 2014, Vol. 34
2014, Vol. 34文章信息
- 张富华, 胡聃, 孙凡, 郭振, 李元征, 王晓琳, 马生丽
- ZHANG Fuhua, HU Dan, SUN Fan, GUO Zhen, LI Yuanzheng, WANG Xiaolin, MA Shengli
- 北京城区夏、秋季节短期增温对月季暗呼吸及光合参数的影响
- Ecological effects of short-time increased air temperature in Summer and Autumn periods on dark respirations and typical photosynthetic characteristics for Rosa chinensis in Beijing urban areas
- 生态学报, 2014, 34(24): 7385-7392
- Acta Ecologica Sinica, 2014, 34(24): 7385-7392
- http://dx.doi.org/10.5846/stxb201303180445
-
文章历史
- 收稿日期:2013-03-18
- 网络出版日期:2014-03-19






2. 中国科学院生态环境研究中心, 城市与区域生态国家重点实验室, 北京 100085
2. State Key Laboratory of Urban and Regional Ecology, Research Center for Eco-Environmental Sciences, Chinese Academy of Sciences, Beijing 100085, China
温度是植物生长、发育的重要生态因子,对植物叶片呼吸有重要影响[1]。随着城市的发展,城市形态改变、人工设施影响加大、大气污染以及人工废热的排放,导致城市气温上升,表现城区气温显著高于郊区气温[2]。研究表明,到21世纪末全球地表温度上升大约1.1—6.4℃[3],温度上升对植物生理过程、种群、群落、生态系统有显著影响[4]。
叶片暗呼吸是植物新陈代谢重要的生理过程,它能返回大气光合固碳量约为40%—50%[5]。本研究考虑两种暗呼吸类型:白天叶片在光照下暗呼吸为RL;白天叶片在黑暗下暗呼吸为RD,这两种暗呼吸与通常人们研究很多的叶片夜间暗呼吸不同[6, 7]。在光照与黑暗下暗呼吸都会发生,大部分研究报道叶片RL低于RD,这表明在光照下叶片暗呼吸存在部分光抑制现象[8, 9, 10, 11, 12, 13, 14]。光抑制暗呼吸速率范围为16%—77%[10],其过程是快速的(大约50s),发生的最低光强为3μmol m-2 s-1[11]。目前,光抑制暗呼吸机理还不清楚[15],但研究表明光抑制呼吸程度似乎依赖于植物光合能力[9]。光对叶片暗呼吸的抑制效应,对未来寻求精确估算生态系统呼吸碳损失非常重要[7]。
目前,国内外研究温度变化对植物呼吸影响是 一个热点问题,但光对植物呼吸抑制效应研究相对 极少。国外通常用生长箱模拟植物生长环境,通过幼苗盆栽实验,观测温度改变对植物呼吸的影响。就城市而言,人工模拟实验无法反映城市开放环境下实际生态因子变化,以及植物对城市环境变化的长期适应性[16]。Owen等人[10]利用生长箱研究发现常绿树种疏花桉幼苗(Eucalyptus pauciflora Sieb. ex Spreng),在光强0—100 μmol m-2 s-1范围内叶片暗呼吸随温度升高而增加,光抑制呼吸随温度升高而降低。Shapiro等人[7]利用生长箱发现苍耳幼苗(Xanthium strumarium L.)叶片暗呼吸随温度(23、28、33℃)升高而增加,并与温度有很强正相关性。因此,采用生态因子梯度法,通过现场模拟对比试验,已成为当今国内外研究生态因子与植物生理生态关系重要的技术途径[17]。
本研究以北京地区绿地中优势植物[18]月季(Rosa chinensis)为实验材料,用Li-6400(Li-Cor,USA)光合仪模拟夏、秋季短期增温对月季光合参数、暗呼吸及光抑制呼吸的影响。本文旨在阐明月季暗呼吸及光抑制呼吸对城市气温上升短时响应能力,分析光抑制呼吸变化对植物碳平衡的影响,为今后研究城市气温上升对植物呼吸长期影响,发掘植物碳源-碳汇调节能力、改善城市植物生态功能技术途径提供前期铺垫。
1 材料与方法 1.1 实验材料2012年8月与9月上旬,选取长势一致成龄月季(Rosa chinensis),在北京城区居住绿地内进行增温实验。月季(Rosa chinensis)是蔷薇科蔷薇属常绿或半常绿低矮灌木,选取的月季呈块状种植,种植时间为2005年,种植面积大约为30m2,生长状况基本一致,平均高度约为0.89m。
1.2 实验地点本研究在北京市海淀区柏儒苑小区中心绿地内,属于北京市四、五环之间典型居民小区。小区内绿化面积约为35%,样地附近无高大建筑物遮挡,月季周围也无乔木遮荫。该研究区属典型温带湿润季风气候区,冬季寒冷干燥,夏季高温多雨,春秋干旱;年均气温12.5℃,极端最低气温为-21.7℃,最高气温41.6℃ 。实验观测点土壤条件见表 1。
| 土壤 Soil | pH | 全NTotal nitrogen/(g/kg) | 全CTotal carbon/(g/kg) | C/N |
| 数值为平均值(括号内为标准差)(n=3) | ||||
| 数值 Value | 8.48(0.153) | 0.104(0.004) | 1.857(0.044) | 17.83(1.055) |
| 测定方法Measurement method | 电位法 | 元素分析仪 | 元素分析仪 | 元素分析仪 |
选取2012年夏季8月与秋季9月上午(9:00—11:00)晴朗天气,采用美国Li-Cor 公司生产的Li-6400便携式光合仪测定系统现场观测。1961—2000年近40年北京市市区夏季平均气温为25.02℃[19],用Li-6400(Li- Cor,USA)光合仪测得夏季气温为28℃,秋季气温为20℃ 。在此基础上用光合仪控温模块短期增温处理:(1)夏季时,白天增温2℃ (28—30℃)(9:00—11:00);(2)白天增温5℃ (28—33℃)(9:00—11:00);(3)秋季时,白天增温3℃ (20—23℃)(9:00—11:00);(4)白天增温5℃ (20—25℃)(9:00—11:00)。每次增温处理至少重复3枚叶片,增温处理后叶片最多适应20min,以诱导气孔展开,增温幅度和处理方法参照苍耳(Xanthium strumarium L)[7]和藏北紫花针茅(Stipa purpurea)[20]的实验方法。
1.3.2 光合参数测量每种增温环境选取3—6株成熟月季,随机选择植株中上部朝阳方向一致的3—5枚成熟叶片进行测定。净光合速率Anet测定:每种增温环境稳定之后,用Li-6400光合仪红蓝光源控制低光强响应曲线(80、70、60、50、40、30、20、10、0、0、0 μmol m-2 s-1),并测量其净光合速率Anet。测定的参数有:净光合速率Anet、气孔导度Gs、胞间CO2浓度Ci 、蒸腾速率Tr 。使用开放气路,设定空气流速为500μmol/s,大气压约100kpa,由Li-6400光合仪配套的CO2注入系统控制CO2浓度,控制现场CO2为380 μmol m-2 s-1,相对湿度在30%—40%之间。
1.3.3 暗呼吸测定实验测定两种暗呼吸:(1)白天月季叶片有光暗呼吸RL,通过Li-6400光合仪测得低光强响应曲线的净光合速率Anet,用线性回归外推估算得到,每种增温环境至少重复3次取其平均值;(2)白天月季叶片无光暗呼吸RD,由光合仪红蓝光源控制光强PAR=0,0,0测得CO2排放速率,3次重复取其平均值[6, 7],如图 1。在实验中,RL/ RD比率反应出白天光抑制叶片暗呼吸的程度,该指标是一个随生态因子变化的变量。如果RL/ RD比率越高,则光抑制暗呼吸程度越小;反之,则光抑制呼吸越高。光抑制暗呼吸表达式为:
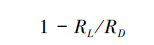
式中,RL为白天叶片有光暗呼吸;RD白天叶片无光暗呼吸[6]。
通常呼吸对温度敏感性用温度系数Q10来表达,但目前国内外还没有报道过用Q10来表达暗呼吸(RL和RD)对温度敏感性。因此,本研究把叶片暗呼吸对温度升高的敏感性表达为:
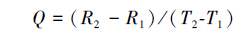
式中,Q为每增温1℃ 暗呼吸相对变化值,T2为较高温度值,T1较低温度值,R2为T2温度对应暗呼吸值,R1为T1温度对应暗呼吸值。

|
| 图 1 月季光合响应曲线 Fig. 1 Photosynthesis- light response curve of Rosa chinensis under low light intensity 当光强PAR=0时,被认为叶片无光暗呼吸(RD); 光响应曲线上半部分延伸到y轴,截距是叶片有光暗呼吸(RL) |
基于SPSS18.0对实验数据统计分析,采用单因素方差分析(One-Way ANOVA),分析夏、秋短期增温处理对月季叶片光合参数、暗呼吸及光抑制呼吸的影响,用LSD多重比较法检验夏、秋增温对不同叶片呼吸差异性。利用Origin8软件绘图。
2 结果与分析 2.1 夏、秋季增温对月季光合参数的影响如图 2所示,秋季时,增温3℃ 对气孔导度(Gs)影响不显著(P>0.05),而增温5℃ 对Gs有显著影响(P<0.05);夏季时,增温2℃ 与5℃ 对Gs影响显著(P<0.05)。秋季增温25℃ 时,Gs达到最大值(0.088±0.001)mol m-2 s-1,33℃ 时为最小值(0.049±0.003) μmol m-2 s-1。总体上看,Gs随温度上升先增加后减少,Gs对温度变化表现非常敏感。

|
| 图 2 不同季节短期增温月季气孔导度 (Gs) 、胞间CO2浓度 (Ci) 和蒸腾速率 (Tr) 的变化 Fig. 2 The variations of Gs,Ci and Tr in the leaves of Rosa chinensis treated with USATR in different seasons 数据为平均数±标准误差,不同字母表示差异达5%显著水平 |
秋季时,增温3℃ 与5℃ 对胞间CO2 浓度(Ci)影响不显著(P>0.05);夏季时,增温2℃ 与5℃ 也对Ci影响不显著(P>0.05)(图 2)。表明Ci对温度上升响应不敏感。
秋季时,增温3℃ 与5℃ 对蒸腾速率(Tr)有显著影响(P<0.05);夏季时,增温2℃ 与5℃ 对Tr影响显著(P<0.05)(图 2)。Tr随温度的上升而增加,增温到33℃ 时,Tr达到最大值(1.844±0.163)mmol m-2 s-1,20℃ 为最小值(0.961±0.069) μmol m-2 s-1。表明温度是影响月季蒸腾速率的一个重要的生态因子,温度升高加速叶片蒸腾速率,特别是局部城市高温天气,可能会导致植物严重缺水。其他学者研究也发现,温度上升对灌木夏蜡梅(Sinocalycanthus chinensis)[21]与紫花雪山报春(Primula sinopurpurea Balf.f.ex Hutch)[1]的蒸腾速率有显著影响。
2.2 夏、秋季增温对月季暗呼吸的影响图 3表明,秋季增温5℃,有光暗呼吸(RL)显著高于增温3℃ 的值(P<0.05),而增温3℃ 对RL影响不显著(P>0.05);夏季增温5℃,RL显著高于增温2℃ (P<0.05),增温2℃ 对RL没有显著影响(P>0.05)。表明RL与温度有很强的正相关性,温度是影响月季RL一个重要的生态因子。

|
| 图 3 不同季节短期增温月季有光暗呼吸RL、无光暗呼吸RD及光抑制呼吸变化 Fig. 3 Changes in RL,RD and light-inhibited leaf R of Rosa chinensis under USATR in different seasons 数据为平均数±标准误差,不同字母表示差异达5%显著水平 |
秋季增温5℃,无光暗呼吸(RD)显著高于增温3℃ 的值(P<0.05),它们都对RD影响显著(P<0.05);夏季增温5℃,无光暗呼吸(RD)也显著高于增温2℃ (P<0.05)(图 3)。夏、秋季增温时,RD都随温度上升而加强,但夏季增温效应高于秋季增温,可能是因为夏季温度普遍高于秋季温度。RD增温到33℃ 时达到最大值(1.935±0.016) μmol m-2 s-1,20℃ 时为最小值(1.168±0.053) μmol m-2 s-1。这表明RD对温度升高响应非常敏感,温度改变2—3℃ 都会对RD造成很大的变化。
2.3 夏、秋季增温对月季光抑制呼吸的影响光抑制呼吸随温度上升而减弱。秋季增温3℃,光抑制呼吸显著高于增温5℃ (P<0.05);夏季时,增温2℃ 与5℃ 没有显著差异(P>0.05)(图 3)。光抑制呼吸比率在20℃ 时达到最大值(47.3±6.731)%,增温到33℃ 时为最小值(19.9±6.897)%。温度20℃ 时,光抑制呼吸的值是33℃ 的2.37倍,这表明低温时光抑制呼吸就会加剧,从而导致植物暗呼吸减少。可见城市气温升高会导致光抑制呼吸降低,增加叶片的暗呼吸,减少单个植株固碳量,导致植物碳汇功能减弱。
2.4 月季暗呼吸对夏、秋增温敏感性的影响由表 2可知,RL对增温敏感性,用增温区间内每增温1℃,RL相对变化值(QL)来表示。秋季增温5℃,QL明显高于增温3℃ 的值,并且有显著差异(P<0.05);夏季时,增温2℃ 与5℃,QL没有显著差 异(P>0.05)。秋季增温5℃,QL显著高于其它3个增温区间(P<0.05)。可以看出秋季增温5℃,RL对增温敏感性高,而RL对夏季增温敏感性低。
RD对增温敏感性,用增温区间内每增温1℃,RD相对变化值(QD)来表示。秋季增温5℃,QD显著高于增温3℃ (P<0.05);夏季时,增温2℃ 与5℃,QD没有显著差异(P>0.05)(表 2)。
| 增温处理/℃USATR treatment | 单位增温有光暗呼吸变化值(QL)Rate of variation of respiration per unit USATR of 1℃ /(μmol m-2 s-1 ℃-1) | 单位增温无光暗呼吸变化值 (QD)Rate of variation of respiration per unit USATR of 1℃ /(μmol m-2 s-1 ℃-1) | 单位增温光抑制呼吸变化值(Q(1-RL/RD))Rate of variation of respiration per unit for light-inhibited/(%/℃) | |
| 数据为平均数±标准误差,不同字母表示差异达5%显著水平; QL为每增温1℃ 有光暗呼吸(RL)相对变化值,QD为每增温1℃ 无光暗呼吸(RD)相对变化值,Q(1-RL/RD)为每增温1℃ 光抑制呼吸率相对变化值; USATR: urban short-time air temperature raising | ||||
| 秋季增温 | 20—23 | 0.033±0.015 a | 0.025±0.011 a | 1.016±0.571 a |
| USATR in Autumn | 20—25 | 0.089±0.011 b | 0.049±0.016 b | 4.488±0.251 b |
| 夏季增温 | 28—30 | 0.045±0.029 a | 0.031±0.013 ab | 2.865±0.672 c |
| USATR in Summer | 28—33 | 0.058±0.032 ab | 0.049±0.007 b | 1.448±0.813 a |
月季光抑制呼吸对增温敏感性(Q(1-RL/RD))见表 2。秋季增温5℃, Q(1-RL/RD)显著高于增温3℃ (P<0.05);夏季增温2℃,(Q(1-RL/RD))显著高于增温5℃ (P<0.05)。表明秋季增温5℃ 与夏季增温2℃,月季光抑制呼吸对增温敏感性高。
3 讨论由于城市化迅速发展、人工地表面积扩大和交通、建筑排热等因素的影响,导致城市热环境改变,气温逐年升高。1961—2000年北京市市区与郊区白天气温温差年平均值为3.3℃,最高达到4.4℃[19]。本研究中,夏、秋增温处理提高了月季气孔导度(Gs)、蒸腾速率(Tr),而对胞间CO2 浓度(Ci)影响不显著(P>0.05)(图 2)。
光抑制呼吸已被很多实验研究所证实,并与植物光合过程有关[22]。温度升高后,叶片RL增加,暗示着对于ATP和NADPH的潜在需要,这个潜在需要决定着光抑制呼吸的程度[6]。实验中,光抑制呼吸随温度上升而降低,秋季增温处理对光抑制呼吸有显著影响(P<0.05)(图 3)。如果温度升高,光抑制呼吸降低,就会导致植物暗呼吸增加,进而会减弱植物的碳汇能力。
从目前研究趋势看,国际上大部分采用生长箱模拟植物生长环境,观测温度变化对植物幼苗呼吸的影响[7, 10]。一般会控制3个倍增温度,这样可以短时间内,对比不同温度对植物暗呼吸影响,弱化了植物对环境适应的生理变化。目前国内外用温度系数Q10来描述植物呼吸对温度敏感性变化,由于本文研究气温变化幅度相对较小,没有足够样本数据拟合呼吸曲线,因而采用每单位增温引起呼吸变化值(QL和QD),来描述呼吸对增温敏感性变化。
本实验在城市开放绿地条件进行,城市生态过程在时空变异上是一种相对小尺度现象。用Li-6400光合仪控制影响植物呼吸主要生态因子(光照、温度、CO2等),来模拟夏、秋季短期增温处理对成年月季暗呼吸的影响。这样既能反映城区实际生态因子小尺度变化特征,也能反映城市植物对生态因子变化的短期响应,这是进一步研究长期、累积性变化的基础。但值得注意的是,这种短期温度效应,不能完全反应长期温度变化对植物暗呼吸的影响,需结合现场长期生态观测来了解植物暗呼吸变化特征[23]。
4 结论温度作为重要的生态因子之一,对植物光合参数及暗呼吸影响非常显著。夏、秋短期增温处理均显著提高了月季暗呼吸,不利于植物有机物的积累,气温上升导致植物叶片碳汇损失,应该在城市碳循环与绿地碳收支模型中有所反映。有关光抑制呼吸及其生态响应机理有待进一步深入研究,这也是进一步探讨城市热岛对城市植物生理生态影响以及构建城市碳平衡模型的基础。
| [1] | Shu W H, Zhang G F. The effect of soil and air temperature on photosynthesis and transpiration of Primula inopurpurea. Acta Botanica Boreali-Occidentalia Sinica, 2002, 22(4): 824-830. |
| [2] | Oke T R. City size and the urban heat island. Atmospheric Environment, 1973, 7(8): 769-779. |
| [3] | IPCC (Intergovernmental Panel on Climate Change). Climate Change 2007: Synthesis Report. Summary for Policymakers. [2012-1-20]. http://www.ipcc.ch. |
| [4] | Usami T, Lee J, Oikawa T. Interactive effects of increased temperature and CO2 on the growth of Quercus myrsinaefolia saplings. Plant, Cell and Environment, 2001, 24(10): 1007-1019. |
| [5] | Amthor J S. Terrestrial higher-plant response to increasing atmosphere [CO2] in relation to the global carbon cycle. Global Change Blology, 1995, 1(4): 243-274. |
| [6] | Wang X Z, James D L, David T T, Jeffrey R S, Kevin L G. Effects of elevated atmospheric CO2 concentration on leaf dark respiration of Xanthium strumarium in light and in darkness. Proceedings of the National Academy of Sciences of the United States of America, 2001, 98(5): 2479-2484. |
| [7] | Shapiro J B, Griffin K L, Lewisand J D, Tissue D T. Response of Xanthium strumarium leaf respiration in the light to elevated CO2 concentration, nitrogen availability and temperature. New Phytologist, 2004, 162(2): 377-386. |
| [8] | Brooks A, Farquhar G D. Effect of temperature on the CO2/O2 specificity ofribulose-1,5-bisphosphate carboxylase/oxygenase and the rate of respiration in the light. Planta, 1985, 165(3): 397-406. |
| [9] | Kromer S. Respiration during photosynthesis. Annual Review of Plant Physiology and Plant MoleCular Biology, 1995, 46: 45-70. |
| [10] | Atkin O K, Evans J R, Ball M C, Lambers H, Pons T L. Leaf respiration of snow gum in the light and dark, interactions between temperature and irradiance. Plant Physiology, 2000, 122(3): 915-923. |
| [11] | Atkin O K, Evans J R, Ball M C, Siebke K, Pons T L, Lambers H. Light inhibition of leaf respiration: the role of irradiance and temperature // Gardestrom P, Gliminius K, Glaser E, eds. Plant Mitochondria: From Gene to Function. Leiden: Bluckhuys Publishers, 1998: 567-574. |
| [12] | Atkin O K, Evans J R, Siebke K. Relationship between the inhibition of leaf respiration by light and enhancement of leaf dark respiration following light treatment. Plant Physiology, 1998, 25(4): 437-443. |
| [13] | Sharp R E, Matthews M A, Boyer J S. Kok effect and the quantum yield of photosynthesis: light partlally inhibits dark respiration. Plant Physiology, 1984, 75(1): 95-101. |
| [14] | Villar R, Held A A, Merino J. Comparison of methods to estimate dark respiration in the light in leaves of two woody species. Plant Physiology, 1994, 105(1): 167-172. |
| [15] | Foyer C H, Noctor G. Oxygen processing in photosynthesis: regulation and signalling. New Phytologist, 2000, 146(3): 359-388. |
| [16] | Feng Q, Hu D, Li N. Comparative analysis of leaf gas exchange and chlorophyll fluorescence of Euonymus japonicus Thunb by field experiment in the typical urban and suburban areas in Beijing. Acta Ecologica Sinica, 2009, 29(7): 3477-3484. |
| [17] | Yu Y Y, Hu D, Guo E H, Xiao Q, Liu M, Zhang X F. Effects of urban shading on photosynthesis of Euonymus japonicas. Acta Ecologica Sinica, 2011, 31(19): 5646-5653. |
| [18] | Zhao J J, Ouyang Z Y, Zheng H, Xu W H, Wang X K. Species composition and spatiol structure of plants in urban parks of Beijing. Chinese Journal of Applied Ecology, 2009, 20(2): 298-306. |
| [19] | Song Y L, Zhang S Y. The study on heat island effect in Beijing during last 40 years. Chinese Journal of Eco-Agriculture, 2003, 11(4): 126-129. |
| [20] | Guo Y Q, Borjigidai A, Gao Q Z, Duan M J, Ganzhu Z B, Wan Y F, Li Y E, Guo H B. Photosynthetic characteristics of Stipa purpurea under irrigation in northern Tibet and its short-term response to temperature and CO2 concentration. Chinese Journal of Plant Ecology, 2011, 35(3): 311-321. |
| [21] | Xu X L, Jin Z X, He W M, Wang X L, Che X X. Effects of different day/ night warming on the photosynthetic characteristics and chlorophyll fluorescence parameters of Sinocalycanthus chinensis seedlings. Acta Ecologica Sinica, 2012, 32(20): 6343-6353. |
| [22] | Mccashin B G, Cossins E A, Canvin D T. Dark respiration during photosynthesis in wheat leaf slices. Plant Physiology, 1988, 87(1): 155-161. |
| [23] | Shaw M R, Zavaleta E S, Chiariello N R, Cleland E E, Mooney H A, Field C B. Grassland responses to global environmental changes suppressed by elevated CO2. Science, 2002, 298(5600): 1987-1990. |
| [1] | 苏文华, 张光飞. 土壤温度与气温对紫花雪山报春光合作用和蒸腾作用的影响. 西北植物学报, 2002, 22(4): 824-830. |
| [16] | 冯强, 胡聃, 李娜. 典型城区与郊区环境大叶黄杨气体交换及叶绿素荧光特性比较. 生态学报, 2009, 29(7): 3477-3484. |
| [17] | 于盈盈, 胡聃, 郭二辉, 肖强, 柳敏, 张小飞. 城市遮阴环境变化对大叶黄杨光合过程的影响. 生态学报, 2011, 31(19): 5646-5653. |
| [18] | 赵娟娟, 欧阳志云, 郑华, 徐卫华, 王效科. 北京城区公园的植物种类构成及空间结构. 应用生态学报, 2009, 20(2): 298-306. |
| [19] | 宋艳玲, 张尚印. 北京市近40年城市热岛效应研究. 中国生态农业学报, 2003, 11(4): 126-129. |
| [20] | 郭亚奇, 阿里穆斯, 高清竹, 段敏杰, 干珠扎布, 万运帆, 李玉娥, 郭红宝. 灌溉条件下藏北紫花针茅光合特性及其对温度和CO2浓度的短期响应. 植物生态学报, 2011, 35(3): 311-321. |
| [21] | 徐兴利, 金则新, 何维明, 王兴龙, 车秀霞. 不同增温处理对夏蜡梅光合特性和叶绿素荧光参数的影响. 生态学报, 2012, 32(20): 6343-6353. |