 2014, Vol. 34
2014, Vol. 34文章信息
- 马威, 孙军
- MA Wei, SUN Jun
- 夏、冬季南海北部浮游植物群落特征
- Characteristics of phytoplankton community in the northern South China Sea in summer and winter
- 生态学报, 2014, 34(3): 621-632
- Acta Ecologica Sinica, 2014, 34(3): 621-632
- http://dx.doi.org/10.5846/stxb201203250402
-
文章历史
- 收稿日期:2012-3-25
- 修订日期:2013-10-9






2. 淮河流域水资源保护局, 蚌埠 233000
2. Huaihe River Basin Water Resources Protection Bureau, Bengbu 233000, China
南海北部位处太平洋西部,有台湾岛、海南岛、菲律宾群岛及广东沿岸围绕。该海域生态环境复杂,既有河口、海湾、陆架及陆坡与开阔海,又有各种成因的升降流系统; 既有近岸富营养区域,又有海盆寡营养区域[1]。其通过吕宋海峡等与开阔大洋进行水交换,且有沿岸的多条河流输入大量淡水(如珠江口)[2]。南海北部具有明显的半封闭边缘海的特征,是以热带海洋性气候为主的海域。
从20世纪70年代起,一些研究者对南海北部海域开展了一些调查研究工作,主要以网采浮游植物为主报道[3, 4]。目前对该海域水采浮游植物的定量研究不多,可以进行对比分析的数据资料还较少,而且以前对南海浮游植物的调查研究大多集中在近海区域。Chen等[5]对冬季南海北部上升流区浮游植物进行了研究,认为海水表层营养盐的增加可以导致浮游植物大量增长。Huang等[6]对冬季南海北部的两个暖水团区浮游植物进行了研究,发现导致浮游植物群集的原因不同。乐凤凤等[7]和孙军等[8]分别报道了2004年夏季和冬季南海北部浮游植物群落结构,对浮游植物的分布特征进行了分析,表明硅藻在细胞丰度和物种丰富度上占很高优势,其次为甲藻。Wang等研究发现,吕宋海峡西南部的次表层上升流及混合层的作用会导致浮游植物水华发生[9]。这些研究结果为南海北部浮游植物群落及其季节变化的进一步研究提供了宝贵资料: 在一些水体形成的上升流区域,也会形成浮游植物富集区,从而形成浮游植物高生物量。本文在以前研究者的基础上,综合夏、冬季在南海北部两个航次的调查取样分析,报道该海域浮游植物群落的组成特征,以期为该海域的海洋生态学研究提供基础资料。
1 研究海域与研究方法 1.1 样品采集与处理于2009年7月19日— 8月16日和2010年1月6—30日在南海北部(18°—23.5°N、109°—120°E)进行了两个航次的调查工作,其中夏季共设采样站位64个,冬季采样站位43个(图1),采样水层分别为5、25、50、75、100、125、150、200m(水深不足200m的站位取水底以上2—5m作为底层)。各水层取样1L倒入聚乙烯瓶中,加入中性福尔马林溶液至最终浓度为体积分数2%固定保存。

|
| 图1 夏、冬季调查站位图 Fig. 1 Sampling Stations in survey |
物种分类及统计采用Utermhl方法, 取25mL或100mL浮游植物分样品于Hydro- Bios的Utermhl计数框(100mL分样品需沉降24h后去除上清液),在American- Optical倒置显微镜200或400倍下进行物种鉴定和计数,细胞数量的统计和误差处理参照孙军等[10]。温度、盐度等环境参数来自CTD进行现场测定。营养盐样品取样和分析,参考《海洋调查规范》[11]进行测定。
物种多样性指数采用Shannon- Wiener指数(H′)[12]:
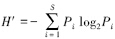
式中,Pi是第i种的个数与该样方总个数之比值; S为样方总数。
Pielou物种均匀度指数Pielou′s index (J)的计算公式[13]:
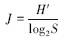
式中,S为样方中物种数。
优势度计算公式如下:
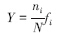
式中,ni为第i个种的个体数;N为全部样品中每种出现的总个体数;fi为该种在各站位出现的频率。
Box-Whisker分析采用浮游植物细胞丰度的对数计算; 浮游植物与环境因子的相关性分析采用SPSS.17.0软件分析;典范对应分析(CCA)应用CANOCO 4.0软件进行,排序结果用物种-环境因子关系的双序图表示。
2 结果与分析 2.1 调查海域水文特征调查区5m层的温度与盐度分布如图2,夏季南海北部5m层海水温度范围在28.3—34℃之间,平均水温达33℃,盐度范围在23.3—30.2之间,平均28.7,温度分布在粤东近岸和海南岛东北部近岸较高; 冬季南海北部海水温度普遍较低,5m层水温范围在16.2—25.8℃之间,平均为22.3℃,盐度范围在32—34.6之间,平均33.8,温度分布则在广东沿岸较高,沿外海方向降低。盐度分布表现出与温度较为一致的趋势。

|
| 图2 5m层温度与盐度分布 Fig. 2 Distribution of temperature and salinity in 5m |
从A断面的角度看(图3),夏季温度和盐度垂向分布变化均匀,层化现象较为明显。上层水体近岸到远岸方向水温升高,水温随深度增加而降低,没有出现明显的跃层现象。而盐度则与温度的分布相反,近岸及上层水体的盐度较高; 冬季温度与盐度垂向分布变化则没有夏季明显,A断面的温度分布在75m层至100m层出现了较为明显的跃层现象。对冬季所有站位的温度与盐度资料分析也能发现,在大约100m层有温度跃层的特点。

|
| 图3 A断面温度与盐度分布 Fig. 3 Distribution of temperature and salinity in A section |
两个调查航次中共鉴定浮游植物物种211种(不包括未定种),分属于硅藻门、甲藻门、金藻门及蓝藻门,及未定类的三裂碎藻(Ebria tripartita)。硅藻门(116种)和甲藻门(90种)物种最多,分别占55.0%和42.6%(表 1)。浮游植物优势物种大多为浮游硅藻类,生态类型主要为暖温带近岸种和浮游广布种。发现夏季主要优势物种为铁氏束毛藻、菱形海线藻、柔弱伪菱形藻及裸甲藻等。束毛藻一般以丝状体存在,本文中对束毛藻以细胞个数计(处理方法:随机计数若干藻丝体细胞数算出平均数后,通过计数每个样品中藻丝体数估算出束毛藻细胞个数)。虽然铁氏束毛藻出现频率不是最高(fi=0.165),但由于其多个细胞主要以藻丝体形式出现,而成为第一优势物种(Y = 0.044); 冬季以菱形海线藻、伏氏海线藻及短刺角毛藻等为主要优势物种。
|
门
Phylum | 季节
Season | 属
Genera | 种
Species | 物种丰度比例/%
Ration of richness |
| 硅藻门Bacillariophyta | 夏季Summer | 50 | 80 | 53.33 |
| 冬季Winter | 40 | 100 | 59.52 | |
| 甲藻门Pyrrophyta | 夏季 | 18 | 65 | 43.33 |
| 冬季 | 14 | 63 | 37.5 | |
| 蓝藻门Cyanophyceae | 夏季 | 2 | 3 | 2 |
| 冬季 | 2 | 3 | 1.79 | |
| 金藻门Chrysophyta | 夏季 | 2 | 2 | 1.33 |
| 冬季 | 2 | 2 | 1.19 |
一些种类出现频率虽不高,但在个别站点数量较高,呈现明显的区域特点,如夏季样品中检测到的柔弱伪菱形藻和束毛藻(主要为铁氏束毛藻); 冬季样品检测中长海毛藻为浮游植物第四优势物种(Y = 0.009),主要在珠江口近岸有较高分布。长海毛藻是典型外洋性物种,常在黑潮中国区分布[15],它的存在说明该海域存在外洋水团入侵。冬季样品中还出现较多舟形藻属中的小舟形藻(Navicula subminuscula),风生流引起的上升流可能促使该底栖物种的悬浮堆积,并成为优势种类。受西南季风或东北季风的影响,导致浮游植物群落组成也随之有明显的冷暖季节差异(表 2)。
|
季节
Season | 物种
Species | 丰度比例/%
Abundance ration | 频度fi Frequency | 优势度Y Dominance |
| *种名更改参见文献[14] | ||||
| 夏季Summer | 铁氏束毛藻Trichodesmium thiebautii | 26.6 | 0.165 | 0.044 |
| 菱形海线藻Thalassionema nitzschioides | 5.6 | 0.345 | 0.019 | |
| 柔弱伪菱形藻*Pseudo-nitzschia delicatissima | 60.6 | 0.030 | 0.018 | |
| 裸甲藻Gymnodinium spp. | 1.0 | 0.476 | 0.005 | |
| 针杆藻Synedra spp. | 0.4 | 0.367 | 0.001 | |
| 冬季Winter | 菱形海线藻Thalassionema nitzschioides | 10.2 | 0.591 | 0.055 |
| 伏氏海线藻*Thalassionema frauenfeldii | 8.6 | 0.377 | 0.030 | |
| 短刺角毛藻Chaetoceros messanensis | 3.9 | 0.355 | 0.013 | |
| 长海毛藻Thalassiothrix longissima | 2.2 | 0.427 | 0.009 | |
| 舟形藻Navicula spp. | 2.3 | 0.386 | 0.008 | |
夏、冬季两个航次调查的浮游植物平均细胞丰度分别为26.49×103个/L和2.69×103个/L,硅藻占细胞丰度的比例最大,平均细胞丰度分别为25.81×103个/L和2.49×103个/L,其次为甲藻。调查结果较历史资料有所不同(表 3),受拖网网目(一般为76μm)影响及拖网时倾角的变化会低估浮游植物细胞丰度,但由于拖网浓缩的海水体积较大,所采集的浮游植物样品中物种丰度也会较高; 一般而言,上层水体所受光照及营养盐等条件较为充足,适宜浮游植物生长,而下层水体由于光照及营养盐等限制,浮游植物细胞丰度可能会较低。此外,受调查季节和调查站位差异的影响,不同调查中浮游植物优势种的构成会有一定的差异。
|
采样时间
Date | 层次/m
Layer | 鉴定物种数
Species number | 平均丰度
Average abundance /(×103个/L) | 采样范围
Sampling areas | 参考文献
References |
| 2009-07 | 0—200 | 150 | 26.49 | 18°—23.5°N,109°—120° | 本文 |
| 2009-12 | 0—200 | 168 | 2.69 | 18°—23.5°N,111°—120° | 本文 |
| 2009-08 | 0—200 | 109 | 8.20 | 18°—22°N,110°—117°E | [16] |
| 2007-08 | 表层 | 122 | 25.21 | 18°—23°N,110°—120° | [17] |
| 2007-08 | 0—200 | 216 | 14.09 | 18°—23°N,110°—120° | [17] |
| 2004-09—10 | 表层 | 152 | 0.11—23.61 | 18°—23.15°N,110°—120° | [18] |
| 2004-08—9 | 0—200 | 162 | 115.00 | 18°—22°N,110°—117°E | [7] |
| 2004-08—9 | 表层 | 112 | 387 | 18°—22°N,110°—117°E | [7] |
| 2004-02—3 | 0—200 | 195 | 206. 40 | 18°—23°N,111°—117° | [8] |
| 1998-012 | 表层 | 85 | 8.46 | 18°—22°N,105°—117°°E | [19] |
| 1998-08 | 表层 | 58 | 181 | 18°—22°N,105°—117°°E | [19] |
| 1998-06—7 | 表层 | 63 | 0.83 | 5°—25°N,105°—120°E | [19] |
| 1985-06 | 0—200 | 152 | 0. 40 | 3°47′—4°02′N,111°58′—112°25′ | [20] |
| 1984 -07 | 0—200 | 177 | 0. 01 | 12°—19°30′N,111°—118° | [21] |
| 1979-06—7 | 0—200 | 287 | 20.00 | 17°—23°N,112°—120°E | [22] |
| 1978-06 | 0—75 | 60 | 0.10 | 12°—15°N,116°—118°E | [23] |
将所调查的站位分为深度浅于200m的沿海大陆架区域和水深大于200m的深水区域进行比较分析(图4)。结果显示,两个季节调查的总浮游植物丰度在水层上的变化趋势较为相似,高细胞丰度主要集中在上层水体,可能与上层水体较强的光照有关; 对于大于200m水深站位,夏季与冬季间表现出了明显的差异,夏季各水层的总细胞丰度变化不大,但冬季随水深增加,总细胞丰度有较为明显的降低趋势; 而对于小于200m水深的站位,夏季浮游植物总细胞丰度随水深表现为先减少后增加,冬季在各水层的总细胞丰度则变化不大。

|
| 图4 调查区浮游植物丰度的Box-Whisker图 Fig. 4 Phytoplankton abundance Box-Whisker plot in survey area |
结合5m层温度及盐度的分布,以及浮游植物水层垂直分布的分析,本文对5m层浮游植物的平面分布进行了分析(图5)。夏、冬季南海北部5m层浮游植物平均细胞丰度分别为149.828 ×103个/L和5.108×103个/L,硅藻的平均细胞丰度分别为141.084×103个/L和4.353 ×103个/L。夏季呈现出由近岸到外海迅速减少的分布,高值区主要出现在广东东部近岸及海南东部近岸; 而冬季主要出现了珠江口近岸和海盆区两个浮游植物细胞丰度高值区。 分析发现: 夏季粤东近岸的高值主要是由于柔弱伪菱形藻在个别站位的大量出现,而海南岛近岸的高值主要是蓝藻类细胞在个别站位出现较多形成; 冬季珠江口近岸及调查海域中南部高值则不是某些单一物种贡献,主要是由多种浮游硅藻的出现造成。

|
| 图5 5m层浮游植物细胞丰度夏季的分布 (a浮游植物,b硅藻; 冬季: c浮游植物,d硅藻) Fig. 5 Distribution in summer of phytoplankton abundence in 5m layer (a phytoplankton,b diatom; winter: c phytoplankton,d diatom) |
对各调查季主要优势物种在5m的分布进行了分析(图6),夏、冬季菱形海线藻均为主要优势物种,平均细胞丰度分别为5.948 ×103个/L和0.559 ×103个/L。菱形海线藻是一种广布性种,分布极为广泛,耐高温和盐度适应能力较强。冬季时浮游植物藻体细胞较夏季明显偏小,可能与温度及营养盐含量较低有关。是浮游植物用以降低营养消耗,达到对特定环境适应性的一种表现。

|
| 图6 调查季主要优势物种在5m层夏季的分布 (a菱形海线藻,b柔弱伪菱形藻,c裸甲藻; 冬季: a菱形海线藻,b伏氏海线藻,c短刺角毛藻) Fig. 6 Distribution in summer of phytoplankton dominant species in 5m layer in the survey seasons (a Thalassionema nitzschioides,b Pseudo-nitzschia delicatissima,c Gymnodinium spp.; winter: d Thalassionema nitzschioides,e Thalassionema frauenfeldii,f Chaetoceros messanensis) |
束毛藻属是一类具有固氮作用的藻类,属外海性暖水种,是海洋中分布最广的远洋性浮游蓝藻[24]。其主要生活在透光率较高、营养盐浓度较低的稳定水团环境中[25]。本文对75m层及以上的样品中束毛藻属进行水柱积分,分析了其在调查区的分布状况(图7)。夏季束毛藻属在大部分调查站位都有出现,平均细胞丰度达29.76×103个/L; 而冬季则只有很少部分站位有束毛藻属的分布,其平均细胞丰度仅为3.51×103个/L; 南海夏季盛行西南季风,高温高盐的黑潮水通过吕宋海峡入侵到南海北部,束毛藻也会随之进入。其分布还会受到强的物理过程(冷空气入侵、海流作用等)的影响[24, 25]。

|
| 图7 75m以浅束毛藻水柱积分细胞丰度的分布 Fig. 7 Distribution of Trichodesmium spp. water coloum cell abundence above 75m layer |
与浮游植物细胞丰度的平面分布相似,浮游植物细胞丰度在A断面也出现了“团块状”分布。夏季在浅水站位和深水站位各出现了一个浮游植物高值区,在浅水站位底层,有浮游植物细胞丰度高于上层的特点; 冬季浮游植物高值,主要分布在3个区域: 近岸区域,A4站位50m以浅区域以及A0与A10间50m以浅区域。这种分布特点主要是由硅藻分布决定的。形成浮游植物这些分布特点的环境因素还有待进一步的讨论(图8)。

|
| 图8 调查季A断面的浮游植物细胞丰度分布 /(×103个/L) Fig. 8 Distribution of phytoplankton cell abundence in A section |
群落物种多样性是群落组织独特的生物学特征,它反映了群落特有的物种组成和个体丰度特征。夏、冬季Shannon-Wiener多样性指数范围分别在0.12—3.77和0.02—4.61之间,平均值分别为2.12和2.36。夏季Shannon-Wiener多样性指数与浮游植物细胞丰度在5m层呈镶嵌的分布,冬季则没有表现出明显的镶嵌分布特征。一般来讲,群落中的单一或某几个种类优势越明显,种类的多样性就会受到影响而降低。冬季主要优势种的优势度高于夏季,5m层平均Shannon-Wiener多样性指数也高于夏季。尽管出现的物种丰度和平均细胞丰度都高于冬季,但种间的数量分布不均匀也会导致多样性指数较低(图9)。

|
| 图9 调查区5m层浮游植物群落多样性指数分布 Fig. 9 Distribution of phytoplankton community diversity indices in 5m in the survey areasseasons |
海水中的无机营养元素供给着浮游植物的生长繁殖,因此营养盐浓度的高低成为浮游植物生长的决定因子[26]。有研究表明南海北部夏季除受到氮营养盐限制外,很可能还受到磷营养盐的限制[26, 27, 28]。本文就所获取的水文资料,主要分析浮游植物与温度、盐度及营养盐间的关系(表 4)。两个航次调查结果显示: 夏、冬季浮游植物细胞丰度均与盐度表现出显著性负相关,冬季浮游植物细胞丰度还与磷酸盐浓度表现出显著性负相关; 整个调查海域磷酸盐浓度较低(很多样本低于检测下限)。
| 夏季Summer | 冬季Winter | ||||||||||
| 温度
Temperature | 盐度
Salinity | NO-2+NO-3 | SiO2-3 | PO3-4 | 温度
Temperature | 盐度
Salinity | NO-2+NO-3 | SiO2-3 | PO3-4 | ||
| * 在 0.05 水平上显著相关; ** 在 0.01 水平上显著相关 | |||||||||||
| 硅藻 | Pearson | 0.083 | -0.27* * | -0.056 | -0.068 | -0.138 | -0.034 | -0.419* * | -0.142* | -0.116 | -0.30* * |
| Diatom | N | 266 | 266 | 230 | 252 | 187 | 218 | 218 | 210 | 198 | 131 |
| 甲藻 | Pearson | 0.147* | -0.172* * | -0.150* | -0.174* * | -0.142 | 0.053 | -0.086 | -0.190* * | -0.099 | 0.020 |
| Dinoflagellate | N | 266 | 266 | 230 | 252 | 187 | 218 | 218 | 210 | 198 | 131 |
| 浮游植物 | Pearson | 0.088 | -0.260* * | -0.071 | -0.081 | -0.093 | -0.032 | -0.417* * | -0.149* | -0.118 | -0.296* * |
| Phytoplankton | N | 266 | 266 | 230 | 252 | 187 | 218 | 218 | 210 | 198 | 131 |
进一步对南海北部调查区浮游植物常见物种(按照优势度排序,表 2)与环境因子进行典范对应分析(CCA)(图10,图11)发现,夏季整个海区的浮游植物常见种主要受到亚硝酸盐的影响,而总氮和磷酸盐只在一定程度上对广布的伪菱形藻和舟形藻起一定作用(图10),这可能从一个侧面反映了夏季南海北部细菌的生长对海区浮游植物的作用亦相当明显; 冬季情况则有所不同,没有一个明显作用的环境因子,温度是影响整个海区浮游植物组成和分布的主要环境因子,但是其作用不甚明显,磷酸盐只在一定程度上影响海区常见物种菱形海线藻及暖水代表性物种短刺角毛藻。

|
|
图10
夏季水采浮游植物主要优势物种与环境因子的双序图
Fig. 10
Biplot of dominant phytoplankton species and environment factors in summer water-samples
S:盐度; T:温度; PO4:磷酸盐; SiO3:硅酸盐; NO2:亚硝酸盐; TN: 硝酸盐与亚硝酸盐总和; T.thi:铁氏束毛藻; Pro:具齿原甲藻; Gym:裸甲藻; Syn:针杆藻; Nit:伪菱形藻; Nav:舟形藻; Tha1:菱形海线藻; Tha2:伏氏海线藻; Ple:曲舟藻; Pse:柔弱伪菱形藻
S: salinity; T: temperature; PO4: phosphate; SiO3: silicate; NO2:nitrite; NO3: nitrate; TN: nitrite + nitrate; T.thi: Trichodesmium thiebaultii; Pro: Prorocentrum dentatum; Gym: Gyminodinium spp.; Syn: Synedria spp.; Nit: Pseudo-nitzchia. spp.; Nav: Navicula spp.; Tha1: Thalassionema nitzschioides; Tha2: Thalassionema frauenfeldii; Ple: Pleurosigma spp.; Pse: Pseudo-nitzschia delicatissima |

|
|
图11
冬季水采浮游植物主要优势物种与环境因子的双序图
Fig. 11
Biplot of dominant phytoplankton species and environment factors in winter water-samples
S:盐度; T:温度; PO4:磷酸盐; SiO3:硅酸盐; NO3:硝酸盐; TN: 硝酸盐与亚硝酸盐总和; T.thi:铁氏束毛藻; T.ery::红海束毛藻; Rhi:笔尖形根管藻; Tha1:菱形海线藻; Tha2:伏氏海线藻; Tha3:长海毛藻; Cha:短刺角毛藻; Cos:圆筛藻; Ple:曲舟藻; Nav:舟形藻
S: salinity; T: temperature; PO4: phosphate; SiO3: silicate; NO3: nitrate; TN: nitrite + nitrate; T.thi: Trichodesmium thiebaultii; T.ery: Trichodesmium erythraeum; Rhi: Rhizosolenia styliformis; Tha1: Thalassionema nitzschioides; Tha2: Thalassionema frauenfeldii; Tha3: Thalassiothrix longissima; Cha: Chaetoceros messanensis; Cos: Coscinodiscus spp.; Ple: Pleurosigma spp.; Nav: Navicula spp. |
但是需要指明的是,南海北部水文动力条件复杂,其东北部常年受黑潮暖流影响,近岸受到广东沿岸流和海南岛沿岸流交互作用,对该海域浮游植物群落特征动态变化与环境因子之间的关系还需要长期细致的调查,目前还很难有一个较明晰的主线。
3 结论夏、冬季对南海北部进行的两个航次调查南海北部共鉴定浮游植物4门85属211种,硅藻类占总物种丰度的55.0%。夏季浮游植物平均细胞丰度明显高于冬季。夏季细胞丰度高值区主要集中在广东东部近岸和海南东部近岸海域; 冬季细胞丰度高值区主要出现在珠江口近岸和中央海盆区海域。夏季出现的主要优势种属有: 铁氏束毛藻、藻菱形海线藻、柔弱伪菱形藻及裸甲藻等; 冬季出现的主要优势物种有: 菱形海线藻、伏氏海线藻、短刺角毛藻等。夏季较冬季相比,束毛藻属的分布范围广且细胞丰度高。
南海北部浮游植物的群落特征复杂,对于浮游植物的物种组成及数量分布与环境因子之间关系的认识,还需要更多的浮游植物相关数据资料积累。
致谢: 厦门大学戴民汉教授提供相关营养盐数据,厦门大学胡建宇教授提供温度和盐度资料,宋书群、栾青杉、郭术津、田伟采集水样,特此致谢。
| [1] | Gong G C, Liu K K, Liu C T, Pai S C. The chemical hydrography of the South China Sea west of Luzon and a comparison with the West Philippine Sea. Terrestrial, Atmospheric and Oceanic Sciences, 1992, 3: 587-602. |
| [2] | Lee M Y, Wei K Y, Chen Y G. High resolution oxygen isotope stratigraphy for the last 150000 years in the southern South China Sea: Core MD972151. Terrestrial, Atmospheric and Oceanic Sciences, 1999, 10: 239-254. |
| [3] | Guo Y J, Ye J S, Zhou H Q. Quantitative distribution of phytoplankton in Xisha & Zhongsha Islands//Marine Biological Survey Research Reports in Xisha & Zhongsha Islands. Beijing: Science Press, 1978: 1-10. |
| [4] | Ye J S, Lin Y S, Yuan W B. Quantitative distribution of phytoplankton in the East Sand Island sea area in summer // South China Sea marine biological research papers(Ⅰ). Beijing: Ocean Press, 1983: 1-6. |
| [5] | Chen C C, Kwo S F, Chung S W. Liu K K. Winter phytoplankton blooms in the shallow mixed layer of the South China Sea enhanced by upwelling. Journal of Marine Systems, 2006, 59: 97-110. |
| [6] | Huang B Q, Hu J, Xu H Z, Cao Z R, Wang D X. Phytoplankton community at warm eddies in the northern South China Sea in winter 2003/2004. Deep-Sea Research II, 2010, 57: 1792-1798. |
| [7] | Le F F, Sun J, Ning X R, Song S Q, Cai Y M, Liu C G. Phytoplankton in the northern South China Sea in summer 2004. Oceanologia et Limnologia Sinica, 2006, 37(3): 238-248. |
| [8] | Sun J, Song S Q, Le F F, Wang D, Dai M H, Ning X R. Phytoplankton in the northern South China Sea in winter 2004. Acta oceanologica cinica, 2007, 29(5): 132-145. |
| [9] | Wang J J, Tang D L, Sui Y. Winter phytoplankton bloom induced by subsurface upwelling and mixed layer entrainment southwest of Luzon Strait. Journal of Marine Systems, 2010, 83(3/4): 141-149. |
| [10] | Sun J, Liu D Y, Qian S B. A quantitative research and analysis method formarine phytoplankton: An introduction to Utermöhl method and its modification. Journal of Oceanography of Huanghai & Bohai Seas, 2002, 20(2): 105-112. |
| [11] | State Bureau of Technical Supervise. The Specification for Oceanographic Survey-marine Biological Survey (GB / T12736. 6-2007). Beijing: China Standard Press, 2007: 30-35. |
| [12] | Shannon C E, Weaver W. The Mathematical Theory of Communication. Urbana IL: University of Illinois Press, 1949: 1-125. |
| [13] | Pielou E C. An Introduction to Mathmatical Ecology. New York: Wiley-Interscience, 1969: 1-28. |
| [14] | Sun J, Liu D Y. The preliminary notion on nomenclature of common phytoplankton in China Sea waters. Oceanologia et Limnologia Sinica, 2002, 33(3): 271-286. |
| [15] | Sun J, Liu D Y, Qian S B. Ryukyu Islands and adjacent waters of planktonic diatoms and the relationship between water masses in the summer of 1997 // China Oceanography Papers II, Beijing: Ocean Press, 2000, 12: 158-169. |
| [16] | Li X, Sun J, Tian W, Wang M. Phytoplankton in the northern South China Sea in summer 2009. Marine Sciences,2012,10:33-39. |
| [17] | Ke Z X, Huang L M, Tan Y H, Yin J Q. Species composition and abundance of phytoplankton in the northern South China Sea in summer 2007. Journal of Tropical Oceanography, 2011, 30(1): 131-143. |
| [18] | Li T, Liu S, Wang G F, Cao W X, Huang L M, Lin Q Y. Species composition of phytoplankton and its distribution in the northern South China Sea in autumn 2004. Journal of Tropical Oceanography, 2010, 29(2): 65-73. |
| [19] | Zhu G H, Ning X R, Cai Y M, Liu Z L, Liu C G. Studies on species composition and abundance distribution of phytoplankton in the South China Sea. Acta Oceanologica Sinica, 2003, 25(sup 2): 8-23. |
| [20] | South China Sea Institute of oceanology chinese academy of sciences. James Shoal-a comprehensive survey of southern China Report. Beijing: Ocean Press, 1987: 106-131. |
| [21] | South China Sea Institute of oceanology chinese academy of sciences. a comprehensive survey of environmental resources in the central of southern China Report. Beijing: Ocean Press, 1988: 162-231. |
| [22] | South China Sea Institute of oceanology chinese academy of sciences. Comprehensive survey conducted in South China Sea area. Beijing: Ocean Press, 1985: 332-357. |
| [23] | Guo Y J, Ye J S. Horizontal and vertical distribution of phytoplankton in the central of South China Sea//Comprehensive survey conducted in South China Sea area (Ⅰ). Beijing: Science Press, 1982: 217-230. |
| [24] | Capone D G, Zehr J P, Paerl H W, Bergman B, Carpenter E J. Trichodesmium, A globally significant marine cyanobacterium. Science, 1997, 276 (53l6): 1221-1229. |
| [25] | Chang J, Chiang K P, Gong G C. Seasonal variation and cross-shelf distribution of the nitrogen-fixing cyanobacterium, Trichodesmium, in southern East China Sea. Continental Shelf Research, 2000, 20: 479-492. |
| [26] | Xu J. Nutrient limitation in the Pearl River Estuary, Hong Kong waters and adjacent South China Sea waters. Hong Kong: Hong Kong Univ. Sci. Technol., 2007. |
| [27] | Chen Lee Y L, Chen H Y. Seasonal dynamics of primary and new production in the northern South China Sea: the significance of river discharge and nutrient advection. Deep-Sea Res. I, 2006, 53: 971-986. |
| [28] | Yin K D, Qian P Y, Wu M C.S., Chen J C, Huang L M, Song X Y, Jian W J. Shift from P to N limitation of phytoplankt on biomass across the Pearl River estuarine plume during summer. Mar. Ecol. Progr. Ser., 2001, 221: 17-28. |
| [3] | 郭玉洁, 叶嘉松, 周汉秋. 西沙、中沙群岛周围海域浮游植物的数量分布. 我国西沙、中沙群岛海域海洋生物调查研究报告集. 北京: 科学出版社, 1978: 1-10. |
| [4] | 叶嘉松, 林永水, 袁文彬. 东沙群岛周围海域夏季浮游植物的数量分布. 南海海洋生物研究论文集(一). 北京:海洋出版社, 1983: 1-6. |
| [7] | 乐凤凤, 孙军, 宁修仁, 宋书群, 蔡昱明, 刘诚刚. 2004年夏季中国南海北部的浮游植物. 海洋与湖沼, 2006, 37(3): 238-248. |
| [8] | 孙军, 宋书群, 乐凤凤, 王丹, 戴明汉. 2004年冬季南海北部浮游植物. 海洋学报, 2007, 29(5): 132-145. |
| [10] | 孙军, 刘东艳, 钱树本. 一种海洋浮游植物定量研究分析方法——Utermhl方法的介绍及其改进. 黄渤海海洋, 2002, 20(2): 105-112. |
| [11] | 国家技术监督局.《海洋调查规范》(GB/T12736. 6—2007). 北京: 中国标准出版社, 2007: 30-35. |
| [14] | 孙军, 刘东艳. 中国海区常见浮游植物种名更改初步意见. 海洋与湖沼, 2002, 33(3): 271-286. |
| [15] | 孙军, 刘东艳, 钱树本. 1997年夏硫球群岛及邻近水域浮游硅藻与水团的相互关系. 中国海洋学文集Ⅱ, 北京: 海洋出版社, 2000, 12: 158-169. |
| [16] | 李欣, 孙军, 田伟,汪岷. 2009年夏季南海北部的浮游植物群落. 海洋科学, 2012,10:33-39. |
| [17] | 柯志新, 黄良民, 谭烨辉, 尹健强. 2007年夏季南海北部浮游植物的物种组成及丰度分布. 热带海洋学报, 2011, 30(1): 131-143. |
| [18] | 李涛, 刘胜, 王桂芬, 曹文熙, 黄良民, 林秋艳. 2004年秋季南海北部浮游植物组成及其数量分布特征. 热带海洋学报, 2010, 29(2): 65-73. |
| [19] | 朱根海, 宁修仁, 蔡昱明, 刘子琳, 刘诚刚. 南海浮游植物物种组成和丰度分布的研究. 海洋学报, 2003, 25(增刊2): 8-23. |
| [20] | 中国科学院南海海洋研究所. 曾母暗沙-中国南疆综合调查研究报告. 北京: 科学出版社, 1987: 106-131. |
| [21] | 中国科学院南海海洋研究所. 南海中部海域环境资源综合调查报告. 北京: 科学出版社, 1988: 162-231. |
| [22] | 中国科学院南海海洋研究所. 南海海区综合调查研究报告. 北京: 科学出版社, 1985: 332-357. |
| [23] | 郭玉洁, 叶嘉松. 南海中部海域浮游植物的水平和垂直分布. 南海海区综合调查研究报告(一). 北京: 科学出版社, 1982: 217-230. |