 2014, Vol. 34
2014, Vol. 34文章信息
- 宗盛伟, 许嘉巍, 吴正方, 乔琳琳, 王丹丹, 孟祥君, 杜海波, 王雷, 王丹, 王鹏, 敖小龙, 夏毓璘, 吴福梅
- ZONG Shengwei, XU Jiawei, WU Zhengfang, QIAO Linlin, WANG Dandan, MENG Xiangjun, DU Haibo, WANG Lei, WANG Dan, WANG Peng, AO Xiaolong, XIA Yulin, WU Fumei
- 长白山西坡小叶章侵入苔原带过程及影响
- Analysis of the process and impacts of Deyeuxia angustifolia Invasion on the Alpine Tundra, Changbai Mountain
- 生态学报, 2014, 34(23): 6837-6846
- Acta Ecologica Sinica, 2014, 34(23): 6837-6846
- http://dx.doi.org/10.5846/stxb201302270319
-
文章历史
- 收稿日期:2013-02-27
- 网络出版日期:2014-03-18






2. 吉林财经大学工商管理学院, 长春 130024
2. Tourism Management, Jilin University of Finance and Economics, Changchun 130024, China
山地生态系统是响应气候及环境变化的敏感区域。近年来,受气候变化及人类活动干扰影响,山地生态系统的植被发生了很大的变化。低海拔植物向高海拔地区侵入现象成为研究热点,但大多表现为树木或者灌木的上侵[1]。草本植物侵入高海拔地区的现象较为少见[2],Tsuyuzaki和Shiro研究发现日本Usu火山喷发后的植被演替过程中,湿生植物不是来自于土壤种子库以及周边植物的繁殖,而是来源于山峰之外的低海拔生境[3];Wearne和Morgan发现了澳大利亚东部金雀儿草侵入亚高山植被现象,认为这是一种本地生态系统重构[1]。在高山生态系统中,一旦发生入侵现象,将会产生严重后果,包括物种的灭绝、生态系统功能退化等[4, 5, 6]。因此,植物入侵研究是保护高山地区生态系统安全的重要研究领域。
长白山苔原带是我国仅有的三处高山苔原带之一。目前,青藏高原苔原带以及阿尔泰山苔原带未见有植物上侵的研究报道,而长白山苔原带的植被变化一直令人瞩目。长白山北坡岳桦(Betula ermanii Charm.)上侵苔原带现象已经得到认同,并将其归因于气候变暖的影响[4]。然而与之对应的长白山西坡苔原带未发现强烈的岳桦上侵,且该地区的植被变化鲜有人关注。钱家驹编写了长白山苔原带植物名录[5],钱宏对长白山苔原带植被进行了分类调查[6],尚未涉及长白山西坡苔原带植被变化。然而长白山西坡植被在20年间发生了巨大的变化,1986年台风过境之后,长白山西坡岳桦林带产生了风倒区,形成了大量的林窗,这种干扰对于苔原带的植被变化有着不可忽视的作用。另外,经过文献分析以及多年的野外实地调察,发现西坡岳桦林下的小叶章(Deyeuxia. angustifolia (Kom.) Y. L. Chang)为代表的草本植物逐步侵入到海拔更高的苔原带,形成与基质差异明显的斑块[7]。从目前的植被分布状况上看,小叶章的分布范围已经覆盖了西坡苔原带较大范围,对于景观水平以及群落水平的影响十分显著。针对小叶章侵入苔原带的研究已非常必要,其首要任务在于揭示侵入的演变过程以及对于生态系统的影响。
遥感,地理信息系统和全球定位系统GPS组成的3S 技术为植物侵入的过程研究提供了技术手段,特别是在长时间序列的影像数据支持下。例如,Bradley利用Landsat Thematic Mapper(TM)影像研究了入侵植物对景观动态的影响[8];近年来光谱技术的应用主要在于区分本地植物与入侵植物的差异。例如,Walsh利用EO—1 Hyperion数据分析了入侵植物的分布[9]。高占国、张利权利用不同植物群落的光谱差异,为上海地区盐沼植被分类以及遥感监测外来种互花米草的空间分布提供了影像解译的判读依据[10]。入侵植物对本地植物群落的影响是植物入侵研究的重点。例如,Wolf从群落角度分析了草木犀上侵对高山草地的影响[11];Hejda分析了植物入侵后群落多样性的变化,认为这种变化源自于入侵植物与本地植物高度、盖度的不同[12];Flory认为植物入侵后会抑制本地植物生长,并改变群落结构[13]。小叶章侵入苔原带必将导致苔原带景观的巨大变化,其对于生态系统的影响亟需深入的研究和分析。
本文在高光谱及影像分析的基础上,结合GPS定位技术揭示小叶章侵入苔原带的演变过程;通过野外调查,对不同侵入强度的斑块进行群落调查及土壤分析;探究小叶章侵入苔原带的过程并阐述其对于苔原带产生的影响。从而为小叶章上侵机理的研究提供数据支撑;为小叶章入侵的治理提供科学依据;并丰富高山地带植物入侵研究,为全球范围内存在的高山植被带上移现象提供实证参考。
1 研究区概况研究区位于长白山苔原带西坡(41°58′—41°59′,127°59′—128°01′),为火山锥体的迎风坡的上部(海拔2 000 m以上)。研究区温度低,生长季(6—9月份)日平均气温只有5.87 ℃。降水丰富,生长季年均降水量可达958.04 mm。风力大,全年平均风速11.7 m/s,太阳辐射量较强,年均5.066 MJ/m2[14];苔原带物理风化强烈,地表多为疏松的火山岩风化物和火山灰等喷发物,流水侵蚀严重;主要发育有石质苔原土及苔原土、灌丛苔原土三类土壤[15]。研究区植被类型为苔原植被,主要植物有牛皮杜鹃(Rhododendron chrysanthum)、宽叶仙女木(Dryas octopetala var. asiatica)、笃斯越橘(Vaccinium uliginosum)、珠芽蓼(Polygonum viviparum)、长圆叶柳(Salix rotundifolia)、长白苔草(Carex siderosticia)、大白花地榆(Sanguisorba stipulata)、 雪白委陵菜(Potentilla nivea)、毛蕊老鹳草(Geranium platyanthum)、 高山红景天(Rhodiola cretinii)等[16]。与低温、强风的环境相适应,本地植物,如牛皮杜鹃、宽叶仙女木等,多匍匐生长,且生长缓慢;植物叶片多革质或背面密生绒毛[17]。
2 研究材料与方法 2.1 研究材料本文所用影像数据为Landsat MSS 1983年9月22日(空间分辨率60 m),以及Landsat ETM+1999年9月2日,2002年8月25日,2006年9月5日和2008年8月25日(空间分辨率均为30 m)共5期影像,选取的影像云量均为0%,而且时相相差很小,具有可对比性。影像预处理包括Landsat ETM+影像去条带,所选影像条带号116,行列号31,影像中心为苔原带,只在边缘处有少量条带,因此受条带影响很小;并进行几何校正,辐射校正等预处理工作。影像数据归一化植被指数(NDVI)的计算通过ENVI 4.7软件实现。气象数据采用天池站(距研究区6.2 km)对应TM影像年份的8—9月份数据。温度取此两个月的平均值,降水取此两个月降水总和。
野外实验针对目前小叶章入侵斑块进行植物群落调查,分析说明小叶章侵入现状及斑块特征,同时测度小叶章侵入对于群落生物多样性的影响;并进行土壤取样,分析小叶章侵入后对苔原带土壤环境的影响。
2.2 研究方法2012年6—9月份期间,在长白山苔原带西坡海拔2000—2500 m范围内,对可见的所有小叶章斑块进行GPS定位,跟踪记录斑块的数量、面积、周长等信息。同时,对样地内小叶章斑块进行群落调查,统计样方内植物的种类、株数、盖度、高度等信息。并根据小叶章侵入现状,以小叶章盖度对侵入斑块进行了分类:小叶章盖度>70%的斑块为重度侵入斑块、小叶章盖度70%—30%的斑块为中度侵入斑块、小叶章盖度<30%的斑块为轻度侵入斑块[18, 19, 20]。同时选择相邻的环境条件相似的本地牛皮杜鹃群落作为对照区。本研究利用景观分析软件Fragstat 3.4对小叶章斑块进行了斑块重要值,斑块数量,斑块平均面积及最大斑块面积等主要指标的分析。并采用4种α多样性指数测度群落物种的多样性,以此反映小叶章入侵对植物群落的影响:Shannon-Wiener多样性指数、Pielou均匀性指数、Margalef′s多样性指数、Richness丰富度指数。不同侵入强度下多样性指数的差异性检验采用单因素方差分析Tukey HSD检验。
(1)应用ASD Field-SpectralTM便携式地物光谱仪进行光谱采样
测量光谱范围350—2500 nm,在350—1000 nm 抽样间隔为1. 4 nm,光谱分辨率为3 nm; 在1000—2500 nm 抽样间隔为2 nm,光谱分辨率为10 nm。采样时间为2012年8月25日—9月5日,对应本研究采用的Landsat TM影像时间(8月底至9月底)。采样在晴朗无云的天气条件下进行,于10:00至14:00测定光谱。每20 min进行1次白板优化。光谱仪探头全视场角为25°,测定光谱时探头垂直向下,距植物群落冠层的高度为1. 5 m。3种植物群落各测定10组共150个数据。获得数据后应用RS3及ENVI软件对数据进行平滑去噪、平均化等处理,最终得出牛皮杜鹃群落(原生植被)、过渡群落(中度侵入斑块及轻度侵入斑块)、小叶章群落(重度侵入斑块)3种典型群落的光谱曲线,并计算NDVI值,公式如下:
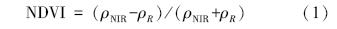
式中,ρNIR为波段775—900 nm(对应TM影像近红外波段)范围内的平均值,ρR为波段630—690 nm(对应TM影像红波段)范围内的平均值。
定量区分3种典型植物光谱红边特征的红边参数选择(1)红边幅值Dr,即红边(680—760 nm)内最大的一阶微分值;(2)红边位置λr,即最大一阶微分值对应的波长;(3)红边面积SDr,即红边内一阶微分值的总和。
(2)对基质和三类斑块进行土壤取样。苔原带土层较薄,因此本研究对0—5 cm土层土壤进行取样,并保证3次重复。采用Hobo土壤温水自动记录仪测定3种植物群落的土壤含水量变化。土壤性质的测定主要包括pH值、全氮、速效氮,以及腐殖质(包括胡敏酸和富里酸)等几个主要方面。土壤性质差异性检验采用单因素方差分析Tukey HSD检验。
3 结果 3.1 小叶章侵入现状表 1列出了5个海拔范围内的小叶章斑块分布现状。可以看出小叶章在低海拔处的斑块数量虽小,但面积较大;高海拔斑块数量多,但面积往往较小。斑块重要值体现了斑块对景观保持连通的重要性,斑块越大,对景观连通性意义越大。本文以100 m景观距离阈值计算斑块重要值。从变化看,小叶章斑块重要值沿海拔逐渐下降,结合入侵程度,可以推断低海拔处的小叶章斑块经过多年的扩张之后已连接成片,而高海拔处的斑块正处于扩张的阶段。低海拔处景观与小叶章斑块分布有着紧密的联系,高海拔处景观受小叶章斑块影响还较小。
| 海拔/m Elevation | 斑块重要值 Importance of patches | 斑块数量 Patch number | 斑块平均面积/ m2 Mean area of patches | 最大斑块面积/ m2 The largest patch area | 侵入程度 Invasive level |
| 2040—2080 | 0.184 | 4 | 2148 | 3509 | 重度 |
| 2100—2140 | 0.115 | 2 | 1299 | 1847 | 重度 |
| 2140—2180 | 0.101 | 8 | 537 | 1737 | 重度 |
| 2180—2220 | 0.089 | 5 | 318 | 411 | 重度 |
| 2220—2260 | 0.088 | 12 | 490 | 1551 | 中度、轻度 |
小叶章为禾本科草本植物,而牛皮杜鹃属常绿灌木。根据观察,8月底的长白山苔原带已进入秋季,小叶章叶片已明显变成黄色,而牛皮杜鹃依然保持着绿色。光谱曲线的差异证明了这一点。从实测光谱曲线来看,3种典型植物群落的光谱值存在着较大差异(图 1)。特别是在能够反映植物叶绿素浓度、生物量的红边波段(680—760 nm)上。红边幅值Dr由牛皮杜鹃群落向小叶章群落逐渐减小,红边面积SDr也随着不断减小,同时红边位置λr向短波方向移动。说明进入秋季,小叶章叶片叶绿素较之牛皮杜鹃叶片迅速下降,产生了显著的光谱差异。因此,通过光谱曲线反演的植被指数能够区分入侵植物群落与本地植物群落。根据计算,3种典型植物群落的NDVI值表现为小叶章群落<过渡群落<牛皮杜鹃群落。可见,NDVI的变化能够反映小叶章侵入牛皮杜鹃群落的过程。

|
| 图 1 牛皮杜鹃群落 (原生植被) 、过渡群落 (中度侵入斑块及轻度侵入斑块) 、小叶章群落 (重度侵入斑块) 3种典型群落的光谱曲线 Fig. 1 Spectrum curves of native plant community (R. chrysanthum community) ,transition plant community (invasive patches at low and medium invasive level) and D. angustifolia patches (invasive patches at high invasive level) λr为红边位置represents the red edge;Dr为红边幅值represents the amplitude of the red edge;SDr为红边面积represents the red edge area |
根据图 2可见,1983—2008年的25年间,经过GPS定位的小叶章斑块内部NDVI呈现出显著的减小趋势。自1999年开始,斑块的NDVI值已经开始变小,特别是在海拔2 060 m附近的两个较大斑块以及海拔2240 m处的几个较小的斑块。NDVI值大幅度的减小发生在21世纪,自2002年开始,所有斑块的NDVI值均低于1999年斑块NDVI的最小值。至2008年,斑块的NDVI值仍然表现为不断下降的趋势。联系影像对应时间段的温度水分条件(图 3),可以看出这5a长白山苔原带的水热配置条件是逐渐改善的,植物群落的长势应更加良好,从而呈现更高的绿度值,即NDVI值。然而1983年获取的影像时间为9月22日,与其他影像获取时间(8月25日—9月5日)相比较,其已明显处于深秋季节,且水热配置较差,温度偏高,降水偏少,却具备较高的NDVI值。这显然说明斑块内原有植物应为具备较高绿度值的牛皮杜鹃群落。然而伴随着小叶章的侵入,植物群落发生了明显的变化,主要表现为绿度值的不断减小。结合两种植物群落光谱值差异,可以看出,80年代初,小叶章还没有形成大规模的侵入,小叶章侵入牛皮杜鹃群落于1999年已经开始,并通过种间竞争逐渐获得了群落的主导地位。2000年后,侵入的小叶章逐渐替代牛皮杜鹃,形成以自身为主导的优势植物群落。

|
| 图 2 小叶章斑块NDVI多年变化图 Fig. 2 Changes in NDVI within the D. angustifolia patches 不同字母表示具有显著性差异(P<0.05) |

|
| 图 3 对应TM影像时间段 (8—9月份) 温度水分条件变化 Fig. 3 Variations of temperature and precipitation of August to September corresponding to the Landsat TM images |
牛皮杜鹃群落为苔原带本地植物群落,其主要受到小叶章的侵入影响。从表 2可以看出,伴随着小叶章的侵入,植物群落结构发生了显著的变化。本地植物群落和轻度侵入群落的优势种是牛皮杜鹃、笃斯越桔等灌木,草本植物很少;中度侵入强度群落的优势种是牛皮杜鹃和小叶章等植物;而重度侵入群落的优势种是小叶章、大白花地榆等草本植物。可以看出,伴随着小叶章的侵入,灌木数量明显减少,草本植物逐渐增多。相对应的,不同侵入强度群落植物的生活型也相应改变。另外,群落的高度也伴随着小叶章的侵入逐渐升高,这与草本植物的增多有关。
| 植物名称 Species | 高度/cm Height | 功能型 Functional type | 生活型 Life form | 对照 Control | 轻度入侵斑块 Invasive patches with low level | 中度入侵斑块 Invasive patches with medium level | 重度入侵斑块 Invasive patches with high level |
| Control为对照群落; +表示该物种存在于该植物群落 | |||||||
| 雪白委陵菜 Potentilla nivea | 5—25 | 多年生草本 | 地面芽植物 | + | |||
| 毛蕊老鹳草 Geranium platyanthum | 30—80 | 多年生草本 | 地面芽植物 | + | |||
| 高山乌头 Aconitum monanthum | 20—40 | 多年生草本 | 地下芽植物 | + | |||
| 藜芦 Veratrum nigrum L. | 50—100cm | 多年生草本 | 地下芽植物 | + | |||
| 大白花地榆 Sanguisorba stipulata | 50—80 | 多年生草本 | 地面芽植物 | + | + | ||
| 长白山橐吾 Ligularia jamesii | 30—60 | 多年生草本 | 地下芽植物 | + | + | ||
| 珠芽蓼 Polygonum viviparum L. | 5—8 | 多年生草本 | 地下芽植物 | + | + | ||
| 小叶章 Deyeuxia angustifolia | 60—90 | 多年生草本 | 地面芽植物 | + | + | + | |
| 长白苔草 Carex peiktusanii | 10—15 | 多年生草本 | 地上芽植物 | + | + | + | |
| 高山红景天 Rhodiola cretinii | 6—20 | 多年生草本 | 地面芽植物 | + | + | + | + |
| 牛皮杜鹃 Rhododendronchrysanthum | 10—25 | 常绿灌木 | 中位芽植物 | + | + | + | |
| 笃斯越桔 Vaccinium uliginosum | 10—15 | 落叶灌木 | 中位芽植物 | + | + | ||
| 圆叶柳 Salix rotundifolia | 2—3 | 矮灌木 | 地上芽植物 | + | |||
从图 4可以看出,小叶章入侵后对本地植物群落产生了很大的影响。从Shannon-Wiener index指数变化来看,从本地牛皮杜鹃群落和轻度入侵斑块到中度、重度侵入群落的多样性随海拔上升减小;中度入侵斑块呈现波动减小的趋势;而重度侵入斑块则表现出波动增加后减小的趋势。总体上看,重度入侵斑块的多样性要大于轻度入侵斑块。说明伴随着小叶章的侵入,群落物种数量在不断增加。物种丰富度指数(Richness index)的变化证明了这一点,且Richness指数沿海拔梯度呈现减小的趋势,说明海拔对于物种分布有着一定的限制作用,即对于小叶章入侵有着一定的抵御作用。伴随着小叶章的侵入,群落物种数量明显升高。重度入侵斑块的波动性说明这种入侵使得植物群落结构处于不稳定的状态。丰富度指数(Margalef′s index)与Shannon-Wiener指数变化相似,其在海拔2151 m以下位置,轻度入侵斑块丰富度要小于重度入侵斑块,在海拔2 151 m以上两者近似;总体而言,中度入侵斑块的Margalef′s丰富度指数要大于其他两种群落。证实了小叶章入侵后,植物群落的物种数量在不断增加。均匀度指数(Evenness index)表现为重度侵入斑块大于中度和轻度侵入斑块,且随海拔上升减小;结合Shannon-Wiener指数和Richness指数的变化来看,伴随着小叶章的侵入,群落物种数量增加,小叶章在群落中的优势主导地位随海拔上升而不断上升;在高海拔处易形成以自身为主导的优势植物群落。

|
| 图 4 不同侵入强度下植物群落的α多样性随海拔的变化 Fig. 4 Changes in α biodiversity of the plant communities at different invasive levels Control为对照群落,即本地牛皮杜鹃群落; 不同字母表示具有显著性差异(P<0.05); R. chrysanthumcommunity is considered as the control plot,which is also the native communities. Different letters mean significantly different(P<0.05) |
小叶章入侵对于土壤性质的影响较大。首先,重度入侵斑块的土壤含水量高于中度入侵斑块及本地牛皮杜鹃群落下的土壤(图 5)。这说明伴随着小叶章的入侵,土壤持水能力逐渐升高。这直接影响了土壤的腐殖质分解过程。从土壤腐殖质含量来看(表 3),轻度入侵斑块较之重度入侵斑块腐殖质含量高,中度入侵群落则表现出最高的腐殖质含量,3种斑块的腐殖质含量均低于本地牛皮杜鹃群落。说明轻度入侵斑块保持了原牛皮杜鹃群落下灌丛苔原土的特性,而伴随着小叶章的侵入,土壤中腐殖质含量呈现下降的趋势。富里酸是土壤腐殖质的重要组成部分,且在3种土壤中均大于胡敏酸含量。可以看出伴随着小叶章的侵入,土壤富里酸是不断下降的。胡敏酸则表现为本地牛皮杜鹃群落以及轻度入侵斑块大于重度入侵斑块。小叶章入侵导致土壤含水量的增加,而富里酸的各个价态都能很好的溶解在水中,胡敏酸则刚好相反。因此,随着小叶章的侵入,土壤富里酸的含量不断上升。3种斑块的土壤中全氮、速效氮的差异并不是很大。伴随着小叶章入侵强度的增加,土壤氮的含量不断减小,而速效氮的含量有略微的增加。土壤C/N比率都较大,表明土壤变化滞后于植被变化。且伴随小叶章的入侵,C/N比率不断下降,说明小叶章入侵后草本植物对土壤贡献提升,土壤草甸化。另外,各类斑块的土壤均呈酸性反应。

| 植物群落类型 Plant community types | 腐殖质 Soil humus/ (g/kg) | 胡敏酸 Humic acid/ (g/kg) | 富里酸 Fulvic acid | 碳氮比 C/N ratio | 全氮 Total nitrogen/ (g/kg) | 速效氮 Available nitrogen/ (g/kg) | pH |
| 土壤深度均为5 cm,数值代表平均值±标准差; 对照为本地牛皮杜鹃群落; 不同字母表示具有显著性差异 | |||||||
| 对照Control | 97.62±1.052a | 36.88±4.91a | 55.16±2.011a | 20.44±4.156a | 11.58±2.301a | 1.22±0.13a | 4.38±0.271a |
| 轻度入侵斑块 Invasive patches with low level | 96.12±2.016a | 35.24±7.21ab | 54.10±9.24b | 20.56±3.81a | 11.00±2.48a | 1.29±0.09a | 4.52±0.17a |
| 中度入侵斑块 Invasive patches with medium level | 92.78±4.149b | 40.92±10.51a | 39.58±12.28ac | 18.36±2.81ab | 10.42±2.69ab | 2.38±0.11a | 4.63±0.19a |
| 重度入侵斑块 Invasive patches with high level | 88.42±2.811ac | 31.55±7.58b | 33.84±11.85b | 16.49±3.84b | 8.45±2.10b | 2.93±0.11a | 4.82±0.11b |
(1)目前小叶章已经成功侵入到长白山西坡苔原带区域。低海拔处的斑块数量小但面积较大;高海拔处小叶章斑块面积较小但数量较多。低海拔处的小叶章斑块经过多年的扩张生长之后已连接成片而高海拔处的斑块正处于扩张的初期阶段。低海拔处景观与小叶章斑块分布有着紧密的联系,高海拔处景观受小叶章斑块影响还较小。
(2)通过影像分析来获取长时间序列的草地植被变化存在着较大困难。目前,较多研究立足于高光谱影像数据的分析,以期获得入侵植物的分布信息,然而对于入侵变化过程较难给出确定的研究结果,这主要受制于早期影像数据的光谱分辨率较低 的限制。另一方面,长时间序列的影像数据目前主要以Landsat TM系列数据为主,此类数据空间及光谱分辨率较低,在区分森林植被和草地植被研究中具有一定的优势,然而区分草地植被中的不同植物群落则效果不佳。利用季相差异是区分入侵植物和本地植物的有效技术方法[9]。本研究依据小叶章与牛皮杜鹃的光谱差异及其反演的植被指数,探讨小叶章侵入苔原带的过程。认为小叶章上侵开始于20世纪80年代末,进入21世纪后逐渐形成稳定的以小叶章为优势物种的植物群落结构。结合长白山西坡植被变化的历史过程,1986年长白山西坡经历了一次较强的台风,在岳桦林带形成了大量的林窗,目前林窗中的小叶章生长旺盛。这很可能与其侵入苔原带有着紧密的联系。同时,这又进一步印证了NDVI值变化的分析结果。
(3)小叶章侵入苔原带对植物群落的影响较大,主要表现为植物群落多样性的升高和物种数量的增加。伴随着小叶章的侵入,灌木数量明显减少,草本植物逐渐增多,其改变了牛皮杜鹃群落的群落结构,并通过竞争逐渐替代牛皮杜鹃成为了群落的主导优势物种,体现了较强的竞争力。伴随着小叶章的侵入,更多的草本植物出现了新的群落中;群落结构的改变影响了土壤的理化性质,小叶章入侵后土壤腐殖质含量和全氮含量下降,但土壤养分的高效利用又进一步推动了小叶章的生长。可见,土壤的变化将进一步影响植物群落结构的改变。
(4)长白山西坡小叶章侵入苔原带现象在我国高山植被研究中是十分独特的,从植物多样性分析和土壤理化性质变化来看,已经对苔原带造成了严重的生态后果。小叶章的进一步发展必将威胁牛皮杜鹃等苔原植物的生存发展,作为我国温带地区仅有的高山苔原景观也必将因其发生而变化。
小叶章侵入苔原带尚需进一步的实验研究来获悉其机制。通常植被的变化与生态系统干扰息息相关,1986年的台风干扰使得小叶章在林窗中得以大量繁殖生长,这无疑为侵入苔原带提供了大量的种源。同时气候变暖的影响也不可忽视。Lenoir认为气候变暖是导致低海拔物种分布范围上移的驱动因素[22];Breshears认为气候变暖将导致高山不同植被带位置整体上移[23]。长白山苔原带气候目前正逐步变暖[14],小叶章侵入苔原带又或许是对于气候变暖的一种响应。然而北坡尚未发现小叶章侵入苔原带的现象,两个坡向的气候条件类似,植被变化却有着如此大的差异,说明两个坡向的植被演替进程存在着差异。由此,理解小叶章上侵应从台风干扰、气候变化、植被演替等多个方面综合研究,并考虑到这些要素之间的耦合作用,这尚需长时间序列的野外观测与调查数据的支持,从而更为深入的理解温带高山地区植被垂直带变动的机理,并进一步解释长白山植被分布的特殊性。
| [1] | Wearne L J, Morgan J W. Shrub invasion into subalpine vegetation: implications for restoration of the native ecosystem. Plant Ecology, 2006, 183(2): 361-376. |
| [2] | Krannitz P, Kesting S.Impacts of climate change on the plant communities of alpine ecosystems//Taylo E, Taylor B, eds. Responding to global climate change in British Columbia and Yukon. Ministry of Environment,Lands and Parks, Victoria, B.C, 1997. |
| [3] | Tsuyuzaki S. Wetland development in early stages of volcanic succession. Journal of Vegetation Science, 1997, 8(3): 353-360. |
| [4] | Shi P L, Li W H. Boundary form effects of timberline ecotone on colonization of woody plants and timberline dynamics in Changbai Mountain. Acta Ecologica Sinica, 2000, 20(4): 389-398. |
| [5] | Qian J J. Vertical Plant Checklist of Changbai Mountain. Changchun: Northeast Normal University Press, 1980: 90-91. |
| [6] | Qian H. Alpine tundra vegetation on the Changbai mountain //Forest Ecological Research vol. 6. Beijing: China Forestry Publishing House, 1992: 72-96. |
| [7] | Zong S W, Xu J W, Wu Z F. Investigation and mechanism analysis on the invasion of Deyeuxia. angustifolia to tundra zone in western slope of Changbai Mountain. Journal of Mountain Science, 2013, 31(4): 448-455. |
| [8] | Bradley B A, Mustard J F. Characterizing the landscape dynamics of an invasive plant and risk of invasion using remote sensing. Ecological Applications, 2006, 16(3): 1132-1147. |
| [9] | Walsh S J, Mccleary A L, Mena C F, Shao Y, Tuttle J P, González A, Atkinson R. QuickBird and Hyperion data analysis of an invasive plant species in the Galapagos Islands of Ecuador: Implications for control and land use management. Remote Sensing of Environment, 2008, 112(5): 1927-1941. |
| [10] | Gao Z G, Zhang L Q. Measuring and analyzing of the multi seasonal spectral characteristics for saltmarsh vegetation in Shanghai. Acta Ecologica Sinica, 2006, 26(3): 793-800. |
| [11] | Wolf J J, Beatty S W, Carey G. Invasion by sweet clover (Melilotus) in montane grasslands, Rocky Mountain National Park. Annals of the Association of American Geographers, 2003, 93(3): 531-543. |
| [12] | Hejda M, Pyšek P, Jarošík V. Impact of invasive plants on the species richness, diversity and composition of invaded communities. Journal of Ecology, 2009, 97(3): 393-403. |
| [13] | Flory S L, Clay K. Non-native grass invasion alters native plant composition in experimental communities. Biological Invasions, 2010, 12(5): 1285-1294. |
| [14] | Zong S W, Wu Z F, Du H B. Variation characteristics of climate change in Changbai Alpine tundra during the growing season in recent 52 years. Arid Zone Research, 2013, 30(1): 41-49. |
| [15] | Huang X C, Li C H. An analysis on the ecology of alpine tundra landscape of Changbai mountains. Acta Geographica Sinica, 1984, 39(3): 285-297. |
| [16] | Qian J J, Zhang W Z. A brief report on the reseatch of the Changbaishan alpine. Journal of Northeast Normal University:Natural Science Edition, 1980, (1): 51-67. |
| [17] | Pauchard A, Kueffer C, Dietz H, Daehler C C, Alexander J, Edwards P J, Arévalo J R, Cavieres L A, Guisan A, Haider S. Ain't no mountain high enough: plant invasions reaching new elevations. Frontiers in Ecology and the Environment, 2009, 7(9): 479-486. |
| [18] | Zhang T R, Huangfu C H, Bai X M, Yang D L, Li G, Lai X, Zhao J N. Effects of Flaveria bidentis invasion on soil nutrient contents and enzyme activities. Chinese Journal of Ecology, 2010, 29(7): 1353-1358. |
| [19] | Yu X J, Yu D, Lu Z J, Ma K P. A possible plant invasion mechanism-invader restricted native plant by altering the soil microbes community. Chinese Science Bulletin, 2005, 50(9): 896-903. |
| [20] | Guo X H, Ren M X, Ding J Q, Zheng J M. Plant species diversity and its seasonal dynamics in woodland invaded by Solidago Canadensis (Asteraceae). Plant Science Journal, 2011, 29(2): 149-155. |
| [21] | Zong S W, Wu Z F, Xu J W, Qin L J, Tao Y, Du H B, Meng X J, Li M, Wang L, Dong D J. Invasion of Deyeuxia angustifolia in the Western Slope of Alpine Tundra, Changbai Mountain. Biological Invasions, 2013 (under review). |
| [22] | Lenoir J, Gegout J C, Marquet P A, de Ruffray P, Brisse H. A significant upward shift in plant species optimum elevation during the 20th century. Science, 2008, 320(5884): 1768-1771. |
| [23] | Breshears D D, Huxman T E, Adams H D, Zou C B, Davison J E. Vegetation synchronously leans upslope as climate warms. Proceedings of the National Academy of Sciences, 2008, 105(33): 11591-11592. |
| [4] | 石培礼, 李文华. 长白山林线交错带形状与木本植物向苔原侵展和林线动态的关系. 生态学报, 2000, 20(4): 389-398. |
| [5] | 钱家驹. 长白山植物垂直分布名录. 长春: 东北师范大学学报, 1980: 90-91. |
| [6] | 钱宏. 长白山高山冻原植被 // 森林生态系统研究(第六卷). 北京: 中国林业出版社, 1992: 72-96. |
| [7] | 宗盛伟, 许嘉巍, 吴正方. 长白山西坡小叶章侵入苔原带调查与机理分析. 山地学报, 2013, 31(4): 448-455. |
| [10] | 高占国, 张利权. 上海盐沼植被的多季相地面光谱测量与分析. 生态学报, 2006, 26(3): 793-800. |
| [14] | 宗盛伟, 吴正方, 杜海波. 近52a长白山苔原带生长季气候变化特征. 干旱区研究, 2013, 30(1): 41-49. |
| [15] | 黄锡畴, 李崇皜. 长白山高山苔原的景观生态分析. 地理学报, 1984, 39(3): 285-297. |
| [16] | 钱家驹, 张文仲. 长白山高山冻原植物的调查研究简报. 东北师大学报: 自然科学版, 1980,(1): 51-67. |
| [18] | 张天瑞, 皇甫超河, 白小明, 杨殿林, 李刚, 赖欣, 赵建宁. 黄顶菊入侵对土壤养分和酶活性的影响. 生态学杂志, 2010, 29(7): 1353-1358. |
| [19] | 于兴军, 于丹, 卢志军, 马克平. 一个可能的植物入侵机制-入侵种通过改变入侵地土壤微生物群落影响本地种的生长. 科学通报, 2005, 50(9): 896-903. |
| [20] | 郭晓辉, 任明迅, 丁建清, 郑景明. 加拿大一枝黄花入侵林地对植物多样性的影响及其季节变化. 植物科学学报, 2011, 29(2): 149-155. |