亚带土壤动物个体数比上部分别多256只和290只,而9月份多670只,表明9月两个亚带之间土壤动物各类群的相对数量差异比5月和7月时大。
| 5月
May | 7月
July | 9月
September | |
| Sorenson相似性 | 0.68 | 0.67 | 0.65 |
| Morisita-Horn相似性 | 0.90 | 0.91 | 0.78 |
土壤动物组成和分布受植被组成特征、湿度和温度等多种因素的影响[18, 19, 20],不同生境中土壤动物的组成和结构存在差异[21, 22],植被条件越优越的生境,土壤动物的多样性越高[23, 24]。上部苔原亚带以苔藓地衣为主,下部苔原亚带以低矮灌木为主,植物种类也相对丰富,更适宜土壤动物的取食与繁殖;另外,下部苔原亚带海拔低,气温高,土壤养分含量更高,较适宜土壤动物的生存与活动。因此,上部苔原亚带土壤动物群落的类群数和多样性等均小于下部苔原亚带。王邵军等[25]在武夷山研究表明土壤动物的类群数和多样性均随海拔升高而降低,与此研究结论相近。
两个亚带土壤动物的Sorenson和Morisita-Horn相似性指数显著不同,也表明两个亚带土壤动物在物种组成和各类群的相对数量均存在一定的差异。螨类、跳虫和线蚓科分布广、适应性强,在两个亚带均为优势类群或常见类群,但是不同亚带其个体数量不同;稀有类群在两个亚带的变化较大,特有类群分别为4和13类。黄旭等[27]在川西高山研究表明不同植被类型下的土壤动物组成不同,Zhu Xinyu等[26]的研究也证明不同的植被条件能够影响土壤动物群落的物种组成和各类群的数量。因此,生境条件对土壤动物群落的组成和多样性有一定的影响。
4.2 土壤动物多样性的时间变化不同季节的降水、气温以及植被的差异是影响土壤动物数量和多样性的主要因素[27, 28, 29]。研究发现,各时段土壤动物的类群数变化不大,但是土壤动物数量变化很大。7月与其它两个月份相比,土壤动物主要类群螨类和跳虫的数量显著减少,而一些稀有类群数量有所增加,虽然土壤动物个体数在7月份最少,而优势度指数降低,多样性指数、丰富度指数和均匀度指数最高。张洪芝等[30]在若尔盖高寒草甸地区的研究表明,土壤动物的个体数也在7月份最少,而多样性指数在10月份最大,与此研究结果存在差异,这可能是地区差异的原因。
4.3 土壤动物的垂直结构土壤动物容易受自然灾害和干扰因素的影响[31]。长白山苔原带温度低,植被低矮,凋落物层土壤动物缺乏相对稳定的栖息环境,容易受到大风和降水等因素的影响,因此,凋落物层土壤动物的个体数和类群数少于0—5 cm土层。
土壤动物的垂直结构受土壤理化性质以及植被条件的影响[18, 32],土壤温湿度的季节变化是影响土壤动物垂直结构季节动态的主要因子[33, 34],而不同土壤动物对土壤温湿度等环境因子的响应不同[35, 36]。上部和下部苔原带植被组成和土壤养分含量差异较大,由海拔变化引起的温湿度变化也存在差异,使各层土壤动物种类组成和数量存在差异,如5月和7月甲螨亚目在两个亚带的凋落物层最多,而9月份凋落物层少于0—5 cm土层;5月份节跳虫科在上部苔原亚带以凋落物层最多,而下部苔原亚带以0—5 cm土层最多。
本研究区位于长白山苔原带,由于自然环境非常恶劣,土壤动物的多样性要低于其他自然带[26, 37]。但是关于苔原带土壤动物与环境因子的相互作用机制研究较少,因此,有待于进一步的研究来揭示其规律。
5 结论(1)在长白山苔原带两个亚带,优势类群为甲螨亚目、节跳虫科和球角跳虫科3类,常见类群为革螨亚目、辐螨亚目、山跳虫科和线蚓科4类,其余30类为稀有类群。两个亚带所捕获个体数分别占总数的44.09%和55.91%,类群数分别为24类和32类。下部苔原亚带土壤动物群落结构较复杂。
(2)土壤动物的个体数随生境和时间变化有很大波动,而类群数变化较小。下部苔原亚带土壤动物的个体数、类群数要多于上部苔原亚带。两个亚带土壤动物的个体数在7月最少,而类群数随着时间的变化有减少的趋势。
(3)上部苔原亚带5月个体数和类群数及7月类群数均在凋落物层最高,下部苔原亚带7月的个体数也有相同的表现,其余均在0—5 cm土层最高。0—5 cm土层向下,土壤动物的个体数和类群数随着土层深度的增加而减少。主要土壤动物的垂直分布随时间和生境的不同而存在差异。
(4)生境差异和时间变化对土壤动物的多样性特征有一定的影响,下部苔原亚带土壤动物多样性高,两个亚带土壤动物多样性随月份变化都呈递减的趋势。下部苔原亚带土壤动物的多样性指数、丰富度指数和均匀度指数均高于上部苔原亚带,优势度则相反。多样性指数、丰富度指数和均匀度指数的变化为7月最高,而5月最低,优势度指数在5月份最高。
(5)Sorenson相似性表明两个亚带土壤动物群落物种组成上存在一定的差异;Morisita-Horn相似性表明,两个亚带之间土壤动物各类群的相对数量差异在9月最大。
| [1] | Lavelle P, Decaëns T, Aubert M, Barot S, Blouin M, Bureau F, Margerie P, Mora P, Rossic JP. Soil invertebrates and ecosystem services. European Journal of Soil Biology, 2006, 42: S3-S15. |
| [2] | Wardle D A, Bardgett R D, Klironomos J N, Setälä H, van der Putten W H, Wall D H. Ecological linkages between aboveground and belowground biota. Science, 2004, 304(5677): 1629-1633. |
| [3] | Dong W H, Yin X Q. Transformation of carbon and nitrogen by earthworms in the decomposition processes of Broad-leaved litters. Chinese Geographical Science, 2007, 17(2): 166-172. |
| [4] | Xia L, Wu F Z, Yang W Q, Tan B. Contribution of soil fauna to the mass loss of Betula albosinensis leaf litter at early decomposition stage of subalpine forest litter in western Sichuan. Chinese Journal of Applied Ecology, 2012, 23(2): 301-306. |
| [5] | Song B, Yin X Q, Zhang Y, Dong W H. Dynamics and relationship of Ca, Mg, Fe in litter, soil fauna and soil in Pinus koraiensis-Broadleaf mixed forest. Chinese Geographical Science, 2008, 18(3): 284-290. |
| [6] | Gao M X, Zhang X P. Fluctuation of soil fauna community during defoliation decomposition under lime and EM treatment. Acta Ecologica Sinica, 2011, 31(1): 164-174. |
| [7] | Yin X Q, Song B, Dong W H, Xin W D, Wang Y Q. A review on the eco-geography of soil fauna in China. Journal of Geographical Sciences, 2010, 20(3): 333-346. |
| [8] | Xin W D, Yin, X Q, Song B. Contribution of soil fauna to litter decomposition in Songnen sandy lands in northeastern China. Journal of Arid Environments, 2012, 77: 90-95. |
| [9] | Wang Q Y, Zhou D M, Cang L, Li L Z, Zhu H W. Indication of soil heavy metal pollution with earthworms and soil microbial biomass carbon in the vicinity of an abandoned copper mine in Eastern Nanjing, China. European Journal of Soil Biology, 2009, 45(3): 229-234. |
| [10] | Chen P, Fu D Y. A preliminary approach to the effect of soil animals on the cycling of materials in the Changbai Shan mountain. Acta Ecologica Sinica, 1984, 4(2): 172-180. |
| [11] | Zhang R Z, Yang M X, Chen P, Zhang T W. Preliminary investigation of soil animal in ecosystems of northern of Changbai Mountains. Forest Ecosystem Research, 1980(1): 133-152. |
| [12] | Yin X Q, Jiang Y F, Tao Y, An J C, Xin W D. Ecogeographical distribution of soil fauna in Pinus koraiensis mixed Broad-leaved forest of Changbai Mountains. Scientia Geographica Sinica, 2011, 31(8): 935-940. |
| [13] | Tong F C, Jin Z D, Wang Q L, Xiao Y H. Co-occurrence of soil fauna communities with change in altitude on the northern slope of Changbai Mountain. Chinese Journal of Applied Ecology, 2003, 14(10): 1723-1728. |
| [14] | Tong F C, Wang Q L, Liu X S, Xiao Y H. Dynamics of soil fauna communities during succession process of secondary forests in Changbai Mountain. Chinese Journal of Applied Ecology, 2004, 15(9): 1531-1535. |
| [15] | Chen P, Zhang Y. Periglacial environment and soil fauna on northern slope of Changbai Mountain. Scientia Geographica Sinica, 1983, 3(2): 133-140. |
| [16] | Chen P, Yang C B, Zhang Y. Ecogeographical distribution of soil fauna in the Alpine tundra of northern of Changbai Mountains. Forest Ecosystem Research, 1983, (3): 119-132. |
| [17] | Qian J J. Investigation report of typical vegetation of different belts on northern of Changbai Mountains // Zhang Z Z. Research on Geosystems of the Changbai Mountains(VOL.1). Changchun: Northern Normal University Press, 2010: 89-115. |
| [18] | Liu J L, Yin X Q, Qiu L L. Large-sized soil fauna and soil factors in Zuojia Nature Reserve. Acta Pedologica Sinica, 2008, 45(1): 130-136. |
| [19] | Oxbrough A, Irwin S, Kelly T C, O'Halloran J. Ground-dwelling invertebrates in reforested conifer plantations. Forest Ecology and Management, 2010, 259(10): 2111-2121. |
| [20] | Sandrine S, Nadia A, Lorenzo F, Roberto Z. Relationships between soil fauna communities and humus forms: Response to forest dynamics and solar radiation. Soil Biology and Biochemistry, 2008, 40(7): 1707-1715. |
| [21] | Lin Y H, Zhang F D, Zhang J Q, Ouyang X J, Mo D S, Zhou G Y. Preliminary investigation on temporal and spatial variation of structure of soil fauna community in different natural vegetations of Dinghushan. Acta Ecologica Sinica, 2005, 25(10): 2616-2622. |
| [22] | Liu J L, Li F R, Liu Q J, Niu R X. Relationships between ground beetle community distribution and microhabitats in an arid desert shrubland of the middle Heihe River basin. Acta Ecologica Sinica, 2010, 30(23): 6389-6398. |
| [23] | Moço M K S, Gama-Rodrigues E F, Gama-Rodrigues A C, Machado R C R, Baligar V C. Relationships between invertebrate communities, litter quality and soil attributes under different cacao agroforestry systems in the south of Bahia, Brazil. Applied Soil Ecology, 2010, 46(3): 347-354. |
| [24] | Li Y H, Yang W Q, Luo C D, Wu F Z, Hu J. Dynamics on soil faunal community during the decomposition of mixed eucalypt and alder litters. Acta Ecologica Sinica, 2013, 33(1): 159-167. |
| [25] | Huang X, Wen W Q, Zhang J, Yang W Q, Liu Y, Yan B G, Huang Y M. Soil faunal diversity under typical alpine vegetations in West Sichuan. Chinese Journal of Applied Ecology, 2010, 21(1): 181-190. |
| [26] | Zhu X Y, Gao B J, Yuan S L, Hu Y C. Community structure and seasonal variation of soil arthropods in the forest-steppe ecotone of the mountainous region in Northern Hebei, China. Journal of Mountain Science, 2010, 7(2): 187-196. |
| [27] | Liao C H, Li J X, Yang Y P, Zhang Z C. The community of soil animal in tropical rain forest in Jianfeng Mountain, Hainan Island, China: Relationship between seasonal change of community structure and climatic factors. Acta Ecologica Sinica, 2003, 23(1): 139-147. |
| [28] | Yang X D, Tang J W. Soil arthropod communities in different successional tropical secondary forests in Xishuangbanna, SW China. Chinese Journal of Applied Ecology, 2004, 15(6): 988-994. |
| [29] | Morón-Ríos A, Rodríguez M Á, Pérez-Camacho L, Rebollo S. Effects of seasonal grazing and precipitation regime on the soil macroinvertebrates of a Mediterranean old-field. European Journal of Soil Biology, 2010, 46(2): 91-96. |
| [30] | Zhang H Z, Wu P F, Yang D X, Cui L W, He X J, Xiong Y Q. Dynamics of soil meso-and microfauna communities in Zoigê alpine meadows on the eastern edge of Qinghai-Tibet Plateau, China. Acta Ecologica Sinica, 2011, 31(15): 4385-4397. |
| [31] | Yan S K, Zhang W D, Liu Y X, Fu S L, Li Y L, Wang S L. Impact of heavy snow storm and freezing rain disasters on soil fauna in Chinese fir plantation in southern China. Chinese Journal of Applied Ecology, 2009, 20(1): 65-70. |
| [32] | Xiao N W, Liu X H, Ge F, Ouyang Z Y. Research on soil faunal community composition and structure in the Gaoligong Mountains National Nature Reserve. Acta Ecologica Sinica, 2009, 29(7): 3576-3584. |
| [33] | Salamon J A, Scheu S, Schaefer M. The Collembola community of pure and mixed stands of beech (Fagus sylvatica) and spruce (Picea abies) of different age. Pedobiologia, 2008, 51(5/6): 385-396. |
| [34] | Briones M J I, Ostle N J, McNamara N P, Poskitt J. Functional shifts of grassland soil communities in response to soil warming. Soil Biology and Biochemistry, 2009, 41(2): 315-322. |
| [35] | Qiu J, Fu R S. The effects of temperature and soil water content for the number of Oribatida and Collembola. Journal of Shandong Normal University: Natural Science, 2004, 19(4): 72-74. |
| [36] | DuPont S T, Ferris H, Van Horn M. Effects of cover crop quality and quantity on nematode-based soil food webs and nutrient cycling. Applied Soil Ecology, 2009, 41(2): 157-167. |
| [37] | Wu P F, Liu S R, Liu X L. Composition and spatio-temporal changes of soil macroinvertebrates in the biodiversity hotspot of northern Hengduanshan Mountains, China. Plant and Soil, 2012, 357(1/2): 321-338. |
| [4] | 夏磊, 吴福忠, 杨万勤, 谭波. 川西亚高山森林凋落物分解初期土壤动物对红桦凋落叶质量损失的贡献. 应用生态学报, 2012, 23(2): 301-306. |
| [6] | 高梅香, 张雪萍. 石灰和EM处理条件下土壤动物群落在落叶分解中的变化. 生态学报, 2011, 31(1): 164-174. |
| [10] | 陈鹏, 富德义. 长白山土壤动物在物质循环中作用的初步探讨. 生态学报, 1984, 4(2): 172-180. |
| [11] | 张荣祖, 杨明宪, 陈鹏, 张庭伟. 长白山北坡生态系统土壤动物初步调查. 森林生态系统研究, 1980(1): 133-152. |
| [12] | 殷秀琴, 蒋云峰, 陶岩, 安静超, 辛未冬. 长白山红松阔叶混交林土壤动物生态分布. 地理科学, 2011, 31(8): 935-940. |
| [13] | 佟富春, 金哲东, 王庆礼, 肖以华. 长白山北坡土壤动物群落物种共有度的海拔梯度变化. 应用生态学报, 2003, 14(10): 1723-1728. |
| [14] | 佟富春, 王庆礼, 刘兴双, 肖以华. 长白山次生林演替过程中土壤动物群落的变化. 应用生态学报, 2004, 15(9): 1531-1535. |
| [15] | 陈鹏, 张一. 长白山北坡冰缘环境与土壤动物. 地理科学, 1983, 3(2): 133-140. |
| [16] | 陈鹏, 杨秉庚, 张一. 长白山北坡高山苔原带土壤动物的生态分布. 森林生态系统, 1983, (3): 119-132. |
| [17] | 钱家驹. 长白山北坡各垂直带的典型植被调查报告 // 张子祯. 长白山地理系统研究(第一辑).长春: 东北师范大学出版社, 2010: 89-115. |
| [18] | 刘继亮, 殷秀琴, 邱丽丽. 左家自然保护区大型土壤动物与土壤因子关系研究. 土壤学报, 2008, 45(1): 130-136. |
| [21] | 林英华, 张夫道, 张俊清, 欧阳学军, 莫定生, 周国逸. 鼎湖山不同自然植被土壤动物群落结构时空变化. 生态学报, 2005, 25(10): 2616-2622. |
| [22] | 刘继亮, 李锋瑞, 刘七军, 牛瑞雪. 黑河中游荒漠灌丛斑块地面甲虫群落分布与微生境关系研究. 生态学报, 2010, 30(23): 6389-6398. |
| [24] | 李艳红, 杨万勤, 罗承德, 吴福忠, 胡杰. 桉-桤不同混合比例凋落物分解过程中土壤动物群落动态. 生态学报, 2013, 33(1): 159-167. |
| [25] | 黄旭, 文维全, 张健, 杨万勤, 刘洋, 闫帮国, 黄玉梅. 川西高山典型自然植被土壤动物多样性. 应用生态学报, 2010, 21(1): 181-190. |
| [27] | 廖崇惠, 李健雄, 杨悦屏, 张振才. 海南尖峰岭热带林土壤动物群落—群落结构的季节变化及其气候因素. 生态学报, 2003, 23(1): 139-147. |
| [28] | 杨效东, 唐建维. 西双版纳不同演替状态热带次生林土壤节肢动物群落特征. 应用生态学报, 2004, 15(6): 988-994. |
| [30] | 张洪芝, 吴鹏飞, 杨大星, 崔丽巍, 何先进, 熊远清. 青藏东缘若尔盖高寒草甸中小型土壤动物群落特征及季节变化. 生态学报, 2011, 31(15): 4385-4397. |
| [31] | 颜绍馗, 张伟东, 刘燕新, 傅声雷, 李媛良, 汪思龙. 雨雪冰冻灾害干扰对杉木人工林土壤动物的影响. 应用生态学报, 2009, 20(1): 65-70. |
| [32] | 肖能文, 刘向辉, 戈峰, 欧阳志云. 高黎贡山自然保护区大型土壤动物群落特征. 生态学报, 2009, 29(7): 3576-3584. |
| [35] | 邱军, 傅荣恕. 土壤温湿度对甲螨和跳虫数量的影响. 山东师范大学学报: 自然科学版, 2004, 19(4): 72-74. |