 2014, Vol. 34
2014, Vol. 34文章信息
- 侯沁文, 铁军, 白海艳
- HOU Qinwen, TIE Jun, BAI Haiyan
- 山西蟒河猕猴国家级自然保护区蛾类多样性
- Preliminary investigation on the moths diversity in Manghe Rhesus Monkeys National Nature Reserve in Shanxi
- 生态学报, 2014, 34(23): 6954-6962
- Acta Ecologica Sinica, 2014, 34(23): 6954-6962
- http://dx.doi.org/10.5846/stxb201312273043
-
文章历史
- 收稿日期:2013-12-27
- 修订日期:2014-08-28






2. 太行山生态与环境研究所, 长治 046011
2. Ecological and Environmental Research Institute of Taihang Mountain, Changzhi 046011, China
生物多样性是生物及其与环境形成的生态复合体以及与此相关的各种生态过程的总和,包括动物、植物、微生物和它们所拥有的基因以及它们与其生存环境形成的复杂生态系统,它随地理位置、气候条件、地理历史过程和人为活动等会发生明显的变化[1]。由于人类不断采伐和多种方式的干扰,对生物多样性产生了深刻的影响,导致物种数减少、物种丰富度变化和特有种濒危[2, 3]。生态环境的变化是蛾类群落减少的主要原因,蛾类生存所需要的寄主植物大量减少,加之人为滥用农药,使得生境进一步破坏,更缩小了蛾类的生存空间。同时蛾类对生境的变化也很敏感,具有广谱生物地理学和生态学探针功能[4],蛾类种类和数量可以预示温带森林的环境质量。因此,蛾类有望成为温带森林中指示鳞翅目物种多样性和森林植被组成变化的指示类群[5]。除此之外,蛾类也是重要的自然资源,在自然保护区中生活着种类众多、数量巨大的蛾类昆虫,它们是保护区生态系统中物种多样性的重要组成部分,在保护区环境的物质和能量流动与转化中起着不可忽视的作用。因此,掌握自然保护区蛾类的动态规律,及其与自然保护区环境变化的关系,及时应用于自然保护区的保护和监测中,在合理利用和保护国家自然保护区资源方面发挥着积极的作用。
山西蟒河猕猴国家级自然保护区是地处中条山东端,以保护猕猴和亚热带植被为主的森林和野生动物类型自然保护区。多年来,对保护区内植物的研究较多,主要集中在植物区系、植物资源、植物生态(包括重要经济植物以及国家重点保护的植物)等方面[6, 7],对动物的研究主要针对猕猴以及鸟类[8],对昆虫的研究尤其是对蛾类昆虫资源及其生态学等方面的研究少之又少。
鉴于此,对山西蟒河猕猴国家级自然保护区蛾类昆虫资源开展调查,在此基础上对不同生境的蛾类群落结构及其多样性特征进行分析,旨在通过探讨蛾类多样性对生境类型的生态响应,从蛾类群落结构及其多样性变化等角度,为评估保护区森林生态系统的健康状况提供科学依据,为保护区的长期发展规划和保护区内物种多样性保护提供一定的理论基础。
1 研究地概况蟒河猕猴国家级自然保护区,位于山西省的东南部,中条山东端,距晋城阳城县30 km。东经112°22′10″—112°31′35″,北纬35°12′50″—37°17′20″。东起三盘山,西至指柱山,南至河南济源省界,北至花园岭。海拔最高达1572 m,最低520 m,总面积约5600 hm2。蟒河一带冬季温和,夏季凉爽,年平均气温为14 ℃,无霜期180 d,年降水量500—800 mm,空气湿润,受季风影响不大,适宜各种动植物生存。
保护区四周环山,中间谷地,境内沟壑纵横,地形复杂,气候多样,植物种类较丰富。据报道,蟒河猕猴国家级保护区种子植物有882种,隶属102科391属,分别占山西省种子植物总科数的75.9%,总属数的62.3%,总种数的52.4%[9]。在本区的区系成分中,属种数量较多的科有(以属数多少为序)菊科(Compositae)、禾本科(Gramineae)、唇形科(Labiatae)、蔷薇科(Rosaceae)、豆科(Leguminosae)、毛茛科(Ranunculaceae)、十字花科(Cruciferae)和百合科(Liliaceae)等。它们在该地区的区系组成中占有重要作用。
2 研究方法 2.1 调查方法选择山西蟒河自然保护区针阔叶混交林、阔叶落叶林、杂木林和灌木林4种植被类型为样地进行研究,4个样地的植物组成见表 1。
| 植被类型 Vegetation types | 植物组成Plant composition | |
| 木本植物Woody plants | 草本植物Herb plants | |
| 针阔叶混交林 Mixed coniferous and broad- leaved forest | 油松Pinus tabuliformis 红桦Betula albo-sinensis 槲栎Quercus aliena 鹅耳枥Carpinus turczaninowii 小叶鹅耳枥Carpinus stipulata 橿子栎Quercus baronii 黄栌Cotinus coggygria 接骨木Sambucus williamsii 石生悬钩子Rubus saxatilis等 | 石竹Dianthus chinensis 紫菀Aster tataricus 地榆Sanguisorba officinalis 披针叶苔草Carex lanceolata 轮叶黄精Polygonatum verticillatum 鹿药Maianthemum japonicum等 |
| 阔叶落叶林 Broad-leaved deciduous forest | 鹅耳枥、槲栎、栓皮栎Quercus variabilis 构树Broussonetia papyrifera 稠李Padus avium 黄栌、黄刺玫Rosa xanthina 杭子梢Campylotropis macrocarpa 五味子Schisandra chinensis等 | 风毛菊Saussurea japonica 三脉紫菀Aster ageratoides 贝加尔唐松草Thalictrum baicalense 华北耧斗菜Aquilegia yabeana等 |
| 杂木林 Miscellaneous wood forest | 野核桃Juglans cathayensis 旱柳Salix matsudana 金银忍冬Lonicera maackii 小叶鼠李Rhamnus parvifolia 山茱萸Cornus officinalis 南蛇藤Celastrus orbiculatus 陕西荚蒾Viburnum schensianum 三裂绣线菊Spiraea trilobata 杠柳Periploca sepium等 | 博落回Macleaya cordata 糙苏Phlomis umbrosa 唐松草Thalictrum aquilegiifolium var. sibiricum 川续断Dipsacus asper 白屈菜Chelidonium majus 旋覆花Inula japonica等 |
| 灌木林 Shrubbery | 连翘Forsythia suspensa、黄栌 灰栒子Cotoneaster acutifolius 胡枝子Lespedeza bicolor 荆条Vitex negundo var. heterophylla 接骨木、白刺花Sophora davidii 土庄绣线菊Spiraea pubescens等 | 毛茛Ranunculus japonicus 夏至草Lagopsis supina 毛地黄Digitalis purpurea 委陵菜Potentilla chinensis 茜草Rubia cordifolia 蒲公英Taraxacum mongolicum 酸模Rumex acetosa 鹅绒藤Cynanchum chinense等 |
2012年6—8月对蛾类进行诱捕采集。每两日调查1次,遇天气不良,则顺延一日。每种植被类型各设3个样点,样点间距离>50 m,调查方法为灯诱,变频汽油发电机(HS1000IS)发电,诱集灯为白色250 W高压汞灯,幕布选用210 cm×150 cm的白化纤布,灯诱时间为20:00—24:00。用竹竿把幕布挂于诱集灯后方15 cm处,清除幕布四周4—5 m范围内的杂草,采集所有落于幕布前后面的蛾类。为鉴定方便,大小蛾类分开采集,大型蛾类使用毒瓶,小型蛾类使用指形管,用乙酸乙酯作为毒杀剂,杀死的蛾类放于储虫瓶(管)中,尽量避免摇动,并注明采集时间、地点及采集人。每天早上将收回的虫体针插、干燥,放置于昆虫盒内。继而按科分类整理后,带回实验室进一步鉴定。
分类鉴定到种,鉴定主要依据《中国动物志·舟蛾科》[10]、《中国动物志·夜蛾科》[11]、《中国经济昆虫志·夜蛾科(二)》[12]、《中国经济昆虫志·夜蛾科(四)》[13]、《中国动物志·毒蛾科》[14]、《中国动物志·枯叶蛾科》[15]、《河南昆虫志·刺蛾科、枯叶蛾科、舟蛾科、灯蛾科、毒蛾科、鹿蛾科》[16]、《中国动物志·尺蛾科、尺蛾亚科》[17]、《中国动物志·灯蛾科》[18]等。所有采集的蛾类标本均保存在长治学院昆虫标本室内。
2.2 数据分析 2.2.1 多样性分析物种丰富度(S):每个样地中出现的物种数。多度(N):个体总数。多样性指数分别采用Simpson指数(D)、Shannon-Wiener指数(H′)、Brillouin指数(H)和McIntosh指数(DMc)。
Simpson指数公式:
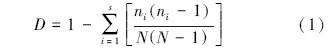
式中,ni为抽样中第i个物种的个体数量,N为抽样中所有物种的个体总和,s为物种总数[19]。
Shannon-Wiener指数公式:
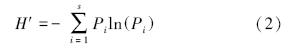
式中,Pi是第i种的个体比例,即Pi= Ni /N ,Ni=第i种的个体数,N=全部物种的个体总数[19]。
Brillouin指数公式:
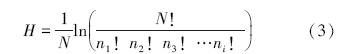
式中,n1为抽样中第1个物种的个体数量,n2为抽样中第2个物种的个体数量,n3为抽样中第3个物种的个体数量,如此类推,N为抽样中所有物种的个体总和[19]。
McIntosh指数公式:
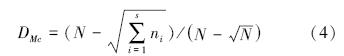
式中,ni为抽样中第i个物种个体数量,N为抽样中所有物种个体总和,S为物种数。
均匀度(J′)采用Pielou公式:

式中,H′为Shannon-Wiener多样性指数,S为群落中物种数[19]。
2.2.2 对数正态模型参数估计对数正态分布模型为:
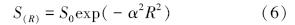
式中,S(R)是第R倍频程中的物种数量,S0是对数正态分布的众数倍频程物种数,α是与分布有关的参数[19]。
2.2.3 群落相似性指标卡方距离(dij):
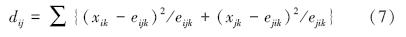
式中,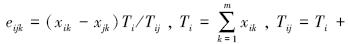
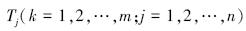 [19]。
[19]。
欧氏距离(dij):
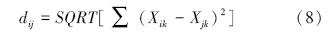
式中,dij为i群落与j群落之间的欧氏距离;Xik为i群落第k个指标的值;Xjk为j群落第k个指标的值[20]。
3 结果 3.1 蛾类群落物种组成在4种植被类型样地中共捕获蛾类4709只,隶属24科184种,其中螟蛾科物种数和个体数最多,有26种950只,分别占总种数和总个体数的14.1%和20.2%。其次是尺蛾科,有25种698只,所占比例分别为13.6%和14.8%。草螟科物种数仅次于尺蛾科,为22种,个体数量却高于前者,为739只,占全部个体总量的15.7%。部分科只有1种,如网蛾科、斑蛾科、鞘蛾科和木蠹蛾科等。
从不同林型的蛾类群落组成来看,针阔叶混交林中有20科100种,优势种群不明显;阔叶落叶林中有14科74种,其中尺蛾科(20.3%)占优势地位,其次是夜蛾科13.5%;杂木林中有18科102种,其中草螟科(17.6%)和夜蛾科(15.7%)二者占绝对优势,其次是螟蛾科(13.7%);灌木林中有20科132种,其中螟蛾科(25.2%)数量占绝对优势,其次是夜蛾科(15.5%),尺蛾科(13.6%)次之。
从蛾类各科水平来看,螟蛾科在4种植被类型中物种数和个体数都最高。其中,灌木林的物种数和个体数皆最多,有22种357只;阔叶落叶林的螟蛾科种数最少,有9种;针阔叶混交林个体数量最少,有145只。仅次于螟蛾科的是尺蛾科,在灌木林的种数最多,有16种,在杂木林中种数最少,有10种。第三为草螟科,在杂木林中种数及个体数都最多,有18种232只。还有部分科的蛾类种类很少,如木蠹蛾科只出现在针阔叶混交林中,鞘蛾科和斑蛾科只出现在灌木林中,均为1种。刺蛾科和枯叶蛾科除了阔叶落叶林之外,在其它植被类型中都有出现,但不超过3种。羽蛾科和展足蛾科在针阔叶混交林和杂木林中出现,不超过3种。鹿蛾科除了杂木林,在其它植被类型中均出现。麦蛾科在4种植被类型中均为1种,其它蛾类在不同植被中种数大致相当。
| 科名 Families | A | B | C | D | 合计 Total | 比例Proportion/% | ||||||
| S | N | S | N | S | N | S | N | S | N | S | N | |
| A:针阔叶混交林Mixed coniferous broad-leaved forest;B:阔叶落叶林Broad-leaved deciduous forest;C:杂木林Miscellaneous wood forest;D:灌木林Shrubbery;S:种数Species number;N:个体数 Individuals number | ||||||||||||
| 舟蛾科Notodontidae | 11 | 110 | 8 | 171 | 8 | 65 | 13 | 128 | 17 | 474 | 9.2 | 10.1 |
| 夜蛾科Noctuidae | 12 | 150 | 10 | 255 | 16 | 130 | 14 | 219 | 18 | 754 | 9.8 | 16.0 |
| 灯蛾科Arctiidae | 7 | 40 | 6 | 48 | 7 | 47 | 10 | 64 | 12 | 199 | 6.5 | 4.2 |
| 卷蛾科Tortricoidae | 10 | 84 | 7 | 50 | 10 | 42 | 13 | 78 | 16 | 254 | 8.7 | 5.4 |
| 毒蛾科Lymantriidae | 2 | 20 | 1 | 30 | 2 | 32 | 5 | 54 | 4 | 136 | 2.2 | 2.9 |
| 尺蛾科Geometridae | 12 | 94 | 15 | 289 | 9 | 122 | 16 | 193 | 25 | 698 | 13.6 | 14.8 |
| 螟蛾科Pyralidae | 13 | 145 | 9 | 232 | 14 | 256 | 22 | 357 | 26 | 950 | 14.1 | 20.2 |
| 草螟科Crambidae | 9 | 98 | 8 | 223 | 18 | 232 | 11 | 146 | 22 | 739 | 12.0 | 15.7 |
| 天蛾科Sphingidae | 4 | 33 | 3 | 78 | 5 | 56 | 6 | 89 | 12 | 256 | 6.5 | 5.4 |
| 刺蛾科Limacodidae | 2 | 17 | 0 | 0 | 2 | 21 | 3 | 29 | 4 | 67 | 2.2 | 1.4 |
| 鹿蛾科Ctenuchidae | 1 | 2 | 1 | 1 | 0 | 0 | 2 | 4 | 2 | 7 | 1.1 | 0.1 |
| 蚕蛾科Bombycidae | 2 | 5 | 3 | 12 | 1 | 6 | 3 | 11 | 2 | 34 | 1.1 | 0.7 |
| 大蚕蛾科Saturniidae | 2 | 12 | 1 | 23 | 1 | 8 | 2 | 15 | 2 | 58 | 1.1 | 1.2 |
| 细蛾科Gracilariidae | 1 | 2 | 0 | 0 | 0 | 0 | 3 | 12 | 5 | 14 | 2.7 | 0.3 |
| 网蛾科Thyrididae | 1 | 1 | 0 | 0 | 0 | 0 | 1 | 2 | 1 | 3 | 0.5 | 0.1 |
| 斑蛾科Zygaenidae | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 1 | 1 | 1 | 0.5 | 0.0 |
| 枯叶蛾科Lasiocampidae | 2 | 5 | 0 | 0 | 1 | 10 | 1 | 1 | 2 | 16 | 1.1 | 0.3 |
| 羽蛾科Pterophoridae | 3 | 6 | 0 | 0 | 1 | 7 | 0 | 0 | 2 | 13 | 1.1 | 0.3 |
| 木蠹蛾科Cossidae | 1 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 0.5 | 0.0 |
| 潜蛾科Lyonetiidae | 0 | 0 | 0 | 0 | 2 | 3 | 1 | 1 | 2 | 4 | 1.1 | 0.1 |
| 展足蛾科Heliodinidae | 2 | 3 | 0 | 0 | 2 | 3 | 0 | 0 | 2 | 6 | 1.1 | 0.1 |
| 麦蛾科Gelechiidae | 1 | 1 | 1 | 2 | 1 | 1 | 1 | 1 | 2 | 5 | 1.1 | 0.1 |
| 菜蛾科Plutellidae | 2 | 4 | 1 | 1 | 2 | 5 | 3 | 7 | 3 | 17 | 1.6 | 0.4 |
| 鞘蛾科Coleophoridea | 0 | 0 | 0 | 0 | 0 | 0 | 1 | 2 | 1 | 2 | 0.5 | 0.0 |
| 总计 | 100 | 834 | 74 | 1415 | 102 | 1046 | 132 | 1414 | 184 | 4709 | 100.0 | 100.0 |
分别对4个植物群落中的蛾类多度进行对数正态分布模型拟合及参数估计,结果见表 3和图 1。由表 3可知,针阔叶混交林群落(P=0.517)、灌木林群落(P=0.939)和杂木林群落(P=0.613)均表现为对数正态模型,其中灌木林群落是最优符合;说明这三类群落均环境条件优越、物种丰富度高。其中,灌木林群落环境条件最优越,蛾类丰富度最大,而优势种却最少,处于中间物种较多,说明灌木林群落中蛾类寄主资源总量最大,优势种和稀有种较少。针阔叶混交林群落类似于杂木林群落,蛾类种数较少,说明这两种植被中蛾类寄主资源总量较小。阔叶落叶林蛾类多度不符合对数正态模型(P<0.05),物种数量最少。从图 1可以看出,阔叶落叶林中处于中间的物种在逐步消亡,而优势种数量在增加,这种现象除了和该植被类型中植物种类少有关系外,还和其环境条件变化有一定关系。
| 植被类型 Vegetation type | 种数 Species number | 个数 Number | α | 物种估计 Species estimate | χ2 | df | p |
| 针阔叶混交林Mixed coniferous and broad-leaved forest | 100±12 | 830±25 | 0.637 | 103 | 3.253 | 4 | 0.517 |
| 阔叶落叶林Broad-leaved deciduous forest | 74±8 | 1415±34 | 0.786 | 79 | 13.217 | 6 | 0.039 |
| 杂木林Miscellaneous wood forest | 101±13 | 991±27 | 0.763 | 102 | 1.257 | 5 | 0.939 |
| 灌木林Shrubbery | 132±18 | 1272±26 | 0.634 | 133 | 3.572 | 5 | 0.613 |

|
| 图 1 4种植被类型中蛾类群落种 Fig. 1 Species-abundance curves for moths in four vegetation types |
经拟合得出针阔叶混交林、杂木林和灌木林蛾类种与多度曲线公式分别为S(R)=25exp(-0.642R2),物种估计为103种;S(R)=31exp(-0.762R2),物种估计为102种;S(R)=27exp(-0.632R2),物种估计为133种。
3.3 蛾类群落的多样性和均匀度多样性指数分别采用Simpson指数(D)、Shannon-Wiener指数(H′)、Brillouin指数(H)和McIntosh指数(DMc),计算结果见表 4。
| 植被类型 Vegetation type | 种数 Species number | 个体数 Number | 多样性diversity | 均匀度 Evenness | |||
| D | H′ | H | DMc | ||||
| D:Simpson指数;H′:Shannon-Wiener指数;H:Brillouin指数;DMc:McIntosh指数 | |||||||
| 针阔叶混交林 Mixed coniferous and broad-leaved forest | 100±12 b | 830±25 d | 0.9727 a | 5.7463 b | 5.4612 b | 0.8712 ab | 0.8649 b |
| 阔叶落叶林 Broad-leaved deciduous forest | 74±8 c | 1415±34 a | 0.9525 b | 4.9144 c | 4.7791 c | 0.8017 c | 0.6914 d |
| 杂木林 Miscellaneous wood forest | 101±13 b | 991±27 c | 0.9784 a | 5.5838 b | 5.3636 b | 0.8273 b | 0.8113 c |
| 灌木林Shrubbery | 132±18 a | 1272±26 b | 0.9728 a | 5.9646 a | 5.7032 a | 0.8774 a | 0.8958 a |
从总体来看,4种不同植被类型蛾类群落多样性与均匀度是一致的,但H′和H的区分度较好,D和DMc的区分度较差,可见用Shannon-Wiener指数和Brillouin指数来描述蛾类群落的多样性更为合适。相比而言,灌木林的蛾类多样性指数和均匀度最高,而阔叶落叶林的蛾类多样性指数和均匀度指数最低,说明灌木林中蛾类的寄主植物种类最丰富、数量多,而阔叶落叶林中蛾类的寄主植物种类少,环境条件比较单一。总的来说,在相同气候条件下,蛾类群落多样性受到植被种类的影响。同时也表明蛾类对环境具有敏感性,蛾类群落的变化表明蛾类所处环境的变迁。
3.4 蛾类群落聚类分析采用欧氏距离和卡方距离两种方式,用最短距离法进行聚类,结果见图 2。从欧氏距离聚类结果来看,杂木林和灌木林首先聚为一类,然后与针阔叶混交林聚在一起,最后再与阔叶落叶林聚类;而从卡方距离聚类结果来看,杂木林、灌木林和针阔叶混交林聚在一起,最后再与阔叶落叶林聚类。说明杂木林、灌木林和针阔叶混交林的蛾类群落组成相似,而与阔叶落叶林的蛾类群落相似性较低。可见,杂木林、灌木林和针阔叶混交林的蛾类群落所处环境条件相近,环境保护得相对较好,群落结构稳定。相反,由于植被类型单一且受外界因素的影响,阔叶落叶林的蛾类群落所处环境条件正在不断变化,群落结构也变得不稳定,其中一些物种已经消亡[21, 22]。

|
| 图 2 蛾类群落欧氏距离和卡方距离聚类图 Fig. 2 Clustering graph of euclidean distance and chi-square distance for moth community A: 针阔叶混交林; B: 阔叶落叶林; C: 杂木林; D: 灌木林 |
蛾类各科在4种不同植被类型中的比例分配有所不同。在针阔叶混交林中优势种群不明显,螟蛾、夜蛾、尺蛾、舟蛾、卷蛾和草螟蛾的种数在10—13种;在阔叶落叶林中尺蛾占绝对优势,相对前者,螟蛾种数减少,而草螟蛾种数增加;在杂木林中草螟蛾占绝对优势,其次是夜蛾;在灌木林中螟蛾数量占绝对优势,其次是尺蛾,再其次是夜蛾。在这4种植被类型中,阔叶落叶林蛾类种类最少,只有74种,而数量最多,尤其是尺蛾数量明显多于其它3个群落。这种现象除了可能和植被类型有关外,还可能是受到外界因素的影响,植物群落结构受到干扰而逐步演化,其内的昆虫群落也与植物群落共同演化发展,优势类群明显,而其他类群则可能受抑,其中一部分物种灭亡[22]。各植被类型蛾类群落结构差异较大,分类阶元越低,蛾的种类、数量、优势种类及优势个体比例受环境影响越大。植物种类与蛾类种类、数量呈正相关,即植物群落越复杂,则蛾类群落越复杂,但是优势种数及优势个体比例却越低。
4.2 蛾类群落的种-多度关系群落指标主要包括种类数、种群数量、多样性指数、均匀度等衡量群落稳定性的重要指标。在判定森林生态系统的健康状态时,多数学者认为,在健康的生态系统中,多样性-多度关系常呈对数正态分布;而在恶劣条件下,多样性-多度格局常常有所变化且不再表现为这种情况;多样性和丰度分布偏离对数正态分布越远,群落或其所在的生态系统就越不健康[23]。在多样性-多度关系的基础上,再结合昆虫群落的种类数、种群数量、多样性指数和均匀度等指标,往往能够提供生态系统偏离健康状态程度的依据。本文也认为阔叶落叶林中蛾类多样性指数受均匀度影响,是由于种-多度关系趋向于生态位优先占领假说,个体数量不呈对数正态分布。表明了阔叶落叶林环境条件不稳定,群落结构变化大,生态系统偏离健康状态。而针阔叶混交林、杂木林和灌木林群落的种-多度曲线呈对数正态分布时,一般情况下多样性指数与均匀度一致。说明针阔叶混交林、杂木林和灌木林群落环境比较好,生态系统健康状态良好,未出现退化的趋势。认为平衡稳定的群落多度曲线通常服从对数正态分布[24]。符合对数正态分布的群落多属于环境条件优越,物种丰富度高的群落[20]。种-多度关系对于检测无脊椎动物对森林干扰的反应比多样性指数能提供更多的信息[25]。
4.3 蛾类群落的多样性均匀度和丰富度(物种数)是与多样性指数密切相关的两个参数。荒漠草原昆虫群落的多样性指数与均匀度是一致的,表明群落结构是稳定的[26]。稻田昆虫群落则在不同季节多样性指数与均匀度不一致[27]。相同气候条件下,蛾类群落多样性受到植被多样性的影响,同时也表明蛾类昆虫对环境的敏感性,相反,蛾类群落的变化表明蛾类昆虫所处环境的变迁[20, 28]。在美国选取保护较好的山林及破坏较为严重,植被较为混杂的地区进行调查,发现尺蛾科多样性在受干扰地区明显增加,分析认为毗邻生境中昆虫的扩散活动是造成这种现象的部分原因,而且结果中较高的单个数量的物种数也预示着那些路过物种的比例较高[29]。蟒河猕猴国家级自然保护区不同植被类型中蛾类的多样性指数与均匀度表现为正相关或弱的正相关,但是落叶阔叶林中蛾类种类最少,数量最多且均匀指数最低,其中尺蛾科种类和数量也是最多的,说明该生态环境受到的干扰最严重,生态环境正趋于恶化。
4.4 植被与蛾类的相关性植被是反映保护区生态环境的重要标志,是保护区蛾类生存的重要依据。植被种类越丰富,则蛾类种就越多;植被资源越丰富,种的个体数就越多。不同林型蛾类群落组成的变化主要是蛾类各科在不同林型间的变化所导致[30]。说明蛾类群落种类与数量和植被有相关性。从蛾类总物种丰富度和蛾类个体总数上看,灌木林中蛾类种类数量明显高于其它植被类型,针阔叶混交林和杂木林则蛾类种类相差无几,但个体数量后者明显多于前者,可能是由于这两种植被类型植物丰富度相似,而植物资源量后者远多于前者造成的[22, 31]。不同的植被状况和气候条件的各种组成产生了各林带间多样性参数的较大差异,表明了蛾类对微环境的敏感性,这与尤平等人的结论相一致,说明把蛾类作为环境变化的指示物是可行的[19]。有关蟒河猕猴国家级保护区植被与蛾类群落定量的相关性,有待进一步研究。
| [1] | Zhang X B, Pan J Z, Gao B J. Research progress and perspective of genetic diversity in insect. Hebei Journal of Forestry and Orchard Research, 2009, 24(4): 433-438, 444-444. |
| [2] | Choi S W, Park M R N, Kim H. Differences in moth diversity in two types of forest patches in an agricultural landscape in Southern Korea-effects of habitat heterogeneity. Journal of Ecology and Field Biology, 2009, 32(3): 183-189. |
| [3] | Zhang X, Hu H Y, Lu Z Z. Butterfly diversity and vertical distribution in eastern Tianshan Mountain in Xinjiang. Acta Ecologica Sinica, 2013, 33(17): 5329-5338. |
| [4] | Samways M J. Insects in biodiversity conservation: some perspectives and directives. Biodiversity and Conservation, 1993, 2(3): 258-282. |
| [5] | Zhang J L, Li H F. Biodiversity of nature reserve in QinLing. Chinese Biodiversity, 1997, 5(2): 155-156. |
| [6] | Zhang G P, Zhang J G, Ru W M. Study on the structure of Taxus mairei community and population in Manghe, Shanxi. Journal of Shanxi University: Natural Science Edition, 2003, 26(2): 169-172. |
| [7] | Zhang Y B, Zhang F, Zhao Y S, Fan M X. Analysis on the flora of seed plants in Manghe Nature Reserve, ShanXi. Bulletin of Botanical Research, 2003, 23(4): 500-506. |
| [8] | Zhao Y S, Tian S W, Tian D Y, Ru L J. A Study on the reproductive ecology of Carduelis sinica in Manghe Natural Reserves, Shanxi. Journal of Shanxi University: Natural Science Edition, 2000, 23(2): 168-171. |
| [9] | Tian S W, Zhang J, Zhang Q X. Study report on vegetation Manghe Nature Reserve. Journal of Taiyuan Normal University: Natural Science Edition, 2012, 11(4): 142-144. |
| [10] | Wu C S, Fang C L. Fauna Insects of China, vol. 31. Lepidoptera: Notodontidae. Beijing: Science Press, 2003. |
| [11] | Chen Y X. Fauna Insects of China, vol. 16. Lepidoptera: Noctuidae. Beijing: Science Press, 1999. |
| [12] | Zhu H F, Yang J K, Lu J R, Chen Y X. Economic Insect Fauna of China, vol. 6. Lepidoptera: Noctuidae (2th part). Beijing: Science Press, 1964. |
| [13] | Chen Y X. Economic Insect Fauna of China, vol. 32. Lepidoptera: Noctuidae(4th part). Beijing: Science Press, 1985. |
| [14] | Zhao Z L. Fauna Insects of China, vol. 30. Lepidoptera: Lymantriidae. Beijing: Science Press, 2003. |
| [15] | Liu Y J, Wu C S. Fauna Insects of China, vol. 47. Lepidoptera: Lasiocampidae. Beijing: Science Press, 2006. |
| [16] | Wu C S, Fang C L. Insects of Henan. Lepidoptera: Limacodidae, Lasiocampidae, Notodontidae, Noctuidae, Liparidae, Amatidae. Beijing: Science Press, 2010. |
| [17] | Han H X, Xue D Y. Fauna Insects of China, vol. 54. Lepidoptera: Geometridae Geometrinae. Beijing: Science Press, 2011. |
| [18] | Fang C L. Fauna Insects of China, vol. 19, Lepidoptera: Noctuidae. Beijing: Science Press, 2000. |
| [19] | Tang Q Y, Feng M G. DPS Data Processing System-Experimental Design, Statistical Analysis and Modeling. Beijing: Science Press, 2006. |
| [20] | You P, Li H H. Species richness and diversity of moth communities in Tianjin Wetlands; implications for environmental management. Acta Ecologica Sinica, 2006, 26(3): 629-637. |
| [21] | Jia Y Z, Zhao X H, Meng Q F. Insect diversity along a successional gradient in conifer and broad-leaved mixed forests in Changbai Mountain. Acta Entomologica Sinica, 2009, 52(11): 1236-1243. |
| [22] | Wang Z, Ji L Z, Zhang Y, Yi X M. Moth community structure and diversity in three forest types in Changbai Mountains of Northeast China. Chinese Journal of Ecology, 2012, 31(5): 1214-1220. |
| [23] | Zhang H Y, Ou X H. Using insect for indicator to monitor and assess forest ecosystem health. World Forestry Research, 2006, 19(4): 22-25. |
| [24] | May R M. Theoretical Ecology: Principles and Applications. Oxford: Blackwell, 1981. |
| [25] | Hawes J, Da C, Motta S, Overal W L, Barlow J, Gardner T A, Peres C A. Diversity and composition of amazonian moths in primary, secondary and plantation forests. Journal of Tropical Ecology, 2009, 25(3): 281-300. |
| [26] | He D H, Tian C, Ren G D, Hao F M, Ma S Y. A preliminary study on the community structure of desert forage grassland. Grassland of China, 1988, 10(6): 24-28. |
| [27] | Wang F H, Chen C M. Studies on the structure of the rice pest-natural enemy community and diversity under IPM area and chemical control area. Acta Ecologica Sinica, 1986, 6(2): 159-170. |
| [28] | Liu W P, De H L. The butterfly diversities in Muli Sichuan Province. Acta Ecologica Sinica, 1997, 17(3): 266-271. |
| [29] | Brehm G, Fiedler K. Diversity and community structure of geometrid moths of disturbed habitat in a montane area in the Ecuadorian Andes. Journal of Research on the Lepidoptera, 2005, 38: 1-14. |
| [30] | Huang Z L. The interactions of population dynamics of Thalassodes quadraria and the plant community structure and climate factors in Dinghushan. Chinese Journal of Ecology, 2000, 19(3): 24-27, 31-31. |
| [31] | Meng L, Li B P. Guild structure and species diversity of insects in saltcedar woodlands in Xinjiang, West China. Acta Ecologica Sinica, 2006, 25(2): 189-193. |
| [1] | 张学卫, 潘建芝, 高宝嘉. 昆虫遗传多样性研究及展望. 河北林果研究, 2009, 24(4): 433-438, 444-444. |
| [3] | 张鑫, 胡红英, 吕昭智. 新疆东部天山蝶类多样性及其垂直分布. 生态学报, 2013, 33(17): 5329-5338. |
| [5] | 张金良, 李焕芳. 秦岭自然保护区群的生物多样性. 生物多样性, 1997, 5(2): 155-156. |
| [6] | 张桂萍, 张建国, 茹文明. 山西蟒河南方红豆杉群落和种群结构研究. 山西大学学报: 自然科学版, 2003, 26(2): 169-172. |
| [7] | 张殷波, 张峰, 赵益善, 樊敏霞. 山西蟒河自然保护区种子植物区系研究. 植物研究, 2003, 23(4): 500-506. |
| [8] | 赵益善, 田随味, 田德雨, 茹李军. 蟒河自然保护区金翅雀的繁殖生态学研究. 山西大学学报: 自然科学版, 2000, 23(2): 168-171. |
| [9] | 田随味, 张军, 张青霞. 蟒河自然保护区植被考察报告. 太原师范学院学报: 自然科学版, 2012, 11(4): 142-144. |
| [10] | 武春生, 方承莱. 中国动物志昆虫纲 (第三十一卷, 鳞翅目: 舟蛾科). 北京: 科学出版社, 2003. |
| [11] | 陈一心. 中国动物志昆虫纲 (第十六卷, 鳞翅目: 夜蛾科). 北京: 科学出版社, 1999. |
| [12] | 朱弘复, 杨集昆, 陆近仁, 陈一心. 中国经济昆虫志 (第六册, 鳞翅目: 夜蛾科) (二). 北京: 科学出版社, 1964. |
| [13] | 陈一心. 中国经济昆虫志 (第三十二册, 鳞翅目: 夜蛾科) (四). 北京: 科学出版社, 1985. |
| [14] | 赵仲苓. 中国动物志昆虫纲 (第三十卷, 鳞翅目: 毒蛾科). 北京: 科学出版社, 2003. |
| [15] | 刘友樵, 武春生. 中国动物志昆虫纲 (第四十七卷, 鳞翅目: 枯叶蛾科). 北京: 科学出版社, 2006. |
| [16] | 武春生, 方承莱. 河南昆虫志 (鳞翅目: 刺蛾科、枯叶蛾科、舟蛾科、灯蛾科、毒蛾科、鹿蛾科). 北京: 科学出版社, 2010. |
| [17] | 韩红香, 薛大勇. 中国动物志昆虫纲 (第五十四卷, 鳞翅目: 尺蛾科 尺蛾亚科). 北京: 科学出版社, 2011. |
| [18] | 方承莱. 中国动物志昆虫纲 (第十九卷, 鳞翅目: 灯蛾科). 北京: 科学出版社, 2000. |
| [19] | 唐启义, 冯明光. DPS数据处理系统—实验设计、统计分析及模型优化. 北京: 科学出版社, 2006. |
| [20] | 尤平, 李后魂. 天津湿地蛾类丰富度和多样性及其环境评价. 生态学报, 2006, 26(3): 629-637. |
| [21] | 贾玉珍, 赵秀海, 孟庆繁. 长白山针阔混交林不同演替阶段的昆虫多样性. 昆虫学报, 2009, 52(11): 1236-1243. |
| [22] | 王珍, 姬兰柱, 张悦, 易雪梅. 长白山三种林型对蛾类群落结构和多样性的影响. 生态学杂志, 2012, 31(5): 1214-1220. |
| [23] | 张红玉, 欧晓红. 以昆虫为指示物种监测和评价森林生态系统健康初探. 世界林业研究, 2006, 19(4): 22-25. |
| [26] | 贺答汉, 田畴, 任国栋, 郝峰茂, 马世渝. 荒漠草原昆虫的群落结构及其演替规律初探. 中国草地, 1988, 10(6): 24-28. |
| [27] | 万方浩, 陈常铭. 综防区和化防区稻田害虫天敌群落组成及多样性的研究. 生态学报, 1986, 6(2): 159-170. |
| [28] | 刘文萍, 邓合黎. 木里蝶类多样性的研究. 生态学报, 1997, 17(3): 266-271. |
| [30] | 黄忠良. 樟翠尺蛾种群动态与植物群落结构及气候因子的关系. 生态学杂志, 2000, 19(3): 24-27, 31-31. |
| [31] | 孟玲, 李保平. 新疆柽柳林地昆虫群落取食功能团结构及物种多样性研究. 生态学杂志, 2006, 25(2): 189-193. |