 2014, Vol. 34
2014, Vol. 34文章信息
- 杨凤萍, 胡兆永, 侯琳, 蔡靖, 崔翠, 张硕新
- YANG Fengping, HU Zhaoyong, HOU Lin, CAI Jing, CUI Cui, ZHANG Shuoxin
- 秦岭火地塘林区油松和华山松林乔木层净生产力与气候因子的关系
- Relationship between net primary productivity of tree layer in Pinus tabulaeformis and Pinus armandi forests and climatic factors at Huoditang forest region in the Qinling Mountains
- 生态学报, 2014, 34(22): 6489-6500
- Acta Ecologica Sinica, 2014, 34(22): 6489-6500
- http://dx.doi.org/10.5846/stxb201302140264
-
文章历史
- 收稿日期:2013-2-14
- 网络出版日期:2014-3-17






2. 陕西秦岭森林生态系统国家野外科学观测研究站, 宁陕 711600
2. Qinling National Forest Ecosystem Research Station, Huoditang, Ningshan 711600, China
森林生态系统生物量和生产力是研究生态系统碳平衡的基础数据[1],同时也是衡量森林植被群落在自然环境条件下生产能力的重要指标,它的变化直接反映了生态系统对环境气候条件的响应。对森林生产力的研究一直都是全球气候变化研究的焦点之一[2]。国内外专家运用不同的研究方法对森林生产力动态变化及其对气候变化响应问题开展了广泛的研究。样地调查法是应用最普遍的方法[3, 4]。Fang等结合50a的森林资源清查资料和森林生物量实测资料,得到中国森林碳库自20世纪70年代末期开始呈增加趋势,并推测气温上升和CO2浓度施肥可能是促进森林生长的重要因子[5]。但是由于此方法耗时长,难以获取年际动态信息,近年来模型模拟法被广泛应用于森林生产力动态模拟[6, 7]。赵敏等[8]利用CENTURY模型模拟了中国北方林近35a来的生产力动态,并认为气温是影响生产力波动的主要气候因子。赵俊芳等[9]利用碳收支模型FORCCHN模拟气候变化对东北森林生态系统净第一性生产力(NPP)和碳收支(NEP)的影响,发现该生态系统NPP和NEP对温度升高比对降雨变化更为敏感。但由于模型本身的局限性及相关背景参数的不完善,使得模型输出结果的不确定性增加。Nemani等[10]、Zhao等[11]利用遥感观测资料从全球尺度上估算了植被净初级生产力及其对气候变化的响应,得出在1982—1999年间云量减少和太阳辐射增加是导致其全球植被净初级生产力增加的主要因素;而2000—2009年间全球植被净初级生产力的下降是干旱造成的,然而遥感观测资料较低的分辨率限制了其研究结果的精度。为了弥补以上方法的不足,国内外已有运用树木年轮法估算长期生物量和净生产力动态的尝试[12, 13]。树木年轮资料在一定程度上可反映树木在自然条件下的长期生长特征,具有分辨率高、样本分布广泛、时间序列长、定年准确、环境变化指示意义明确且可定量等优点,在估算森林生物量和生产力长期动态方面具有一定优势。滕菱等[14]利用树木年轮资料揭示了长期气温波动与鼎湖山马尾松种群生产力的正相关关系。程瑞梅等[15]的研究表明北亚热带马尾松年际变化主要是气候因素引起的,在未来该地区升温增湿的条件下马尾松林的生产力可能会升高。彭俊杰等[16]应用BIOME-BGC模型和树木树轮数据模拟1952—2008年华北地区油松林生态系统净初级生产力动态,发现5—6月平均温度和上年8月至当年7月的降水是限制油松林生态系统NPP的主要因子,随着区域暖干化趋势的加强,生态系统NPP呈下降趋势。
秦岭作为长江和黄河两大水系的分水岭,是我国南北气候交流的屏障,也是我国暖温带和亚热带气候的分界线[17],属于我国气候变化敏感区域,有关森林生产力动态变化与气候因子相互作用关系的研究还比较匮乏。蒋冲等[18]采用气候相关模型及统计方法分析了秦岭南北近52 a气象要素的时空变化特征及其对植被净初级生产力的影响,认为降水的增多会促进植被净初级生产力的累积,但生产力限制因子呈现出地区差异。油松和华山松林是秦岭的主要森林类型之一,其蓄积量占秦岭林区针叶树种总蓄积量的80%[19]。因此,本文选取秦岭火地塘林区油松林、华山松林为研究对象,以树木年轮学为基础,分析其在长时间序列上生物量、生产力的动态变化特征,探索其乔木层生产力与气候因子的关系,以期为森林经营措施的制定提供科学依据。
1 研究区概况研究区位于秦岭南坡中段火地塘林区陕西秦岭森林生态系统国家野外科学观测研究站(E 108°21′—108°29′,N 33°18′— 33°28′),年平均气温为8—10 ℃,年降水量900—1200 mm,年蒸发量800—950 mm,年日照时数1100—1300 h,无霜期170 d,海拔在1450—2470 m之间。区内地形复杂,为花岗岩和片麻岩石质山地,平均坡度在35°左右。土壤为火成岩、变质岩以及石灰岩母质上发育的中到弱酸性山地棕壤。火地塘林区在20世纪60—70年代经历了一场全面的森林主伐,原生植被几乎荡然无存。到20世纪90年代,森林基本恢复,形成大面积的天然次生林,平均林龄40a以上。主要成林树种有油松(Pinus tabulaeformis)、华山松(Pinus armandi)、锐齿栎(Quercus aliena var. acuteserrata)、红桦(Betula albo-sinensis)等。林区森林覆被率高达93.8%。
2 研究方法 2.1 样地选择与采样在充分考虑海拔、起源、林龄等因素的基础上,选择垂直分布具有代表性地段的油松林和华山松林,共设置20 m×20 m标准地24块(表 1)。对样地内所有乔木进行每木检尺。根据样地调查数据,将林木按径级分组(4 cm为1个径级),一般在株数多的中央径级选2—3株,其他径级选1—2株,对两端径级、特别是最大径级至少要有一株标准木作为钻芯树木。在距地面1.3 m处沿平行山坡等高线方向钻取树芯,钻芯至树干髓心处即可,每株树钻取2根树芯[20],现场测量湿树芯的长度,并及时装入自制的纸管内,带回实验室分析。
|
森林类型
Forest type | 样地数
Number of plots | 海拔
Elevation/m | 坡度
Slope/(°) | 坡向
Aspect/(°) | 林分密度
Stand density/ (株/hm2) | 郁闭度
Canopy density | 平均胸径
Mean diameter at breast height (DBH) /cm | 林龄
Stand age | 样芯数
Number of cores |
|
油松
P.tabulaeformis | 9 | 1400—1726 | 16—38 | 211—261 | 816—1575 | 0.6—0.8 | 22.3—26.1 | 41—47 | 196 |
|
华山松
P.armandi | 15 | 1410—1983 | 24—34 | 113—288 | 800—1433 | 0.7—0.8 | 21.6—24.4 | 40—45 | 314 |
将所采树芯带回实验室中,常温下干燥一周,测量干样芯的长度,以便计算树芯干湿长度比。油松和华山松的干湿长度比分别为0.979、0.991,可见,干、湿树芯长度差距不大,由此引起的生物量计算误差很小,故可忽略不计。将干燥后的树芯固定到木槽中,用不同粗细的砂纸依次打磨至表面平整光滑,年轮界限清晰。用骨架图法[21]交叉定年,再用WinDENDROTM年轮分析系统扫描样芯。并基于扫描图像测量年轮宽度(该系统测量精度为0.01 mm)。
2.3 生物量估算
借鉴程瑞梅等[15]的方法,单木历年胸径Dn-1通过当年胸径Dn减去当年两个树芯宽度之和Wn(式(1))。用陈存根等[22]建立的树高与胸径的关系推算出单木历年树高Hn(表 2)。根据已有的生物量回归模型[22](表 2)计算单木各器官历年的生物量,它们之和即是单木历年的生物量Bn。依据各样地的林木密度和径级分组情况,首先计算出各径级组树木历年的生物量,然后相加得到各样地乔木层历年的生物量Pn,样地乔木层历年生物量之差就是样地乔木层的生产力NPP(式(2))。最后分别利用各样地乔木层历年生物量之和除以样地数得出油松林和华山松林乔木层历年平均生物量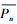 (式(3))和生产力
(式(3))和生产力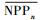 (式(4)),并换算成单位面积(每公顷)生物量和生产力。具体计算公式如下:
(式(4)),并换算成单位面积(每公顷)生物量和生产力。具体计算公式如下:

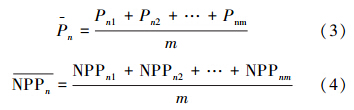 表示平均生物量,表示平均生产力,m表示样地个数。
表示平均生物量,表示平均生产力,m表示样地个数。
 |
气象数据来自于离火地塘林区最近的陕西省宁陕县气象站(E108°18′54″,N33°18′39″)1973—2011年的年降水量、年均温、逐月降水量和逐月平均温度。由于温度、降水不仅影响当年树木的生长,还会影响到下一年甚至以后几年树木的生长[23],因此本研究采用上年5月到当年12月的气象资料进行分析。
气候要素趋势变化率用一次线性方程表示[24],公式为:

根据样地调查资料(不同径级林木株数),计算各样地不同径级林木密度(单位面积林木株数),油松林和华山松林不同径级林木密度为所有样地的平均值。通过查定各样地不同径阶钻芯树木的年龄,计算各径阶算术平均年龄,油松林和华山松林不同径级平均年龄为所有样地的平均值。
2.5.2 平均敏感度在树木年轮的气候研究中,把年轮宽度逐年变化状况作为树木对气候反映的敏感度[23]。
计算公式为:
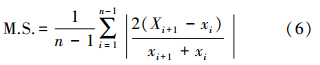
对油松林和华山松林乔木层生产力与上年5月至当年12月单月以及不同月份组合的平均温度、降水序列进行Pearson相关分析,在此基础上,将相关显著的月值气象因子进一步进行多元逐步回归分析,筛选出对树木生长影响最大的气象因子。所有的统计分析用IBM SPSS Statistics 20.0软件处理。
3 结果与分析 3.1 油松、华山松林径级分布与年龄结构单位面积油松和华山松林各径级林木株数(密度)随着径级的增大而减小(图 1,图 2)。小径级林木个体占林分主体部分。其中油松林和华山松林8—12 cm,12—16 cm,16—20 cm,20—24 cm 4个径级林木株数分别占整个林分的62%和78%,36—40 cm和>40 cm径级的林木株占整个林分的6.2%和1.9%。油松和华山松林林木平均年龄随径级增加 而增大(图 1,图 2)。油松林木平均年龄变化幅度在35—50a之间,华山松变化幅度在30—45a之间。

|
| 图 1 油松林径级分布与年龄结构 Fig. 1 Diameter class distribution and age structure of P. tabulaeformis forest |

|
| 图 2 华山松林径级分布与年龄结构 Fig. 2 Diameter class distribution and age structure of P. armandi forest |
从年轮宽度统计特征可以看出(表 3),油松和华山松逐年年轮宽度变化较大,华山松对气候变化的敏感度略高于油松。
|
树种
Tree species | 时段
Time period | 最大轮宽
Maximum tree ring width/mm | 最小轮宽
Minimum tree ring width/mm | 平均轮宽
Average tree ring width/mm | 平均敏感度
Mean sensitivity |
| 油松P. tabulaeformis | 1973—2011 | 9.46 | 0.13 | 2.18±1.29 | 0.175—0.238 |
| 华山松P. armandi | 1973—2011 | 10.52 | 0.10 | 2.29±1.56 | 0.193—0.261 |
在全球变暖的背景下,研究区近39年平均气温整体呈上升趋势(图 3),变化曲线线性拟合倾向率为每10年0.234 ℃。多项式拟合呈一峰一谷型,1973—1984年间年均气温呈下降趋势,最低值出现在1984年,为11.63 ℃。而后波动上升,最大值出现在1998年,为13.18 ℃,之后呈下降趋势。
近39年研究区年降水量变化趋势不明显(图 3),年降水量呈现随机变化过程,其最大值出现在2003年,为1298.3 mm,最小值出现在2001年,为575.4 m。变幅为722.9 mm,年际变化较大。

|
| 图 3 1973到2011年的年平均温度和降水量 Fig. 3 Annul mean temperature and precipitation from 1973 to 2011 |
研究区气候变化属典型的雨热同季的气候特征(图 4)。月平均气温与降水量最高值均出现在7月份。

|
| 图 4 39a月平均气温和降水量年内分布 Fig. 4 Monthly mean temperature and precipitation |
油松和华山松林乔木层单位面积生物量在过去39年中增长迅速(图 5),分别从1973年的15.32 t/hm2和7.53 t/hm2增长到2011年的175.98 t/hm2、130.98 t/hm2,油松林乔木层单位面积生物量一直高于华山松林,且二者差异显著(n=39,P<0.01),且随着时间推移差异逐渐增大。
从1973年到2011年的39年间,油松和华山松林乔木层生产力均呈现波动上升趋势(图 5),两者的变化曲线线性拟合倾向率为每10年0.288 t/hm2和 0.661 t/hm2,华山松林乔木层NPP变化曲线拟合倾向率大于油松林,说明39a间华山松林生产力增长较快。两者在1973—1998年间稳定渐增,1998年,两者有较大起伏波动,并在2001年达到最大值,随后呈下降趋势。与华山松林相比,油松林乔木层生产力波动较大。两者最小值均出现在1973年分别为2.50 t hm-2 a-1和1.28 t hm-2 a-1,最大值均出现在2001年,分别为5.62 t hm-2 a-1和4.21 t hm-2 a-1,多年平均值分别为4.18 t hm-2 a-1和3.19 t hm-2 a-1,对其生产力进行Pearson相关分析结果表明,两者相关极显著(P<0.001),相关系数达到0.795,两者波动一致性较好,说明除了自身生物学特性造成的生产力差异外,两者生长可能受到相同外界条件因素的限制。与华山松林相比,油松林生产力一直高于华山松林,但两者的差距在2001年后逐渐缩小。

|
| 图 5 1973年到2011年油松林、华山松林生物量和生产力变化 Fig. 5 Biomass and net primary productivity (NPP) dynamics from 1973 to 2011 |
油松林生产力与年降水量负相关系数为-0.243(P>0.05)(图 6);各月降水量与其生产力的负相关系数最高仅为-0.28(9月降水及上年9月降水),而上年6月、12月和当年12月降水量与年净生产力呈些微正相关。华山松林生产力与年降水量相关系数仅为-0.084(图 6);单月气象因子与华山松林生产力相关分析结果显示,上年9月、当年9月、10月的降水与华山松林生产力负相关系数分别为-0.279、-0.281、-0.174。而上年8月、当年6月、11月的降水与生产力相关系数分别为0.211、0.215、0.181和0.159。总体来说,降水与华山松林生产力的相关关系均未达到显著水平(P>0.05)。
|
|
| 图 6 降水与油松林和华山松林乔木层生产力的相关性 Fig. 6 Correlations between precipitation and NPP P表示上年各月;Yp表示年降水量;*表示显著水平小于0.05,**表示显著水平小于0.01 |
年平均温度对油松林生产力呈正相关(图 7),相关系数达0.516,极显著(P<0.01)。各月平均温度对油松林生产力的相关关系大多表现为正相关(图 7),其中上年9月、当年1月、3月平均温度与生产力的相关关系显著(P<0.05),相关系数分别为0.318、0.350和0.368,而上年7月、当年6月、7月平均温度与生产力的相关关系达到了极显著水平(P<0.01),相关系数都较高,其中当年7月平均温度对油松生长影响最大,相关系数最高为0.498,其次是上年7月、当年6月的平均温度,相关系数分别为0.482、0.472、0.450和0.426。

|
| 图 7 温度与油松和华山松林生产力的相关性 Fig. 7 Correlations between temperature and NPP Yt表示年平均温度;*表示显著水平小于0.05,**表示显著水平小于0.01 |
与油松相似,年平均温度与华山松生产力呈正相关(图 7),相关系数为0.403,相关达到显著水平(P<0.05)。单月平均温度与华山松生产力主要表现为正相关。其中上年7月的平均温度与净生产力的相关系数最高达0.364,且相关关系显著(P<0.05),而当年6月、7月的平均温度与生产力的相关系数分别为0.358、0.360、0.319和0.342,且均达到显著水平(P<0.05)。
由于各季节气候状况对树木生长的影响有更好的代表性,因此,进一步对单月气候因子进行月份组合之后并与油松林生产力做相关分析后得出,对油松林生产力影响最显著的气象因子是1—7月组合后的平均温度(图 7),相关系数最高为0.563,且达到极显著水平(P<0.001);对华山松林生产力影响最显著的气象因子是5—7月平均温度(图 7),相关系数最高为0.430,达到极显著水平(P<0.01)。
3.6 气象因子与生产力的回归分析 3.6.1 单月平均温度与油松林生产力上年9月、当年1月、3月平均气温与油松林生产力的相关显著(P<0.05),而上年7月、当年6月、7月平均气温与生产力的相关达到极显著水平(P<0.01),进一步建立的最优多元线性回归方程为:
NPP油=-8.959+0.291T7+0.276Tp7
式中,NPP油表示油松林乔木层净生产力,T7表示当年7月平均温度序列,TP7表示上一年7月平均温度序列。方程对1973—2011年油松林净生产力变化的方差解释量为38.1%,调整自由度后方差解释量为34.7%,各回归系数达到的显著水平(P<0.01)。回归方程的自变量为上年7月和当年7月平均温度序列,说明在研究时段内(1973—2011年)上年7月和当年7月平均温度对油松林年净生产力的影响最大。
3.6.2 单月平均温度与华山松生产力上年7月、当年6月、7月平均气温与华山松净生产力的相关显著(P<0.05),进一步建立一元线性回归方程为:
NPP华=-4.204+0.319Tp7
式中,NPP华为华山松林乔木层净生产力,TP7为上一年7月平均温度序列。该方程的相关系数r为0.364(P<0.05),方程对1973—2011年华山松林净生产力变化的方差解释量为13.2%,调整自由度后方差解释量为10.9%。华山松林年净生产力与单月平均温度回归分析的相关系数r及方差解释量相对油松较低。
4 讨论 4.1 生物量与生产力秦岭火地塘林区油松和华山松林乔木层平均生物量分别从1973年的15.32、7.53 t/hm2上升到2011年的175.98、130.98 t/hm2,在39a间增长迅速,且油松林乔木层生物量一直显著高于华山松林。1996年的油松和华山松林乔木层平均生物量分别为106.49 t/hm2和74.71 t/hm2,与陈存根等[22]1996年实测的油松和华山松林的平均生物量107.90 t/hm2和84.72 t/hm2相近,两者的研究对象均是20世纪60、70年代森林皆伐后经过封山育林恢复起来的天然次生林,林龄和林分密度差距不大,但两者也有微小差距,可能是样地位置不同所致。中小尺度森林生态系统样地设置面积较小,且设置时倾向于选择树木生长良好、包含大径级林木的地段,因此通常会造成估算的生物量和生产力偏高,并且不同研究人员对样地选择、采样标准有差异,最终导致结果不完全一样。
本研究得出1985年油松林生物量为60.44 t/hm2,与马钦彦[25]估算的陕南地区10—20年生油松林生物量(47.64 t hm-2 a-1)相比偏大,这是由于后者是整个陕南地区的油松林生物量平均值。吴刚等[26]认为四川巫溪30年生油松林生物量和生产力为145.593 t/hm2和11.34 t hm-2 a-1,而本研究1994年油松林生物量和生产力分别为97.87 t/hm2和4.36 t hm-2 a-1,相对偏小。这可能是因为本研究林龄较小,且研究区地处亚热带最北缘,热量和水分条件与四川巫溪地区有差异。1995年华山松林乔木层生物量和生产力与吴刚等[27]推算的秦岭中山地带30—31年生华山松林乔木层生物量和生产力分别为71.72 t/hm2,3.68 t hm-2 a-1非常接近。
从1973年到2011年油松林和华山松林生产力整体上呈波动上升趋势。与侯英雨等[28]、方精云等[29]估算的全国范围内的植被生产力年际变化趋势基本一致。两者生产力在1973年最低分别为2.51 t hm-2 a-1和1.28 t hm-2 a-1,2001年最高分别为4.21 t hm-2 a-1和5.62 t hm-2 a-1,39a平均生产力为4.18 t hm-2 a-1和3.19 t hm-2 a-1。根据李登科等[30]、李晶等[31]在陕南、秦巴山区的研究结果,1981—2006年间该地区植被净初级生产力的年际变化趋势为增加为主,与本研究结果一致。蒋冲等[18]详细分析了1961—2011年秦岭南北植被净初级生产力动态变化趋势,得出秦岭以南地区增加趋势更为明显,特别是与本研究区相近的佛坪、镇安地区,生产力曲线线性拟合倾向率分别为每10年0.668 t/hm2和1.265 t/hm2,与本研究结果一致。
3 9a间油松林生产力一直高于华山松,这可能与两个树种的生物学特性有关。秦岭是油松和华山松分布的交汇区,油松主要分布在秦岭及其以北地区,而华山松分布在秦岭及其以南地区,油松抗逆性优于华山松,所以在降水不是限制因子的前提下,油松林具有更高的生产力[32]。根据华山松林和油松林的年龄结构和径级分布情况,华山松林乔木层年龄较大的个体相对较少,年龄较小的个体占多数。而油松林乔木层中年龄中等和较大的个体所占比例较华山松林多(图 1,图 2),油松种子生产能力强于华山松,且前者具有较大的高生长和侧生长速率,使得1973—1998年间油松存活和定居能力强于华山松,在此期间华山松林乔木层生产力较低,且增速不如油松林[32]。后期随着油松林个体数量增加,油松林占据了较大水平空间,幼树不能得到充足的光照和水分,种内竞争加剧,使得油松定居数量下降,幼树生长受到抑制,而华山松定居数量和生长相对稳定,因此在2001年后两者生产力差距逐渐减小。
此外,油松林和华山松林乔木层生产力动态变化有较大一致性(R=0.795,P<0.01)。说明除了两者的生物学特性导致的生产力差异外,两者共同的外界环境因素可能是引起生产力变化的主要因素。
4.2 气候变化对生产力的影响降水量对油松和华山松林生产力的影响不显著。这主要是因为火地塘林区降水充足,通常情况下水分条件能够满足树木生长的需要,因此降水不是限制该地区树木生长的气候因子。因此本文重点阐述温度对油松林和华山松林生产力的影响。
温度是影响秦岭火地塘林区油松、华山松生产力的主要气候因子,表现出正相关性。此结果很好地印证了方精云等[33]关于气温上升可能是促进森林生长的重要因子的推测。蒋冲等[18]也指出气温上升与降水增加共同导致秦岭以南的大部分地区植被净初级生产力的增加。这是由于秦岭火地塘林区雨量充沛,温度的上升有利于油松和华山松的生长。油松和华山松的生长与温度的年际变化有较高的一致性。在全球气候变暖的背景下油松林和华山松林生产力整体上呈波动上升趋势,且两者与温度年际变化的峰谷对应较好。
油松林和华山松林生产力与绝大多数月平均气温的相关系数为正值,在剔除了与油松林和华山松林生产力相关不紧密单月气候因子后,上年7月均温、当年7月均温对油松林生产力影响最大,上年7月均温对华山松林生产力影响最大。这可能与火地塘林区气温和降水量最高值均出现在7月有关,此时林内湿度较高,温度较低,不利于光合作用,使得7月的平均温度成为限制树木生长的主要因子。而上年7月均温对当年生产力的显著影响说明温度对植物生长的影响具有“滞后效应”,上年温度状况可以通过影响光合产物的积累及落叶前养分的回运等而影响下一年甚至几年的生长[34]。
火地塘油松林生产力与1—7月平均温度正相关关系达到极显著水平。从单月气温与油松生产力的相关可看出1—7月份中各月平均气温存在较好的正相关关系,1、3月份达到了0.05的显著水平,6、7月份达到0.01显著水平。这是因为1—7月是油松生长的关键时期。秦岭地区油松快速生长期一般在6月下旬或7月初[35],而生长前期1—4月的温度的高低将直接影响树木生长期开始的早晚,前期温度较高使得树木形成层的细胞分裂早,有利于生长期的提前,从而在客观上使生长期提前,形成较宽的年轮[36]。进入生长期后,气温达到了一年内最高时期,土壤及树木本身蒸腾也随气温升高间接达到一年内最高期。同时,5—8月是油松耗水高峰期,最高旬耗水量为7月上旬,虽然对水分条件要求很高,但是由于研究区雨热同季的特点,降水亦达到一年内的最高时期,降水增加能够满足油松生长和土壤及树木本身蒸腾对水分需求,该过程中温度上升对土壤水分影响不大,但能够加快树木新陈代谢的速度,从而提高生长速率,增大生产力。
华山松林生产力与5—7月平均温度正相关关系达到极显著水平。由于形成层在春季平均温度或积温到达一定值后才开始活动,即形成层活动存在温度阈值[37]。火地塘林区1—4月温度较低不足以对研究区华山松形成层活动产生显著影响,而随着5—7月温度上升并达到一年中最高值,刺激华山松形成层活动,从而影响当年华山松生长。曹受金等[38]的研究结果与本研究相似,并认为夏季降水量较为充沛的地区,生长季平均气温是树木生长的主要气候限制因子。当年生长季较高的温度有利于光合作用,从而有利于树木有机质的积累,而温度较低导致树木生长期缩短则不利于树木的生长[39, 40]。
5 结论秦岭火地塘林区油松和华山松林乔木层生物量从1973年到2011年随着年龄的增长呈上升趋势,分别从15.32 t/hm2、7.53 t/hm2增长到175.98 t/hm2、130.98 t/hm2;39a间油松林和华山松林生产力均呈波动上升趋势,平均年净生产力分别为4.18 t hm-2 a-1和3.20 t hm-2 a-1,油松林乔木层生物量和生产力均高于华山松林,两种林型的生物量和生产力差异是由于油松和华山松生物学特性所致,而生物量和生产力长期动态变化受两种林型演替规律和气候因子的影响。油松和华山松林乔木层生产力与温度年际变化有较高的一致性,随着温度的升高而增大,与降水的关系不紧密;单月气候因子中,上年7月温度和当年7月温度对油松乔木层生产力对响应强烈;油松林生产力与1—7月平均温度关系密切,而华山松林生产力与5—7月平均温度关系密切,油松林生产力与温度因子的相关性高于华山松林。
致谢: 野外采样过程中得到王来、仲崇高同学的帮助,特此致谢。
| [1] | Xiang W H, Tian D L, Yan W D. Review of researches on forest biomass and productivity. Central South Forest Inventory and Planning, 2003, 22(3): 57-60, 64-64. |
| [2] | Scurlock J M O, Cramer W, Olson R J, Parton W J, Prince S D. Terrestrial NPP: toward a consistent data set for global model evaluation. Ecological Applications, 1999, 9(3): 913-919. |
| [3] | Metsaranta J M, Lieffers V J. Using dendrochronology to obtain annual data for modelling stand development: a supplement to permanent sample plots. Forestry, 2009, 82(2): 163-173. |
| [4] | Wang B, Liu M C, Zhang B. Dynamics of net production of Chinese forest vegetation based on forest inventory data. Forest Resources Management, 2009, (1): 35-43. |
| [5] | Fang J Y, Chen A P, Peng C H, Zhao S Q, Ci L J. Changes in forest biomass carbon storage in China between 1949 and 1998. Science, 2001, 292(5525): 2320-2322. |
| [6] | Cao M, Woodward F I. Dynamic responses of terrestrial ecosystem carbon cycling to global climate change. Nature, 1998, 393(6682): 249-252. |
| [7] | Tian H Q, Chen G S, Liu M L, Zhang C, Sun G, Lu C Q, Xu X F, Pan S F, Chappelka A. Model estimates of net primary productivity, evapotranspiration, and water use efficiency in the terrestrial ecosystems of the southern United States during 1895-2007. Forest Ecology and Management, 2010, 259(7): 1311-1327. |
| [8] | Zhao M, Zhou G S. Modeling variation trend of boreal forest NPP in China and its relations to temperature and precipitation. Acta Botanica Boreali-Occidentalia Sinica, 2005, 25(3): 466-471. |
| [9] | Zhao J F, Yan X D, Jia G S. Simulating the responses of forest net primary productivity and carbon budget to climate change in Northeast China. Acta Ecologica Sinica, 2008, 28(1): 92-102. |
| [10] | Nemani R R, Keeling C D, Hashimoto H, Jolly W M, Peter S C, Tucker C J, Myneni R B, Running S W. Climate-driven increases in global terrestrial net primary production from 1982 to 1999. Science, 2003, 300(5625): 1560-1563. |
| [11] | Zhao M S, Running S W. Drought-induced reduction in global terrestrial net primary production from 2000 through 2009. Science, 2010, 329(5994): 940-943. |
| [12] | Graumlich L J, Brubaker L B, Grier C C. Long-term trends in forest net primary productivity: Cascade Mountains, Washington. Ecology, 1989, 70(2): 405-410. |
| [13] | Krakauer N Y, Randerson J T. Do volcanic eruptions enhance or diminish net primary production? Evidence from tree rings. Global Biogeochemical Cycles, 2003, 17(4): 1118. |
| [14] | Teng L, Peng S L, Hou A M, Xie Z Y. Effect of air temperature change on the productivity of Pinus Massoniana population in Dinghushan. Journal of Tropical and Subtropical Botany, 2001, 9(4): 284-288. |
| [15] | Chen R M, Feng X H, Xiao W F, Wang R L, Wang X R, Du H T. Response of net productivity of Masson Pine plantation to climate change in north subtropical region. Acta Ecologica Sinica, 2011, 31(8): 2086-2095. |
| [16] | Peng J J, He X Y, Chen Z J, Cui M X, Zhang X L, Zhou C H. Response of Pinus tabulaeformis forest ecosystem in north China to climate change and elevated CO2: A simulation based on BIOME-BGC model and tree-ring data. Chinese Journal of Applied Ecology, 2012, 23(7): 1733-1742. |
| [17] | Liu H, Lei R D, Hou L, Wang F Q. Research on carbon storage and carbon density of main forest types at Huoditang forestry region in the Qinling Mountains. Journal of Northwest A&F University: Natural Science Edition, 2009, 37(3): 133-140. |
| [18] | Jiang C, Wang F, Mu X M, Li R. Effects of climate change on net primary productivity of vegetation in the northern and southern regions of the Qinling Mountains (Ⅱ): Net primary productivity of vegetation in recent 52 a. Science of Soil and Water Conservation, 2012, 10(6): 45-51. |
| [19] | Liu H, Hou L, Lei R D. Carbon storage and carbon density of Pinus tabulaeformis and Pinus armandii forests at Huodingtang forest region in Qinling Mountain. Chinese Journal of Eco-Agriculture, 2007, 15(1): 5-7. |
| [20] | Zhang Y D, Liu Y C, Liu S R, Zhang X H. Dynamics of stand biomass and volume of the tree layer in forests with different restoration approaches based on tree-ring analysis. Chinese Journal of Plant Ecology, 2012, 36(2): 117-125. |
| [21] | Gou X H, Shao X M, Wang Y J, Chen F H. The establishment of tree-ring chronology in east region of Qilian Mountains. Journal of Desert Research, 1999, 19(4): 364-367. |
| [22] | Chen C G, Peng H. Standing crops and productivity of the major forest-types at the Huoditang forest region of the Qinling Mountains. Journal of Northwest Forestry College, 1996, 11(Supplement): 92-102. |
| [23] | Wu X D. Tree Rings and Climate Change. Beijing: China Meteorological Press, 1990: 55, 71. |
| [24] | Wei F Y. Statistic Diagnose and Foreshadow Technology in Present Climate. Beijing: Meteorological Press, 2007: 37-40. |
| [25] | Ma Q Y. A study on the biomass of Chinese Pine forests. Journal of Beijing Forestry University, 1989, 11(4): 1-10. |
| [26] | Wu G, Feng Z W. Study on the social characteristics and biomass of the Pinus tabulaeformis forest systems in China. Acta Ecologica Sinica, 1994, 14(4): 415-422. |
| [27] | Wu G, Feng Z W. The sociological characteristics and biomass of stone Pine forests in China. Acta Ecologica Sinica, 1995, 15(3): 260-267. |
| [28] | Hou Y Y, Liu Q H, Yang H, Tian G L. Variation trends of China terrestrial vegetation net primary productivity and its responses to climate factors in 1982-2000. Chinese Journal of Applied Ecology, 2007, 18(7): 1546-1553. |
| [29] | Fang J Y. Forest productivity in China and its response to global climate change. Acta Phytoecologica Sinica, 2000, 24(5): 513-517. |
| [30] | Li D K, Fan J Z, Dong J F. Spatial and temporal patterns of net primary productivity from 1981 to 2000 in Shaanxi Province. Acta Botanica Boreali-Occidentalia Sinica, 2011, 31(9): 1873-1877. |
| [31] | Li J, Sun G N, Ren Z Y. Study on mass survey and value evaluation of land-vegetation′s primary productivity in Qinba Mountains in Shaanxi Province. Acta Ecologica Sinica, 2002, 22(12): 2254-2259. |
| [32] | Fei S M, He Y P, Chen X M, Jiang J M, Guo Z H. Quantitative features of populations of Pinus tabulaeformis and Pinus armandii regenerated following water damage at Qinling Mountains, China. Journal of Plant Ecology, 2008, 32(1): 95-105. |
| [33] | Fang J Y, Chen A P. Dynamic forest biomass carbon pools in China and their significance. Acta Botanica Sinica, 2001, 43(9): 967-973. |
| [34] | Li J F, Yuan Y J, You X R. Research and Application on Dendrohydrology. Beijing: Science Press, 2000: 11-85. |
| [35] | Tian Q H, Liu Y, Cai Q F, Bao G, Wang W P, Xue W L, Zhu W J, Song H M, Lei Y. The maximum temperature of May-July inferred from tree-ring in Funiu Mountain since 1874 AD. Acta Geographica Sinica, 2009, 64(7): 879-887. |
| [36] | Liu H B, Shao X M. Reconstruction of early-spring temperature of Qinling Mountains using tree-ring chronologies. Acta Geographica Sinica, 2003, 58(6): 879-884. |
| [37] | Feng X H, Cheng R M, Xiao W F, Wang R L, Wang X L, Liu Z B. The critical temperature to Huashan Pine (Pinus armandi) radial growth based on the daily mean temperature. Acta Ecologica Sinica, 2012, 32(5): 1450-1457. |
| [38] | Cao S J, Cao F X, Xiang W H. Tree-ring-based reconstruction of temperature variations from May to July since 1840 in Yanling county of Hunan province, China. Journal of Central South University of Forestry & Technology, 2012, 32(4): 10-14. |
| [39] | Gindl W, Grabner M, Rupert W. The influence of temperature on latewood lignin content in treeline Norway spruce compared with maximum density and ring width. Trees, 2000, 14(7): 409-414. |
| [40] | Sheppard P R, Graumlich L J, Conkey L E. Reflected-light image analysis of conifer tree rings for reconstructing climate. The Holocene, 1996, 6(1): 62-68. |
| [4] | 王斌, 刘某承, 张彪. 基于森林资源清查资料的森林植被净生产量及其动态变化研究. 林业资源管理, 2009, (1): 35-43. |
| [8] | 赵敏, 周广胜. 中国北方林生产力变化趋势及其影响因子分析. 西北植物学报, 2005, 25(3): 466-471. |
| [9] | 赵俊芳, 延晓冬, 贾根锁. 东北森林净第一性生产力与碳收支对气候变化的响应. 生态学报, 2008, 28(1): 92-102. |
| [14] | 滕菱, 彭少麟, 侯爱敏, 谢中誉. 长期气温波动对鼎湖山马尾松种群生产力的影响. 热带亚热带植物学报, 2001, 9(4): 284-288. |
| [15] | 程瑞梅, 封晓辉, 肖文发, 王瑞丽, 王晓荣, 杜化堂. 北亚热带马尾松净生产力对气候变化的响应. 生态学报, 2011, 31(8): 2086-2095. |
| [16] | 彭俊杰, 何兴元, 陈振举, 崔明星, 张先亮, 周长虹. 华北地区油松林生态系统对气候变化和CO2浓度升高的响应——基于BIOME-BGC模型和树木年轮的模拟. 应用生态学报, 2012, 23(7): 1733-1742. |
| [17] | 刘华, 雷瑞德, 侯琳, 王富群. 秦岭火地塘林区主要森林类型的碳储量和碳密度. 西北农林科技大学学报: 自然科学版, 2009, 37(3): 133-140. |
| [18] | 蒋冲, 王飞, 穆兴民, 李锐. 气候变化对秦岭南北植被净初级生产力的影响 (Ⅱ)——近52年秦岭南北植被净初级生产力. 中国水土保持科学, 2012, 10(6): 45-51. |
| [19] | 刘华, 侯琳, 雷瑞德. 秦岭火地塘林区油松和华山松林的空间分布格局及碳储量与碳密度研究. 中国生态农业学报, 2007, 15(1): 5-7. |
| [20] | 张远东, 刘彦春, 刘世荣, 张笑鹤. 基于年轮分析的不同恢复途径下森林乔木层生物量和蓄积量的动态变化. 植物生态学报, 2012, 36(2): 117-125. |
| [21] | 勾晓华, 邵雪梅, 王亚军, 陈发虎. 祁连山东部地区树木年轮年表的建立. 中国沙漠, 1999, 19(4): 364-367. |
| [22] | 陈存根, 彭鸿. 秦岭火地塘林区主要森林类型的现存量和生产力. 西北林学院学报, 1996, 11(S1): 92-102. |
| [23] | 吴祥定. 树木年轮与气候变化. 北京: 气象出版社, 1990: 55, 71. |
| [24] | 魏凤英. 现代气候统计诊断与预测技术. 北京: 气象出版社, 2007: 37-40. |
| [25] | 马钦彦. 中国油松生物量的研究. 北京林业大学学报, 1989, 11(4): 1-10. |
| [26] | 吴刚, 冯宗炜. 中国油松林群落特征及生物量的研究. 生态学报, 1994, 14(4): 415-422. |
| [27] | 吴刚, 冯宗炜. 中国主要五针松群落学特征及其生物量的研究. 生态学报, 1995, 15(3): 260-267. |
| [28] | 侯英雨, 柳钦火, 延昊, 田国良. 我国陆地植被净初级生产力变化规律及其对气候的响应. 应用生态学报, 2007, 18(7): 1546-1553. |
| [29] | 方精云. 中国森林生产力及其对全球气候变化的响应. 植物生态学报, 2000, 24(5): 513-517. |
| [30] | 李登科, 范建忠, 董金芳. 1981—2000年陕西省植被净初级生产力时空变化. 西北植物学报, 2011, 31(9): 1873-1877. |
| [31] | 李晶, 孙根年, 任志远. 陕西秦巴山区植被第一性生产物质量与价值量测评研究. 生态学报, 2002, 22(12): 2254-2259. |
| [32] | 费世民, 何亚平, 陈秀明, 蒋俊明, 郭志华. 秦岭水灾迹地油松和华山松更新种群数量特征. 植物生态学报, 2008, 32(1): 95-105. |
| [33] | 方精云, 陈安平. 中国森林植被碳库的动态变化及其意义. 植物学报, 2001, 43(9): 967-973. |
| [34] | 李江风, 袁玉江, 由希尧. 树木年轮水文学研究与应用. 北京: 科学出版社, 2000: 11-85. |
| [35] | 田沁花, 刘禹, 蔡秋芳, 包光, 王伟平, 薛文亮, 朱文杰, 宋慧明, 雷莺. 油松树轮记录的过去134年伏牛山5—7月平均最高温度. 地理学报. 2009, 64(7): 879-887. |
| [36] | 刘洪滨, 邵雪梅. 利用树轮重建秦岭地区历史时期初春温度变化. 地理学报, 2003, 58(6): 879-884. |
| [37] | 封晓辉, 程瑞梅, 肖文发, 王瑞丽, 王晓荣, 刘泽彬. 基于日均温度的华山松径向生长敏感温度研究. 生态学报, 2012, 32(5): 1450-1457. |
| [38] | 曹受金, 曹福祥, 项文化. 利用树木年轮研究湖南炎陵气温变化情况——1840年以来5—7月份气温变化情况重建. 中南林业科技大学学报, 2012, 32(4): 10-14. |