 2014, Vol. 34
2014, Vol. 34文章信息
- 潘帅, 于澎涛, 王彦辉, 王占印, 袁彩霞, 余治家, 胡永强, 熊伟, 徐丽宏
- PAN Shuai, YU Pengtao, WANG Yanhui, WANG Zhanyin, YUAN Caixia, YU Zhijia, HU Yongqiang, XIONG Wei, XU Lihong
- 六盘山森林植被碳密度空间分布特征及其成因
- Spatial distribution of carbon density for forest vegetation and the influencing factors in Liupan Mountains of Ningxia, NW China
- 生态学报, 2014, 34(22): 6666-6677
- Acta Ecologica Sinica, 2014, 34(22): 6666-6677
- http://dx.doi.org/10.5846/stxb201302230291
-
文章历史
- 收稿日期:2013-2-23
- 网络出版日期: 2014-3-14






2. 江西省林业勘察设计院, 南昌 330046;
3. 宁夏六盘山林业局, 宁夏泾源 756400
2. Jiangxi Forest Inventory & Planning Institute, Nanchang Jiangxi 330046, China;
3. Ningxia Liupanshan Bureau of Forestry, Jingyuan Ningxia 756400, China
我国西北生态环境脆弱,水土流失严重,增加森林植被是改善环境的重要途径之一[1]。为此,几十年来连续实施三北防护林、天然林保护、退耕还林等多项重点林业工程,使森林植被大幅增加[2]。近些年来,由于增大森林固碳量被看作是应对气候变化的有效途径之一[3],导致该区造林活动进一步加强。如胡锦涛主席在2009年联合国气候变化峰会上郑重承诺:2020年中国森林面积将比2005年新增4000万 hm2。为实现此目标,西北地区森林需在2000年900万 hm2的基础上增加到2020年的2095万 hm2[4]。不少地方政府已采取措施,如陕西仅2010年就造林26.7万 hm2[5]。
造林活动是固碳和耗水的一种利弊权衡[6]。同草地和灌丛相比,森林要消耗更多的水,导致径流减少,如全球湿润区(年降水量>800 mm)造林导致年径流量平均减少166 mm,这占年径流量的38%[7]。在干旱地区造林影响更大,如黄土高原造林平均减少年径流量23 mm,这相当于年径流量的一半[8],这会进一步加剧区域水资源短缺的状况,给区域发展带来极大的限制[9]。
事实上,森林的固碳功能及其对径流的减少作用不仅与森林面积大小、林分结构有关,也与森林分布的位置密切相关,如在六盘山香水河小流域,10%的流域面积造林后,因立地差异导致的年径流减少量在3.5—19.3 mm之间变动[10]。因此,了解森林碳储量及其时空分布对于合理造林、平衡碳水矛盾十分重要。
六盘山是我国西北黄土高原的重要林区和水源地,泾河等多条河流均源于此,年产径流2亿 m3[11],对维护周边和下游的生态环境及水资源安全具重要作用[12]。通过几十年的封山育林和人工造林等,森林覆盖率已由1975年的27.8%[11]增加至2005年的52.8%[13]。然而目前有关该区森林植被固碳的研究仅讨论了生长状况较好的典型森林样地的植被碳密度,如吴建国等[14]研究了六盘山北部农牧交错带的主要土地覆被类型的植被碳密度(t/hm2),表明次生林植被碳密度为21.4—33.5,人工林植被碳密度为18.4—52.1;刘延惠等[15]研究表明,六盘山南段香水河小流域典型森林样地上的植被碳密度(t/hm2)在22.67—56.69之间。然而六盘山地区的森林植被碳密度及其空间分布规律并不清楚,这限制了该区森林固碳耗水成本的估算、林-水-碳的综合管理以及多功能森林植被的恢复等。
六盘山的森林集中分布在六盘山自然保护区内,因此本文根据该保护区2005年的森林资源一类清查数据,应用文献中植被生物量回归(经验)模型和碳含量数据资料,计算了森林植被碳密度,并分析了其与森林的分布环境、林分结构特征的关系,旨在为定量确定区域森林固碳耗水成本、指导林-水-碳的综合管理提供依据。
1 研究区概况宁夏六盘山位于黄土高原西部,由平行排列的东南-西北走向的两列山脉组成,其中包含主峰“米缸山”的西脉为主脉,海拔1900—2942 m,多在2500 m以上;东脉海拔介于1657—2420 m,多低于2300 m。
六盘山作为东亚季风的西北边缘,受东南季风的影响由东南向西北逐渐减弱,气候亦由温带半湿润区过渡到温带半干旱区,年降水量在六盘山东南部二龙河(海拔2150 m,820 mm)经西峡(海拔2050 m,770 mm)、和尚铺(海拔2040 m,671 mm)至西北部的固原(海拔1752 m,478 mm)一线逐渐减少[11]。此外,六盘山气候的垂直分异也十分显著,如在米缸山东坡海拔2000 m处,年均气温5.4℃,年均降水770 mm,而山顶(海拔2942 m)年均气温0.6℃,年均降水690 mm,其温度垂直递减率为0.4—0.6℃/100 m[11],在海拔2640 m以下,雨季(6—10月)降水量垂直递增率为13.7 mm/100 m,而海拔2640 m以上的降水量垂直递减率为14.5 mm/100 m[16]。
六盘山自然保护区设立于1982年,位于六盘山南端(35°15′—35°55′N,106°09′—106°30′E),包括14个林场(图 1),面积904 km2,是六盘山森林分布的核心区。六盘山的森林以天然次生林为主(表 1),占保护区森林总面积的73.6%,主要树种有辽东栎(Quercus liaotungernsis)、山杨(Populus davidiana)、桦木(Betula spp.)、柳树(Salix cathayana)、华山松 (Pinus armandii),、榆树(Ulmus pumila)、椴树(Tilia chinensis)。 人工林主要为华北落叶松(Larix principris-upprechtii)林,占人工林总面积的90%,还有少量的云杉(Picea asperata)和油松(Pinus tabulaeformis)林。保护区现有森林多为中幼龄林,林龄平均30 a,以20—40 a为主;平均胸径11.8 cm,平均树高8.3 m。林分整体比较稀疏,有50%林分的郁闭度低于0.4;平均密度为827株/hm2,65%的林分低于1000株/hm2。

|
| 图 1 研究区位置及调查样地分布 Fig. 1 Spatial distribution of inventory plots and the location of study area A:华山松林 Pinus armandii forest;B:辽东栎林 Quercus liaotungernsis forest;C:桦木林Betula spp forest.;D:山杨林Populus davidiana forest;E:柳树林Salix cathayana forest;F:其它硬阔叶林Other hardwoods forest;G:其它软阔叶林Other softwoods forest;H:华北落叶松人工林Larix principris-upprechtii plantation;I其它针叶林 Other conifer forest |
| 起源 Origin | 森林类型 Forest types | 面积 Area/hm2 | 占森林总面积的比例/%
Proportion in total forest area |
| 天然次生林 | 辽东栎林 | 6901 | 18.6 |
| Natural | 桦木林 | 4722 | 12.8 |
| secondary | 山杨林 | 3995 | 10.8 |
| forest | 柳树林 | 6901 | 18.6 |
| 华山松林 | 726 | 2.0 | |
| 其它硬阔叶林 | 1452 | 3.9 | |
| 其它软阔叶林 | 2542 | 6.8 | |
| 人工林 | 华北落叶松人工林 | 8717 | 23.5 |
| Plantation | 油松人工林 | 726 | 3.0 |
| 云杉人工林 | 363 | 1.0 |
为比较六盘山森林植被碳密度的空间差异,依据年降水量空间分布,并结合林场分布边界,将保护区划分为Ⅰ、Ⅱ、Ⅲ 3个亚区(图 1)。其中,Ⅰ区年均降水量大于700 mm,包括二龙河、西峡、龙潭3个林场;Ⅱ区年均降水量为600—700 mm,包括东山坡、和尚铺、秋千架、卧羊川4个林场;Ⅲ区年均降水量为500—600 mm,包括绿源、水沟、丰台、挂马沟4个林场。
2 材料与方法 2.1 数据来源本研究采用六盘山2005年森林资源一类清查数据。森林资源一类清查按照2 km×2 km的格网在整个研究区均匀布设面积为20 m×30 m的森林样地共115个(图 1),调查指标包括样地的地理位置、海拔高度 、坡向、坡位、坡度、树种组成、胸径、郁闭度、灌木层和草本层物种组成及覆盖度等特征。本文运用海拔高度、坡向、胸径、郁闭度、灌木层和草本层覆盖度数据计算各样地植被碳密度。
2.2 样地碳密度(CD)计算森林植被碳密度包括乔木层、灌木层和草本层的碳密度,各层碳密度由其生物量密度乘以含碳系数得到:
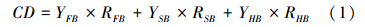
根据样地树木生长特征,用生物量模型计算单木总生物量,再计算样地内所有单木总生物量之和,得到乔木层生物量密度(YFB):

|
树种
Tree species | 树高-胸径关系
Relation between tree height and DBH | 资料来源
Cited from | 生物量模型
Biomass model | 参考文献
References |
| H为树高(m),W根为单木地下生物量(kg),W地上为单木地上生物量(kg) | ||||
| 桦木Betula spp. | H=2.8943+0.7992D-0.0149D2 | [17] | lnWj=0.8114ln(D2H)-1.8989 | [18] |
| 辽东栎Quercus liaotungernsis | H=1.0853+0.7877D-0.0139D2 | [17] | lnWj=0.9979ln(D2H)-3.5426 | [18] |
| 山杨Populus davidiana | H=2.5547+0.6486D-0.0099D2 | [17] | lnWj=0.9222ln(D2H)-2.8360 | [18] |
| 柳树Salix cathayana | H=3.388547+0.5155018D-0.00335713D2 | [17] | W根=0.0495502(D2H)^0.952453/3.85
W地上=0.0495502(D2H)^0.952453 Wj= W根+ W地上 | [19] |
| 其它硬阔Other hardwoods forest | H=4.1423+0.3336D-0.0038D2 | [17] | lnWj=0.9037ln(D2H)-2.5700 | [18] |
| 其它软阔Other softwoods forest | H=4.1423+0.3336D-0.0038D2 | [17] | W根=0.0495502(D2H)^0.952453/3.85
W地上=0.0495502(D2H)^0.952453 Wj= W根+ W地上 | [19] |
根据样地灌木层的总覆盖度(XSC,小数),计算样地灌木层生物量密度[15]:

根据样地的坡向(AP,°)和海拔高度(XE,m)以及草本层总覆盖度(XHC,小数),计算草本层生物量密度[15]:
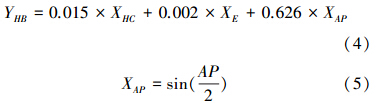
六盘山森林植被碳密度为0.67—120.63 t/hm2,平均为(26.17±21.2) t/hm2,碳密度在0—15、15—30、30—45 t/hm2范围内的林分占森林面积比例为24.3—31.1%;碳密度介于45—60、60—75 t/hm2之间的林分所占比例均为9.1%;只有0.8%的森林碳密度高于75 t/hm2,说明保护区森林植被碳密度以低碳密度林分为主,高碳密度林分较少。
天然次生林植被碳密度平均为(30.2±21.4) t/hm2,显著高于人工林的植被碳密度(P<0.05)(表 4)。在天然次生林中,各森林类型碳密度(t/hm2)平均值依次为山杨林(22.3±12.7)<桦木林(28.4±14.7)<柳树林(35.9±20.8)<辽东栎林(43.4±25.2),即由先锋群落至顶级群落,碳密度呈增加的趋势,但不同森林类型间差异并不显著(P>0.05)。人工林碳密度平均为(15.7±16.0) t/hm2,在人工林中,华北落叶松人工林的碳密度更低,平均仅(12.4±14.7) t/hm2(表 4),这与华北落叶松林龄较小、林分密度较低有关。
|
起源
Origin |
森林类型
Forest types | 样地数
Plot numbers | 总碳密度
Carbon density /(t/hm2) |
| ()外为平均值±标准差,()内为碳密度变化范围,a、b表示差异显著程度,同一列相同字母表示差异不显著((P>0.05)),不同字母表示差异显著(P<0.05) | |||
| 天然次生林 | 辽东栎林 | 19 | 43.4±25.2(15.7—120.6)a |
| Natural secondary | 柳树林 | 19 | 35.9±20.8(10.3—76.3) a |
| forest | 桦木林 | 13 | 28. 4±14.7(10.7—63.8) a |
| 山杨林 | 12 | 22.3±12.7(7.6—46.3) a | |
| 全部天然次生林 | 86 | 30.2±21.4(7.6—120.6) a | |
| 人工林 | 华北落叶松人工林 | 26 | 14.5±14.7(0.67—66.7)b |
| Plantation | 全部人工林 | 29 | 15.7±16.0(0.67—66.7)b |
乔木层为森林植被固碳的主体,其碳密度为0.14—119.7 t/hm2,占植被总碳密度的比例为8.86%—99.2%,平均(69.4±27.1)%,乔木层碳密度占总碳密度的比列高于50%的林分占保护区森林面积的77.3%,而且植被总碳密度越大,乔木层碳密度所占比例越大,在植被总碳密度介于0—15、15—30、30—45、45—60和大于60 t/hm2的林分中,乔木层碳密度所占总碳密度的比例依次为(43.1±25.0)%、(74.3±11.1)%、(87.6±6.6)%、(92.7±2.8)%、(95.4±3.1)%。由于乔木层是植被固碳的主体,因此不同森林类型间乔木层碳密度差异与总碳密度差异一致,表现为天然次生林显著高于人工林(P<0.05),其平均值分别为(25.4±16.5)和(12.5±16.0) t/hm2;在天然次生林中,桦木林和山杨林作为先锋群落,乔木层碳密度分别为(16.4±13.1)、(23.9±14.4) t/hm2,而顶级群落辽东栎和柳树林的乔木层碳密度分别(38.8±26.7)、(32.6±21.2) t/hm2(表 5),但不同天然次生林间乔木层碳密度的差异不显著(P>0.05)。
|
起源 Origin | 森林类型 Forest types | 碳密度Carbon density(t/hm2) | ||
| 乔木层Trees | 灌木层Shrub | 草本层Herb | ||
| ()外为平均值±标准差,()内为碳密度变化范围; a、b表示差异显著程度,同一列不同字母表示差异显著(P<0.05),相同字母表示差异不显著(P>0.05) | ||||
| 天然次生林 | 辽东栎林 | 38.8±26.7(9.3—119.7)a | 4.3±2.5(0.67—8.9) a | 0.22±0.12(0.04—0.48) a |
| Natural secondary forest | 柳树林 | 32.6±21.2(6.2—71.7) a | 4.1±2.1(1.2—8.9) a | 0.52±0.16(0.26—0.91)b |
| 桦木林 | 23.9±14.4(2.7—55.3) a | 4.6±2.5(1.2—8.4) a | 0.57±0.17(0.27—0.9) b | |
| 山杨林 | 16.4±13.1(1.1—40.4) a | 5.0±2.5(1.2—8.4) a | 0.38±0.26(0.10—0.87)b | |
| 全部天然次生林 | 25.4±16.5(1.1—119.7) a | 4.4±2.3(0.67—8.9) a | 0.42±0.22(0.04—0.91) b | |
| 人工林 | 华北落叶松人工林 | 11.3±15.3(0.14—69.1)b | 2.65±2.2(0.15—6.9) b | 0.58±0.22(0.08—0.96) b |
| Plantation | 全部人工林 | 12.5±16.0(0.66—69.7)b | 2.62±2.1(0.15—6.9) b | 0.58±0.23(0.08—0.96) b |
灌木层碳密度为0.15—8.9 t/hm2,占植被总碳密度的比例为1.6—89.1%,平均25.6±24.3%,灌木层碳密度占总碳密度的比例大于50%林分仅占保护区森林面积17.3%,而且植被总碳密度越大,灌木层碳密度所占比例越低,在植被总碳密度介于0—15、15—30、30—45、45—60和大于60 t/hm2的林分中,灌木层碳密度所占比例依次为(43.6±28.1)%、(22.8±11.5)%、(11.2±6.8)%、(6.2±3.6)%、(4.3±3.6)%。灌木层碳密度表现为天然次生林显著高于人工林(P<0.05),其平均值分别为(4.4±2.3)、(2.65±2.1) t/hm2;不同类型的天然次生林灌木层碳密度接近,辽东栎林、柳树林、桦木林、山杨林依次为(4.3±2.5)、(4.1±2.1)、(4.6±2.5)、(5.0±2.5) t/hm2(表 5)。
草本层对森林植被固碳功能的贡献最低,介于0.01—0.91 t/hm2之间,占植被总碳密度的比例为0.12—32.3%,平均仅(4.7±9.8)%,草本层碳密度占总碳密度的比例以低于5%的林分为主,占保护区森林面积的76.7%,随着植被总碳密度增加,草本层碳密度所占比例也呈下降趋势,在植被总碳密度介于0—15、15—30、30—45、45—60和大于60 t/hm2的林分中,草本层碳密度所占比例依次为(9.7±14.3)%、(2.4±1.3)%、(1.27±0.07)%、(0.45±0.41)%、(0.53±0.23)%。天然林的草本层碳密度为(0.42±0.17) t/hm2、小于人工林(0.58±0.22) t/hm2,但差异不显著(P<0.05);在天然次生林中,辽东栎林、柳树林、桦木林、山杨林草本层碳密度依次为(0.22±0.12)、(0.52±0.16)、(0.57±0.17)、(0.38±0.26) t/hm2,辽东栎林显著低于其它森林类型(P<0.05)(表 5),这可能是因为与其它次生林相比,辽东栎林仅分布于阴坡、半阴坡、半阳坡,其中半阳坡仅占26.7%,而且郁闭度较大,平均为(0.5±0.21)(其它次生林为0.46±0.23),导致林下光照缺乏所致。
3.2 森林植被碳密度与林分特征的关系 3.2.1 碳密度随林龄的变化林木生长随林龄变化通常表现为典型的“S”型曲线,依据“S”曲线上两个曲率最大的点可将生长曲线划分为3个阶段,按时间顺序依次为缓慢期、速生期、平缓期[20]。栎林、桦林分的碳密度快速增长期为15—80 a,落叶松人工林碳密度的快速增长期为10—30 a[21]。在研究区96.3%的人工林的林龄介于10—30 a之间,93.3%的天然林的林龄介于15—80 a之间。由此来看,保护区绝大部分森林正处于速生期,所以植被的碳密度随林龄增加快速增加,其增长速率天然次生林为1.16 t hm-2 a-1,人工林为2.48 t hm-2 a-1(图 2)。研究区现有辽东栎林、桦木林、华北落叶松人工林平均碳密度为43.4、28.4、14.5 t/hm2,而它们的成熟林碳密度在全国的平均值依次为98.5、81.7、65.1 t/hm2[21],可见保护区现有森林碳密度远低于全国成熟林平均碳密度值,但是研究区33 a生华北落叶松人工林碳密度可达66.7 t/hm2,超过其成熟林在全国的平均值,40 a生辽东栎林和桦木林碳密度即可达72.0、63.8 t/hm2,已非常接近全国成熟林的平均值,这说明保护区林分碳密度能达到甚至超过全国森林碳密度的平均水平,固碳潜力较大。

|
| 图 2 碳密度随林龄的变化 Fig. 2 Variation of carbon density with forest stand age |
无论是天然次生林还是人工林,在各林分密度范围内,碳密度的变异较大,如林分密度为800—1200株/hm2时,天然次生林碳密度变化范围为13.3—76.3 t/hm2,人工林碳密度变化范围为9.8—32.8 t/hm2,这主要因为碳密度还同时受林龄、立地条件等多因素的共同作用。
林木生长对某一环境因子的外包线可以看作该环境因子作用下的最大生长潜力[22, 23]。依据碳密度随林分密度变化的外包线(图 3)来看,所有森林的植被碳密度均随林分密度增加而线性增大,但在林分密度大于1000株/hm2后不再增大,而是趋于一个极值,这表明碳密度存在一个林分密度阈值,低于这一阈值,碳密度随林分密度增加迅速增加,高于这一阈值,碳密度不再增加,此极值对天然林约为75.4 t/hm2,对人工林约为34.6 t/hm2。这种碳密度随林分密度变化的阈值现象在其它研究区也有发现,如在黄土高原方山县,采用集水造林措施营造的25 a生刺槐林林分密度在475—1600 株/hm2之间时,密度越大,蓄积量越大,高于这一范围,蓄积量随密度增加而减少[24];在长白山地区,28年生落叶松林分密度高于1190株/hm2时,随着林分密度变大,生物量增速缓慢,甚至停止[25]。

|
| 图 3 碳密度随林分密度的变化 Fig. 3 Variation of carbon density with tree density in forest |
植被碳密度随林分郁闭度的变化趋势与林分密度类似,依据碳密度随郁闭度变化的外包线(图 4)来看,郁闭度的阈值为0.5,当郁闭度小于0.5时,碳密度随郁闭度增加而线性增大,但当郁闭度超过0.5后植被碳密度不再增加,而是趋近于其极值。

|
| 图 4 碳密度随郁闭度的变化 Fig. 4 Variation of carbon density with forest canopy density |
通常认为,由于竞争光和生长空间,成熟林会表现出明显的密度效应,即当林分密度低于某一阈值时,林分生物量随密度增加而增加,林分密度高于该阈值时,林分生物量不再增加,甚至表现出随林分密度增加而下降的趋势[26]。而在本研究中的林分尚属中幼龄林,即表现出密度效应,这很可能不是由于林木之间竞争光和生长空间的结果,更多的可能是因为竞争水分竞争的结果,因为水分是该区最重要的生态环境限制因子[27],这可以从水分限制区,植被结构与密度的关系得到印证,如在黄土高原陕西省永寿县,由于受水分不足的影响,刺槐林的叶面积指数在林龄为11—15 a时即达到最大值,且林分密度大于900—1000株/hm2时,叶面积指数不再随林分密度的增加而增加[28]。
3.3 森林植被碳密度随生境条件的变化 3.3.1 碳密度与年降水量的关系森林植被碳密度随年降水量减少而显著降低(表 6)。年降水量高于700 mm的Ⅰ区是森林植被碳密度的高值中心,平均为(32.5 ±22.1)t/hm2,在该亚区,植被碳密度低于15 t/hm2的林分占该区森林总面积的25.9%,植被碳密度介于15—45 t/hm2之间的林分所占比例为51.7%,植被碳密度高于45 t/hm2的林分占该区森林总面积的22.4%,此外,植被碳密度高于60 t/hm2的林分均分布在该区。在年降水量600—700 mm的Ⅱ区,植被碳密度为(23.2±17.4)t/hm2,显著低于Ⅰ区(P<0.05),在该亚区,碳密度低于15 t/hm2的林分占该亚区森林总面积的比例高达42.5%,植被碳密度介于15—45 t/hm2之间的林分占40%,而植被碳密度高于45 t/hm2的林分仅占17.5%。在年降水量低于600 mm的Ⅲ区,森林 植被碳密度最低,平均为(10.9 ±11.5)t/hm2,以植被碳密度低于15 t/hm2的林分为主,占该亚区森林总面积的72.7%,植被碳密度介于15—45 t/hm2的林分占22.7%,高于45 t/hm2林分仅占4.5%。
|
亚区
Sub area | 年降水量
MAP/mm | 样地数量
Plot numbers | 植被碳密度
Carbon density for vegetation/ (t/hm2) |
| MAP = Mean annual precipitation; ()外为平均值±标准差,()内为碳密度变化范围; a、b表示差异显著程度,同一列不同字母表示差异显著(P<0.05) | |||
| Ⅰ区 Sub area Ⅰ | >700 | 22 | 32.5±22.1(7.6—120.6)a |
| Ⅱ区 Sub area Ⅱ | 600—700 | 37 | 23.2±17.4(2.6—59.0)b |
| Ⅲ区 Sub area Ⅲ | <600 | 56 | 10.9±11.5(0.67—42.9)c |
森林植被碳密度随海拔升高先是增大,但在达到某一海拔高度后反而下降。最大森林植被碳密度对应的海拔高度变化在1900—2500 m,并随年降水量降低而增高(图 5)。

|
|
图 5
碳密度随海拔的变化
Fig. 5
Variation of carbon density of forest stands along elevation
MAP(Mean annual precipitation)=年均降水量,y = 植被碳密度,x = 海拔
① (1900—2800 m) y= 0.0457x+141.74 R2 = 0.126; ② (1900—2200 m) y= 0.0522x-81.09 R2 = 0.199; ③ (2200—2800 m) y= 0.0468x+133.96 R2 = 0.178; ④ (1700—2400 m) y= 0.041x-74.152 R2 = 0.355; ⑤ (2400—2800 m) y= 0.0472x+135.71 R2 = 0.165 |
在年降水量大于700 mm的I区,植被碳密度的最大值为120.63 t/hm2,出现的海拔高度范围为1900—2100 m,植被碳密度的垂直递减率为4.57 t hm-2 100m-1,这很可能是由于该区受水分的限制不强,更多的是受温度限制所致。
在年降水量为600—700 mm的Ⅱ区,最大森林植被碳密度降低到59.1 t/hm2,并出现在海拔2100—2300 m处;在此海拔高度以下,植被碳密度随海拔高度升高快速增加,其垂直递增率为5.22 t hm-2 100m-1,这可能是因为海拔升高导致湿度增加;而在此海拔高度范围以上,植被碳密度随海拔升高而快速降低,其垂直递减率为4.68 t hm-2 100m-1,与I区相差不大。
在年降水量500—600mm的Ⅲ区,森林植被碳密度受水分的限制十分强烈,最大森林植被碳密度降低到46.37 t/hm2,而其出现海拔高度为2300—2500 m,在此海拔高度以下,植被碳密度随海拔高度升高的增速为4.11 t hm-2 100m-1,这明显小于Ⅱ区;当海拔超过2300—2500 m后,碳密度随海拔升高而降低,垂直递减率为4.72 t hm-2 100m-1,与Ⅱ区基本一致。
3.3.3 碳密度随坡向的变化整体来看,阴坡和半阴坡的森林植被碳密度相对较高,略高于半阳坡,明显高于阳坡,但是,植被碳密度的坡向差异大小受到年降水量影响,表现为不同亚区之间有明显差别(图 6)。

|
| 图 6 不同降水量条件下碳密度随坡向的变化 Fig. 6 The variation of carbon density with different aspects in Liupan Mountains 箱图表示不同坡向碳密度最大值、3/4分位数、平均值、1/4分位数和最小值 |
在年降水量高于700 mm的I区,阴坡、半阴坡、半阳坡的森林植被碳密度接近,依次为(32.3±21.9)、(37.1 ±35.8)、(34.4±19.54) t/hm2,明显高于阳坡的24.5±9.29 t/hm2。半阴坡的森林植被碳密度均值和极值较其它坡向大,而且该坡向33%的林分碳密度高于45 t/hm2,阴坡和半阳坡分别仅为18.3%、23.8%。阳坡碳密度均小于34.3 t/hm2,而且该坡向55.6%的林分的植被碳密度低于30 t/hm2,这主要是因为阳坡的土壤水分条件较差,如在香水河小流域,5—9月份阳坡平均土壤体积含水量在14%以下,而半阴坡和半阳坡平均土壤体积含水量则在20%以上[12]。
在年降水量600—700 mm的Ⅱ区,森林植被碳密度不同坡向间差异规律与I区相似,阴坡、半阴坡、半阳坡森林植被碳密度接近,依次为(26.2±17.7)、(27.4±21.1)、(24.1±10.4) t/hm2,明显高于阳坡的(10.1±5.4) t/hm2。其中半阴坡30%的林分植被碳密度高于45 t/hm2,而阴坡仅为16.7%,半阳坡和阳坡的森林植被碳密度均小于38.7 t/hm2,其中半阳坡66.7%的林分碳密度低于30 t/hm2,而阳坡可能受到水分不足的限制进一步增强,其碳密度进一步降低,其最大森林植被碳密度仅为20.7 t/hm2。
在降水量低于600 mm的Ⅲ区,阴坡森林植被碳密度最大,为(15.5±13.6) t/hm2,其中40%林分植被碳密度介于15—49.27 t/hm2之间;而半阴坡、半阳坡、半阴坡碳密度依次为(10.2±6.7)、(10.4±7.3)、(7.1±5.7) t/hm2,最大值仅17.9 t/hm2,明显低于阴坡,这很可能是因为阴坡的水分条件最好。
然而,非参数检验结果表明,在各亚区的不同坡向之间的森林植被碳密度差异并不显著(P>0.05),这很可能是因为较大的林龄和林分密度差异,掩盖了坡向之间森林植被碳密度的差异。
4 讨论 4.1 六盘山森林植被碳密度与其它地区的比较六盘山的森林植被碳密度平均为26.7 t/hm2,与李海奎等[19]根据第七次森林资源连续清查资料(2004—2008)估算的宁夏森林植被碳密度(25.7 t/hm2)比较接近,但远低于西北地区的45.28 t/hm2、东北地区的49.05 t/hm2和全国的42.82 t/hm2,这主要因为六盘山地区林龄较小、林分稀疏、先锋群落所占比重大、降水量较低所致。这也从另一个角度说明六盘山森林碳储量还有较大的增长空间,固碳潜力较大。
4.2 水分条件的差异是对六盘山森林植被碳密度空间分异的主要原因温度和降水是影响植被碳密度的重要因子,水热因子组合越有利于植物生长的地区,植物生物量越大,植被碳密度也越大[29, 30]。六盘山地处半湿润气候向半干旱气候过渡地区,水分是森林植被生长的主要限制因子[31],因此,从I区到Ⅲ区,随着降水量降低,碳密度显著降低。此外在降水量不同的3个亚区,森林碳密度随海拔及坡向的变化格局并不一致,这很可能是因为这些因子进一步改变了水分条件的格局。
在降水低于600 mm(Ⅲ区)的较低海拔地带(1900—2300 m),可能由于水分相对不足,植被生长受水分不足的限制,随着海拔升高,水分条件改善,并且促进生长的作用明显大于温度降低不利生长的作用,因此随着海拔升高,碳密度增加;而当海拔上升到一定高度(2500 m)以后,低温成为影响植被生长的主导限制因子,随着海拔升高,温度降低,植被碳密度降低。在降水量高于600—700 mm的Ⅱ区,最大森林植被碳密度出现的海拔高度较Ⅱ区有所下降,这很可能是因为该区降水量较Ⅲ区高,水分的抑制作用减轻所致。而在降水量高于700 mm的I区,水分相对充足,植被生长主要受温度的限制,随着海拔升高,温度降低,植被碳密度降低,这与长白山东坡的研究结果一致(海拔范围700—2000 m,年均降水量720—1400 mm)[32]。
坡向通过改变光照及干旱胁迫程度影响植被生长进而影响植被碳密度的大小,一方面,从阴坡至阳坡,光照的增加,有利于有机物的积累,另一方面,潜在蒸散增加,干旱胁迫程度增加,限制植被的生长,二者作用的强弱对比决定了植被生长状况和植被碳密度的大小[15, 33]。在年降水量低于600 mm的地区,植被生长可能受到水分不足的强烈限制,从阴坡至阳坡,碳密度降低,这很可能是因为干旱胁迫对植被生长的抑制作用大于光照对植被生长的促进作用。在年降水量高于600 mm的地区,阳坡碳密度仍然最低,从阳坡至半阳坡,碳密度增大,这说明阳坡及半阳坡可能仍然受到较强烈的干旱胁迫;而从半阴坡至阴坡,碳密度降低,这很可能是因为光照对植被生长的影响超过了干旱胁迫的影响。
4.3 对六盘山地区林-水-碳综合管理的启示本研究表明,林分郁闭度0.5是森林植被固碳的拐点,即当林分郁闭度大于0.5时,森林植被固碳量不再增加,因此从森林植被固碳功能角度判断,林分郁闭度调节至0.5即可,林分郁闭度继续增加,不仅对植被固碳无益,而且还会徒增耗水,这是因为林分的郁闭度越高,林冠截持和蒸散耗水会越多,林地产水量越少[34]。因此,本文认为该区林分郁闭度不宜超过0.5。但林分郁闭度为0.5时,森林的综合功能是否最大,这有待于结合森林的其它功能进一步确定。前人从生物多样性、保土、木材生产角度得出的最佳郁闭度范围为0.4—0.8[35, 36]。因为本研究区(六盘山自然保护区)为黄土高原的水源地,所以其主导功能应为水源涵养功能,因此,如果为了进一步提高该区的产水功能,其最佳的郁闭度可能还需要在0.5的基础上进一步降低,这需要根据森林固碳耗水的定量关系进一步来确定。
5 结论(1)六盘山森林植被碳密度为0.67—120.63 t/hm2,平均(26.17±21.2) t/hm2,天然次生林碳密度为(30.2±21.4) t/hm2,显著高于人工林的(15.7±16.0) t/hm2(P<0.05)。在天然次生林中,主要森林类型碳密度(t/hm2)平均值依次为山杨林(22.3±12.7)<桦木林(28.4±14.7)<柳树林(35.9±20.8)<辽东栎林(43.4±25.2),由先锋群落至顶级群落,碳密度呈增加的趋势,但不同类型间差异并不显著(P>0.05)。
(2)森林植被碳密度与林龄、林分密度和郁闭度等森林结构密切相关。随着林龄增加,碳密度迅速增大,天然林碳密度年平均增速为1.16 t hm-2 a-1,人工林碳密度年平均增速为2.48 t hm-2 a-1。以林分密度1000株/hm2和郁闭度0.5为拐点,低于此拐点,森林植被碳密度随林分密度和郁闭度增加而线性增大;高于此拐点后,森林植被碳密度不再增加,而是趋于一个极值,对天然林和人工林,该极值分别为75.4、34.6 t/hm2。
(3)六盘山森林碳密度随降水量降低而显著降低,由700 mm以上地区的32.5 t/hm2降低至500—600 mm地区的10.9 t/hm2。在Ⅰ区(年降水量大于700 mm)、Ⅱ 区(年降水量600—700 mm)、Ⅲ区(年降水量小于600 mm),最大森林植被碳密度出现的海拔高度分别为1900—2100m、2100—2300m、2300—2500 m。在Ⅰ区和Ⅱ区(年降水量高于600 mm),碳密度从阴坡至阳坡表现出先增加后降低的趋势,在半阴坡最高,阳坡最低;在Ⅲ区(年降水量低于600 mm),从阴坡至阳坡,碳密度呈递减趋势。
| [1] | Gong L, Jin C L. Problems about ecological environment construction and water resources sustainable utilization in northwest China. Journal of Desert Research, 2004, 24(4): 135-139. |
| [2] | Xu X L, Cao M K, Li K R. Temporal-spatial dynamics of carbon storage of forest vegetation in China. Progress in Geography, 2007, 26(6): 1-16. |
| [3] | Grubb M, Vrolijk C, Brack D. The Kyoto Protocol: A Guide and Assessment. London: Royal Institute of International Affairs London, 1999. |
| [4] | Dai J, Yang W J, Jiang L H, Liu G Q. The objective basis on making the development strategy of forestry in the Northwest China. Journal of Northwest Forestry University, 2003, 18(4): 148-150. |
| [5] | Zhao X. Four measures were implemented to improve carbon sequestration in Shaanxi province. (2010-03-09) [2013-06-15]. http://www.forestry.gov.cn/portal/main/s/72/content-386426.html. |
| [6] | Jackson R B, Jobbágy E G, Avissar R, Roy B S, Barrett D J, Cook C W, Farely A K, Maitre D C L, McCarl B A, Murray C B. Trading water for carbon with biological carbon sequestration. Science, 2005, 310(5756): 1944-1947. |
| [7] | Farley K A, Jobbágy E G, Jackson R B. Effects of afforestation on water yield: a global synthesis with implications for policy. Global Change Biology, 2005, 11(10): 1565-1576. |
| [8] | Wang Y H, Yu P T, Feger K H, Sun G, Bonell M, Xiong W, Zhang S L, Xu L H. Annual runoff and evapotranspiration of forestlands and non-forestlands in selected basins of the Loess Plateau of China. Ecohydrology, 2011, 4(2): 277-287. |
| [9] | Wang Y H, Jin M, Yu P T. The environment problems related with forest/vegetation and water resources in China and future research requirements. Forest Research, 2003, 16(6): 739-747. |
| [10] | Yu P T, Krysanova V, Wang Y H, Xiong W, Mo F, Shi Z J, Liu H L, Vetter T, Huang S C. Quantitative estimate of water yield reduction caused by forestation in a water-limited area in northwest China. Geophysical Research Letters, 2009, 36(2): L2406. |
| [11] | Liupan Mountains Reserve of Scientific Survey Editorial Board. Scientific Survey Reports of Liupan Mountains Reserve. Ningxia: Ningxia People′s Press, 1988. |
| [12] | Zhang S L. The Assessment of Impact of Land Use Change and Climate Variability on Hydrological Process in Basin [D]. Beijing: Chinese Academy of Forestry, 2011. |
| [13] | Wang Z Y, Yu P T, Wang S G, Yu Z J, Hu Y Q, Xia G C, Wang Y H, Xiong W. The characteristics of stand structure and spatial distribution of Quercus liaotungensis forests in Liupan Mts.. Forest Research, 2011, 24(1): 97-102. |
| [14] | Wu J G, Zhang X Q, Xu D Y, Zhu G. The storage of biomass carbon under different land use in Liupan Mountain Forest Zone. Forest Research, 2006, 19(3): 277-283. |
| [15] | Liu Y H. The Characteristics of Growth and Carbon Sequestration and Water Consumption in the Small Watershed of Xiangshuihe, Liupan Mountains [D]. Beijing: Chinese Academy of Forestry, 2011. |
| [16] | Shi Z J, Wang Y H, Xu L H, Xiong W, Yu P T, Hu Z S, Gao J X. Effect of topographic form and vegetation type on the runoff coefficient in the Xiangshuihe Watershed of Liupan Mountains. Science of Soil and Water Conservation, 2009, 7(4): 31-37. |
| [17] | Ningxia Liupanshan Bureau of Forestry. Volume Table of Liupan Mountains Forest Zone. Ningxia Liupanshan Bureau of Forestry, 1988. |
| [18] | Cheng T R, Feng J, Ma Q Y, Wang Y T, Kang F F, Feng Z K, Zhang Y L, Deng X R. Carbon pool and allocation of forest vegetations in Xiaolong Mountains, Gansu Province. Acta Ecologica Sinica, 2008, 28(1): 33-44. |
| [19] | Li H K, Lei Y C, Zeng W S. Forest carbon storage in China estimated using forestry inventory data. Scientia Silvae Sinicae, 2011, 47(7): 7-12. |
| [20] | Ge R L, Si Q B L G, Jin R. Research on the growth characteristics of the introduced Pinus sylvestris var. in Maowusu sandy Land. Journal of Arid Land Resources & Environment, 2004, 18(5): 159-162. |
| [21] | Xu B, Guo Z D, Piao S L, Fang J Y. Biomass carbon stocks in China′s forests between 2000 and 2050: A prediction based on forest biomass-age relationships. Science China Life Sciences, 2010, 53(7): 776-783. |
| [22] | Webb R A. Use of the boundary line in the analysis of biological data. Journal of Horticultural Science, 1972, 47: 309-319. |
| [23] | Black B A, Abrams M D. Use of boundary-line growth patterns as a basis for dendroecological release criteria. Ecological Applications, 2003, 13(6): 1733-1749. |
| [24] | Gao Y P. Assessment of Main Plantation Density Effect in the Semiarid Loss Hilly-Gully Regions [D]. Beijing: Beijing Forestry University, 2011. |
| [25] | Ju W Z, Wang X J, Sun Y J. Age structure effects on stand biomass and carbon storage distribution of Larix olgensis plantation. Acta Ecologica Sinica, 2011, 31(4): 1139-1148. |
| [26] | Wu Z Z. Reasonable density-control is an important way to increase the forest stand productivity. World Forestry Research, 1988, 2(5): 42-47. |
| [27] | Baird A J, Wilby R L. Eco-hydrology: Plants and Water in Terrestrial and Aquatic Environments. London: Routledge, 1990. |
| [28] | Zhou H. A Study on the Characters Based on Leaf Area Index of Robinia Pseudoacacia. Xi′an: North West Agriculture and Forestry University, 2011. |
| [29] | Lü C Q, Sun S C. A review on the distribution patterns of carbon density in terrestrial ecosystems. Acta Phytoecologica Sinica, 2004, 28(5): 692-703. |
| [30] | Yu G R. Global Change, Carbon Cycle and Storage in Terrestrial Ecosystem. Beijing: Meteorology Press, 2003. |
| [31] | Liu G, Lu Y C, Li X H, Lei X D, Ning J K. Effects of climatic factors on tree ring growth in Liupanshan region, Northwest China. Journal of Northeast Forestry University, 2009, 37(4): 1-4. |
| [32] | Zhu B, Wang X P, Fang J Y, Piao S L, Shen H H, Zhao S Q, Peng C H. Altitudinal changes in carbon storage of temperate forests on Mt Changbai, Northeast China. Plant Research, 2010, 123(4): 439-452. |
| [33] | Cheng X R, Yu M K, Wu T G, Zhang C X, Zeng G Q. Effect of site condition on carbon storage of Quercus acutissima plantations. Ecology and Environmental Sciences, 2012, 21(10): 1674-1677. |
| [34] | Huang M B, Kang S Z, Li Y S. A comparison of hydrological behaviors of forest and grassland watersheds in gully region of the Loess Plateau. Journal of Natural Resources, 1999, 14(3): 226-231. |
| [35] | Li J L, Zheng X X, Lu Y C. Optimized stand structure regulation for watershed forests in Liupan Mountains. Forest Resources Management, 2012, (5): 31-38. |
| [36] | Wang Y H. A quantitative study on the benefits of Block Locust (Robinia Pseudoacacia L.) on water and soil conservation in the eastern loess area in Gansu province. Journal of Beijing Forestry University, 1986, 8(1): 35-52. |
| [1] | 贡力, 靳春玲. 西北地区生态环境建设和水资源可持续利用的若干问题. 中国沙漠, 2004, 24(4): 135-139. |
| [2] | 徐新良, 曹明奎, 李克让. 中国森林生态系统植被碳储量时空动态变化研究. 地理科学进展, 2007, 26(6): 1-16. |
| [4] | 代军, 杨文杰, 姜丽华, 刘国强. 制定西北地区林业发展战略的客观依据. 西北林学院学报, 2003, 18(4): 148-150. |
| [5] | 赵侠.陕西四项措施增加森林碳汇. (2010-03-09) [2013-06-15]. http://www.forestry.gov.cn/portal/main/s/72/content-386426.html. |
| [9] | 王彦辉, 金旻, 于澎涛. 我国与森林植被和水资源有关的环境问题及研究趋势. 林业科学研究, 2003, 16(6): 739-747. |
| [11] | 六盘山自然保护区科学考察编辑委员会. 六盘山自然保护区科学考察报告. 宁夏: 宁夏人民出版社, 1988. |
| [12] | 张淑兰. 土地利用和气候变化对流域水文过程影响的定量评价 [D]. 北京: 中国林业科学研究院, 2011. |
| [13] | 王占印, 于澎涛, 王双贵, 余治家, 胡永强, 夏固成, 王彦辉, 熊伟. 宁夏六盘山区辽东栎林的空间分布及林分特征. 林业科学研究, 2011, 24(1): 97-102. |
| [14] | 吴建国, 张小全, 徐德应, 朱高. 六盘山林区几种土地利用方式植被活体生物量C贮量的研究. 林业科学研究, 2006, 19(3): 277-283. |
| [15] | 刘延惠. 六盘山香水河小流域典型植被生长固碳及耗水特征 [D]. 北京: 中国林业科学研究院, 2011. |
| [16] | 时忠杰, 王彦辉, 徐丽宏, 熊伟, 于澎涛, 胡哲森, 高吉喜. 六盘山香水河小流域地形与植被类型对降雨径流系数的影响. 中国水土保持科学, 2009, 7(4): 31-37. |
| [17] | 六盘山林管局. 六盘山林区材积表. 六盘山林管局, 1988. (未链接到本条文献信息) |
| [18] | 程堂仁, 冯菁, 马钦彦, 王玉涛, 康峰峰, 冯仲科, 张彦林, 邓向瑞. 甘肃小陇山森林植被碳库及其分配特征. 生态学报, 2008, 28(1): 33-44. |
| [19] | 李海奎, 雷渊才, 曾伟生. 基于森林清查资料的中国森林植被碳储量. 林业科学, 2011, 47(7): 7-12. |
| [20] | 格日勒, 斯琴毕力格, 金荣. 毛乌素沙地引种樟子松生长特性的研究. 干旱区资源与环境, 2004, 18(5): 159-162. |
| [21] | 徐冰, 郭兆迪, 朴世龙, 方精云. 2000—2050 年中国森林生物量碳库: 基于生物量密度与林龄关系的预测. 中国科学: 生命科学, 2010, 40(7): 587-594. |
| [24] | 高艳鹏. 半干旱黄土丘陵沟壑区主要树种人工林密度效应评价 [D]. 北京: 北京林业大学, 2011. |
| [25] | 巨文珍, 王新杰, 孙玉军. 长白落叶松林龄序列上的生物量及碳储量分配规律. 生态学报, 2011, 31(4): 1139-1148. |
| [26] | 吴增志. 合理的密度管理是提高林分生产力的重要途径——从世界种群密度理论研究的现状看深入研究的必要性. 世界林业研究, 1988, 2(5): 42-47. |
| [28] | 周慧. 刺槐人工林叶面积指数特征研究. 西安: 西北农林科技大学, 2011. |
| [29] | 吕超群, 孙书存. 陆地生态系统碳密度格局研究概述. 植物生态学报, 2004, 28(5): 692-703. |
| [30] | 于贵瑞. 全球气候变化于陆地生态系统碳循环和碳蓄积. 北京: 气象出版社, 2003. |
| [31] | 刘刚, 陆元昌, 李晓慧, 雷相东, 宁金魁. 六盘山地区气候因子对树木年轮生长的影响. 东北林业大学学报, 2009, 37(4): 1-4. |
| [33] | 成向荣, 虞木奎, 吴统贵, 张春祥, 曾广泉. 立地条件对麻栎人工林碳储量的影响. 生态环境学报, 2012, 21(10): 1674-1677. |
| [34] | 黄明斌, 康绍忠, 李玉山. 黄土高原沟壑区森林和草地小流域水文行为的比较研究. 自然资源学报, 1999, 14(3): 226-231. |
| [35] | 李金良, 郑小贤, 陆元昌. 六盘山水源林林分结构优化调整研究. 林业资源管理, 2012, (5): 31-38. |
| [36] | 王彦辉. 陇东黄土地区刺槐林水土保持效益的定量研究. 北京林业大学学报, 1986, 8(1): 35-52. |