 2014, Vol. 34
2014, Vol. 34文章信息
- 杨丽丽, 王彦辉, 杜敏, 于澎涛, 郝佳, 李振华
- YANG Lili, WANG Yanhui, DU Min, YU Pengtao, HAO Jia, LI Zhenhua
- 六盘山典型森林伴随降水的总有机碳(TOC)通量变化特征
- The variation of total organic carbon (TOC) flux with rain water in regional forests of Liupan Mountains, China
- 生态学报, 2014, 34(21): 6297-6308
- Acta Ecologica Sinica, 2014, 34(21): 6297-6308
- http://dx.doi.org/10.5846/stxb201302060253
-
文章历史
- 收稿日期:2013-2-6
- 网络出版日期:2014-3-13






2. 中南林业科技大学林学院, 长沙 410004
2. College of Forestry, Central South University of Forestry and Technology, Changsha 410004, China
降水在携带各种元素进入森林生态系统的同时,随着与植被的接触,还会发生水量与元素浓度的明显变化,这些变化都对森林生长和其功能发挥具有重要作用[1, 2, 3]。碳元素虽然不是营养元素,但由于森林固碳功能日益受到重视,深入理解森林对碳循环的影响就显得格外重要。碳元素在森林生态系统内的流通和固定,在很大程度上也是森林生物地球化学循环的研究内容。维持碳循环与碳平衡是森林生态系统的重要服务功能,其大小与森林植被结构紧密相关,并受到森林水文过程的影响,突出表现在伴随水流而发生的碳浓度及通量的明显变化。当前的森林生态系统碳循环研究,还存在着很大不确定性[4],且大部分研究集中在植被、土壤以及以气体为载体的碳循环方面[5, 6],在以水为载体的生态系统碳循环研究还较少。
总有机碳(TOC) 指在水中以溶解或悬浮形态存在的全部有机碳,即可溶性有机碳(DOC) 和颗粒有机碳(POC)[7]。国外不少学者对温带和热带森林的大气降水、穿透降水、干流、土壤溶液和溪流水的DOC进行过报道[8, 9, 10],包括我国南方亚热带森林水文学过程中的TOC动态[11, 12, 13],然而在西北干旱半干旱地区研究还很少,限制着对森林生态功能的认识与管理。
本研究在宁夏六盘山区选择了六种典型森林类型,在生长季测定评价了伴随降水输入转化过程而发生的TOC通量变化,以便能为当地及类似地区的森林元素循环、养分平衡、固碳功能等相关过程的理解和定量评价提供科学依据。
1 研究区域与方法 1.1 研究区域和样地概况研究地点是六盘山自然保护区的香水河小流域,它处在黄土高原中西部的宁夏回族自治区南端,属黄河二级支流泾河的源头地区,海拔范围2060—2931 m,位于东经109°9′—109°30′,北纬35°15′—35°41′,属暖温带大陆性季风气候,年均气温5.8 ℃,年均降水量770.7 mm,多集中于6—9月。小流域内植被良好,森林覆盖率高达72.9%,主要为天然次生林,主要树种包括华山松(Pinus armandii)、辽东栎(Quercus liaotungesis)、白桦(Betula platyphlla)、红桦(Betula albosinensis)、山杨(Populus davidiana)等;其中人工林覆盖率为23.6%,主要为华北落叶松(Larix principis-rupprechtii)林和油松(Pinus tabulaeformis)林。山体阳坡分布着较多灌丛;高海拔(2700 m以上)地区则分布着亚高山草甸群落。土壤以灰褐土为主,还分布着一定的亚高山草甸土。土壤中砾石较多,成土母质为沙质泥岩、页岩、灰岩风化的残积物和坡积物。
选择了具代表性的华北落叶松人工林、华山松次生林、桦木次生林、辽东栎次生林和油松人工林建立了面积30 m×30 m的样地各1个,并在野李子(Prunus salicina)灌丛建立了1个20 m×20 m的样地。华北落叶松林、华山松林和桦木林样地分布于洪沟子流域,野李子灌丛样地分布于草沟子流域,辽东栎林和油松林样地分布于大南沟子流域。在华北落叶松林样地,林下的灌木和草本数量较少,灌木主要有华西箭竹(Fargesia nitida)、黄刺玫(Rosa hugonis)等,草本主要有华北苔草(Carex hancokiana)、蕨(Pteridium aquilum)等。华山松次生林中混生有少量的白桦和红桦,林下的灌木和草本数量较多,主要是忍冬(Lonicera japonica Thunb)、榛子(Corylus heterophylla)、峨嵋蔷薇(Rosa omeiensis)、冰草(Agropyron cristatum)、苔草(Carex)等。桦木天然次生林样地由白桦和少量红桦组成,林下的灌木和草本较多,主要有三裂绣线菊(Spiraea trilobata)、箭竹(Fargesia spathacea Franch)、榛子、小叶丁香(Sytinga microphylla)、忍冬、蕨、菝契(Smilax china L.)等。野李子灌木样地的林下草本数量较多,主要由冰草组成。辽东栎次生林样地的伴生树种较多,主要有白桦、山杨、少脉椴(Tilia paucicostata Maxim)等,其林下的灌木和草本较多,主要有甘肃山楂(Crataegus Kansuensis)、水栒子(Cotoneaster multiflorus)、柔毛绣线菊(Spriaea pubescens),刺五加(Eleutnerococcus giraldii)、箭竹等灌木以及苔草等。油松人工林样地为林分结构单一的纯林,林下草本层发育明显,种类较多,以东方草莓(Fragaria orientalis)、苔草、冰草为主要优势种。各试验地基本特征见表 1、表 2。
| 样地名称 Sample plot | 海拔Elevation/m | 坡向Slope aspect | 坡度Slope gradient/(°) | 坡位Slope position | 林龄Stand age/a | 林冠郁闭度Canopy density | 林分密度Stand density/(株/hm2) | 乔木平均胸径Mean tree DBH/cm | 乔木平均树高Mean tree height/m | 灌木层盖度Shrub coverage/% | 草本层盖度Herb coverage/% |
| 华北落叶松人工林 Larix principis-rupprechtii plantation | 2320 | 东南 | 30 | 坡下 | 23 | 0.8 | 1125 | 15.6 | 13.9 | 5 | 40 |
| 华山松次生林 Pinus armandii secondary forest | 2280 | 北 | 35 | 坡下 | 43 | 0.8 | 675 | 12.14 | 6.81 | 60 | 15 |
| 桦木次生林 Betula platyphylla secondary forest | 2380 | 北 | 30 | 坡中 | 45 | 0.6 | 725 | 18.3 | 9.34 | 65 | 10 |
| 野李子灌丛 Prunus salicina shrub | 2230 | 西 | 39 | 坡中 | 20 | 2.57 | 1.96 | 50 | 70 | ||
| 辽东栎次生林 Quercus liaotungensis secondary forest | 2080 | 东 | 34 | 坡下 | 41 | 0.6 | 625 | 16.16 | 9.23 | 60 | 10 |
| 油松人工林 Pinus tabulaeformis plantation | 2155 | 东南 | 7 | 坡下 | 32 | 0.65 | 2275 | 14.85 | 9.41 | 10 | 50 |
| 样地名称 Sample plot | 林冠层LAI Canopy LAI | 枯落物层厚度Thickness of humus/cm | 枯落物层重量Weight of humus/(t/hm2) | ||||||
| 未分解Un-decomposed | 半分解Half-decomposed | 已分解Decomposed | 合计 Total | 未分解Un-decomposed | 半分解Half-decomposed | 已分解Decomposed | 合计 Total | ||
| 华北落叶松人工林 Larix principis-rupprechtii plantation | 3.16 | 1.0 | 3.5 | 1.0 | 5.5 | 1.82 | 18.25 | 2.76 | 22.83 |
| 华山松次生林 Pinus armandii secondary forest | 2.94 | 0.7 | 2.2 | 0.5 | 3.4 | 1.54 | 12.68 | 10.51 | 24.73 |
| 桦木次生林 Betula platyphylla secondary forest | 3.32 | 0.8 | 3.3 | 2.0 | 6.1 | 1.72 | 13.19 | 10.95 | 25.87 |
| 野李子灌丛 Prunus salicina shrub | 2.7 | 0.7 | 3.4 | 2.62 | 3.34 | 5.96 | |||
| 辽东栎次生林Quercus liaotungensis secondary forest | 4.23 | 2.5 | 3.8 | 2.2 | 8.5 | 3.30 | 18.14 | 10.12 | 31.57 |
| 油松人工林 Pinus tabulaeformis plantation | 3.15 | 2.0 | 2.0 | 1.9 | 5.9 | 1.59 | 20.36 | 10.35 | 32.30 |
| *野李子灌丛样地的“乔木平均胸径”和“乔木平均树高”均为灌木数据 | |||||||||
|
样地名称 Sample plot | 土层深度 Soil depth/cm | 土壤容重 Soil bulk density /(g/cm3) | pH | 元素含量Element content/(g/100g) | |||||
| 有机COrganic C | 全N Total N | 全P Total P | 全K Total K | Ca | Mg | ||||
| 华北落叶松人工林 | 0—10 | 0.88 | 7.15 | 3.88 | 0.29 | 0.08 | 0.35 | 0.47 | 1.33 |
| Larix principis-rupprechtii | 10—20 | 1.00 | 7.35 | 3.45 | 0.26 | 0.09 | 0.38 | 0.65 | 1.33 |
| plantation | 20—40 | 1.02 | 7.32 | 2.97 | 0.25 | 0.08 | 0.35 | 0.68 | 1.32 |
| 40—60 | 1.11 | 7.45 | 2.83 | 0.25 | 0.09 | 0.31 | 1.17 | 1.27 | |
| 60—80 | 1.31 | 7.45 | 2.79 | 0.24 | 0.09 | 0.37 | 0.86 | 1.38 | |
| 80—100 | 1.45 | 7.57 | 2.02 | 0.18 | 0.09 | 0.33 | 0.85 | 1.40 | |
| 0—100 | 1.13 | 7.38 | 3.04 | 0.25 | 0.09 | 0.35 | 0.78 | 1.34 | |
| 华山松次生林 | 0—10 | 0.70 | 6.95 | 7.92 | 0.68 | 0.10 | 0.40 | 0.72 | 1.05 |
| Pinus armandii | 10—20 | 0.74 | 6.89 | 6.78 | 0.61 | 0.10 | 0.29 | 1.00 | 1.06 |
| secondary forest | 20—40 | 0.73 | 7.08 | 6.67 | 0.61 | 0.09 | 0.29 | 0.89 | 1.04 |
| 40—60 | 0.78 | 7.16 | 3.76 | 0.36 | 0.08 | 0.24 | 0.71 | 1.15 | |
| 60—80 | 0.94 | 7.25 | 3.18 | 0.33 | 0.08 | 0.19 | 0.72 | 1.16 | |
| 80—100 | 1.11 | 7.31 | 2.79 | 0.26 | 0.07 | 0.15 | 0.65 | 1.16 | |
| 0—100 | 0.83 | 7.11 | 5.18 | 0.47 | 0.09 | 0.26 | 0.78 | 1.10 | |
| 桦木次生林 | 0—10 | 0.69 | 6.95 | 8.54 | 0.64 | 0.10 | 0.27 | 1.32 | 0.98 |
| Betula platyphylla | 10—20 | 0.75 | 6.78 | 7.97 | 0.62 | 0.09 | 0.29 | 1.15 | 0.96 |
| secondary forest | 20—40 | 0.86 | 6.65 | 6.92 | 0.55 | 0.05 | 0.14 | 0.99 | 0.64 |
| 40—60 | 1.17 | 6.95 | 5.28 | 0.44 | 0.07 | 0.25 | 0.91 | 1.04 | |
| 60—80 | 1.24 | 6.94 | 4.85 | 0.39 | 0.07 | 0.24 | 0.94 | 1.08 | |
| 80—100 | 1.41 | 6.98 | 3.40 | 0.27 | 0.05 | 0.21 | 0.77 | 1.13 | |
| 0—100 | 1.02 | 6.88 | 6.16 | 0.49 | 0.07 | 0.23 | 1.01 | 0.97 | |
| 野李子灌丛 | 0—10 | 0.98 | 7.77 | 2.62 | 0.22 | 0.07 | 0.27 | 0.61 | 1.39 |
| Prunus salicina shrub | 10—20 | 1.07 | 7.72 | 2.20 | 0.20 | 0.06 | 0.25 | 0.49 | 1.38 |
| 20—40 | 1.02 | 7.87 | 2.02 | 0.18 | 0.07 | 0.23 | 0.66 | 1.44 | |
| 40—60 | 1.18 | 8.08 | 1.95 | 0.15 | 0.07 | 0.22 | 0.57 | 1.44 | |
| 60—80 | 0.96 | 8.28 | 1.80 | 0.11 | 0.06 | 0.20 | 1.86 | 1.65 | |
| 80—100 | 1.20 | 8.19 | 1.11 | 0.10 | 0.07 | 0.22 | 1.54 | 1.64 | |
| 0—100 | 1.07 | 7.99 | 1.95 | 0.16 | 0.07 | 0.23 | 0.96 | 1.49 | |
| 辽东栎次生林 | 0—10 | 0.74 | 7.22 | 4.21 | 0.34 | 0.04 | 0.14 | 0.72 | 0.93 |
| Quercus liaotungensis | 10—20 | 0.86 | 7.29 | 3.48 | 0.28 | 0.05 | 0.15 | 0.76 | 0.91 |
| secondary forest | 20—40 | 1.08 | 7.37 | 2.62 | 0.21 | 0.04 | 0.11 | 0.57 | 0.99 |
| 40—60 | 1.15 | 7.15 | 1.71 | 0.19 | 0.03 | 0.10 | 0.51 | 0.99 | |
| 60—80 | 1.15 | 7.34 | 1.91 | 0.19 | 0.05 | 0.13 | 0.49 | 1.14 | |
| 80—100 | 1.27 | 7.53 | 1.10 | 0.09 | 0.06 | 0.17 | 0.41 | 1.31 | |
| 0—100 | 1.04 | 7.32 | 2.51 | 0.22 | 0.05 | 0.13 | 0.58 | 1.04 | |
| 油松人工林 | 0—10 | 0.99 | 7.51 | 5.81 | 0.53 | 0.10 | 0.27 | 0.80 | 0.97 |
| Pinus tabulaeformis | 10—20 | 0.99 | 7.86 | 4.07 | 0.40 | 0.10 | 0.25 | 0.90 | 1.04 |
| plantation | 20—40 | 1.12 | 7.95 | 3.20 | 0.30 | 0.09 | 0.19 | 0.71 | 1.00 |
| 40—60 | 1.02 | 7.99 | 3.02 | 0.27 | 0.10 | 0.17 | 0.74 | 0.98 | |
| 60—80 | 1.00 | 8.08 | 2.74 | 0.25 | 0.09 | 0.17 | 0.62 | 0.98 | |
| 80—100 | 1.13 | 8.22 | 2.40 | 0.23 | 0.09 | 0.16 | 0.72 | 0.99 | |
| 0—100 | 1.04 | 7.94 | 3.54 | 0.33 | 0.09 | 0.20 | 0.75 | 0.99 | |
在林外开阔地放置一台自动气象站(美国,Weatherhawk公司),配有计量精度为1 mm的翻斗式雨量计,2011年5—10月连续观测降水量和降水过程;同时配有一台虹吸式自计雨量计和一个标准雨量筒,配合测定林外降雨量及降雨过程。在林外布设了3个内径20 cm的自制雨量筒,用于收集林外降水的样品。
为收集穿透水,在六种森林样地内,根据上方林冠(以及高大灌木)的遮蔽程度,选择代表性地点,在距地面20 cm的高度,分别布设6—12个雨量筒,每次雨后测定水量,计算其算术平均值,作为样地整个冠层下的穿透水量。
为测定干流,按林木径级和树冠中等标准,每个样地内选择5—6棵林木,将直径2.0 cm的聚乙烯塑料软管沿中缝剖开,从树干2 m高处自上向下蛇形缠绕于树干上,塑料管与水平面角度呈30°,用玻璃胶密封树干与胶管间的空隙,再将胶管引入密封口的塑料容器。每次雨后及时测水量,并依据林冠投影面积换算为单株和林分的干流深。
为收集穿过枯落物层的渗漏水,在尽量不破坏原来枯落物层次结构的条件下,每个样地内选择5个样点,把0.5 m×0.5 m的原状枯落物整体移放到塑料膜上。每次降雨后将渗过枯落物层的雨水引入塑料盆中收集,测定枯落物渗漏水量,并保存水样以测定元素浓度。
为测定主根系层的土壤渗漏水,在华北落叶松林和华山松林样地内,随机布设7个自制的内径20 cm的蒸渗仪,蒸渗仪内装填了未扰动的林地土壤,其土层厚度为30 cm,筒内保留采集原状土柱地点的原有枯落物层及可能生长的草本和小灌木等林下植被。在每次降雨停止后,测定蒸渗仪底筒收集的土壤渗漏水量,并采集化学分析的水样。
每场降雨后,将多点采集的林外降水以及各样地内多点采集的穿透水、干流、枯落物渗漏水和土壤渗漏水混合,分别形成混合水样;另外,在洪沟、草沟、大南沟小流域分别采集溪水水样。保留水样的体积为500 mL,存储于冰柜内,及时送回北京的实验室进行处理、保存和分析。在2011生长季(5月24日—10月20日),共测定22次降雨事件,取得528瓶水样。
1.2.2 水样处理和化学分析在野外用定性滤纸过滤水样后,立即用上海雷磁公司出产的pH计测定其pH值及电导率。在清华大学分析实验室,采用燃烧氧化-非分散红外吸收法,测定总有机碳(TOC)浓度。
1.2.3 TOC浓度计算某类水样在生长季中的TOC浓度平均值是各降雨事件中相应水量的加权计算值(式(1)):
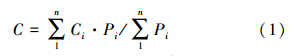
林下降水包括林冠穿透水和干流,故其数值等于穿透水和干流的深度之和,其TOC浓度平均值是单独测定的穿透水与干流的TOC浓度的水量加权平均值。
本研究中,将森林生态系统复杂结构简化为3个作用层,即冠层(林冠层+灌草层)、枯落物层、主根系层土壤层(0—30 cm),则伴随降水转化通过各作用层的TOC通量依下面式(2)计算:

在降水通过各个作用层时,会有TOC的吸附、交换、洗脱等作用过程,从而在发生水量变化的同时也发生TOC浓度变化,综合形成各作用层对TOC通量的影响。依据下面式(3),计算通过某作用层的TOC通量(Fi)相对其相邻上作用层输入的TOC通量(Fj)的变化(ΔF),即该作用层对上面相邻作用层的影响:
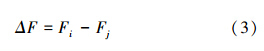
在香水河小流域的2011生长季(5月24日—10月20日)研究期间,共观测到22次降雨,总降雨量为724.3 mm。其月际分配差异较大(表 3):在5月份降雨量为0,6月份降雨量不大,到7月份开始急剧增加,8月份比7月份稍有降低,9月份由于暴雨出现而达到最大,在10月份则迅速减少。
| 样地名称 Sample plot | 月份 Months | 降雨量 Precipitation/mm | 截留量 Interception | 干流量 Stemflow | 穿透降雨 Throughfall | 林下降水 Rainfall under canopy | 枯落物渗漏水 Humus leakage/mm | 土壤渗漏水 Soil leakage/mm | ||||
| /mm | /% | /mm | /% | /mm | /% | /mm | /% | |||||
| *观测从5月24日开始,10月20日结束 | ||||||||||||
| 华北落叶松人工林 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.2 | ||||
| Larix principis-rupprechtii | 6 | 83.3 | 19.2 | 23.02 | 0.1 | 0.09 | 64.0 | 76.88 | 64.1 | 76.98 | 34.7 | 32.4 |
| plantation | 7 | 188.0 | 37.9 | 20.17 | 0.5 | 0.29 | 149.5 | 79.54 | 150.1 | 79.83 | 104.7 | 23.3 |
| 8 | 171.1 | 25.0 | 14.60 | 0.6 | 0.34 | 145.5 | 85.06 | 146.1 | 85.40 | 112.7 | 107.8 | |
| 9 | 232.3 | 36.6 | 15.76 | 1.0 | 0.44 | 194.7 | 83.80 | 195.7 | 84.24 | 135.6 | 59.4 | |
| 10 | 49.6 | 6.2 | 12.58 | 0.3 | 0.65 | 43.0 | 86.78 | 43.4 | 87.42 | 19.5 | 21.1 | |
| 5—10 | 724.3 | 124.9 | 17.25 | 2.6 | 0.35 | 596.8 | 82.40 | 599.4 | 82.75 | 407.2 | 244.2 | |
| 华山松次生林 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.4 | ||||
| Pinus armandii | 6 | 83.3 | 14.3 | 17.13 | 0.3 | 0.39 | 68.7 | 82.48 | 69.0 | 82.87 | 40.8 | 32.8 |
| secondary forest | 7 | 188.0 | 40.1 | 21.33 | 0.4 | 0.22 | 147.5 | 78.45 | 147.9 | 78.67 | 100.5 | 29.4 |
| 8 | 171.1 | 14.7 | 8.58 | 0.3 | 0.20 | 156.1 | 91.21 | 156.4 | 91.42 | 119.7 | 123.3 | |
| 9 | 232.3 | 24.9 | 10.70 | 0.5 | 0.22 | 206.9 | 89.08 | 207.4 | 89.30 | 143.4 | 61.0 | |
| 10 | 49.6 | 8.4 | 16.98 | 0.2 | 0.31 | 41.0 | 82.71 | 41.2 | 83.02 | 20.0 | 29.0 | |
| 5—10 | 724.3 | 102.3 | 14.13 | 1.8 | 0.24 | 620.2 | 85.63 | 622.0 | 85.87 | 424.3 | 275.9 | |
| 桦木次生林 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |||||
| Betula platyphylla | 6 | 83.3 | 17.8 | 21.33 | 0.1 | 0.11 | 65.4 | 78.56 | 65.5 | 78.67 | 34.8 | |
| secondary forest | 7 | 188.0 | 35.1 | 18.66 | 0.3 | 0.17 | 152.6 | 81.17 | 152.9 | 81.34 | 95.9 | |
| 8 | 171.1 | 31.6 | 18.47 | 0.4 | 0.22 | 139.1 | 81.31 | 139.5 | 81.53 | 101.1 | ||
| 9 | 232.3 | 52.4 | 22.54 | 0.5 | 0.20 | 179.5 | 77.26 | 179.9 | 77.46 | 115.8 | ||
| 10 | 49.6 | 8.8 | 17.81 | 0.1 | 0.11 | 40.7 | 82.08 | 40.8 | 82.19 | 20.7 | ||
| 5—10 | 724.3 | 145.6 | 20.11 | 1.3 | 0.18 | 577.3 | 79.71 | 578.7 | 79.89 | 368.3 | ||
| 野李子灌丛 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |||||
| Prunus salicina | 6 | 83.3 | 11.6 | 13.98 | 0.2 | 0.21 | 71.5 | 85.81 | 71.7 | 86.02 | 45.2 | |
| shrub | 7 | 188.0 | 23. 4 | 12.44 | 1.2 | 0.63 | 163.4 | 86.93 | 164.6 | 87.56 | 136.1 | |
| 8 | 171.1 | 18.6 | 10.86 | 1.4 | 0.81 | 151.1 | 88.33 | 152.5 | 89.14 | 139.1 | ||
| 9 | 232.3 | 28.2 | 12.14 | 1.5 | 0.66 | 202.6 | 87.20 | 204.1 | 87.86 | 185.9 | ||
| 10 | 49.6 | 4.4 | 8.91 | 0.4 | 0.86 | 44.8 | 90.23 | 45.2 | 91.09 | 18.7 | ||
| 5—10 | 724.3 | 86.2 | 11.91 | 4.7 | 0.65 | 633.4 | 87.44 | 638.1 | 88.09 | 525.0 | ||
| 辽东栎次生林 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |||||
| Quercus liaotungensis | 6 | 83.3 | 15.8 | 19.02 | 0.2 | 0.30 | 67.2 | 80.69 | 67.5 | 80.98 | 29.2 | |
| secondary forest | 7 | 188.0 | 44.4 | 23.61 | 0.7 | 0.40 | 142.9 | 76.00 | 143.6 | 76.39 | 119.3 | |
| 8 | 171.1 | 41.8 | 24.43 | 0.5 | 0.31 | 128.8 | 75.26 | 129.3 | 75.57 | 46.0 | ||
| 9 | 232.3 | 42.8 | 18.43 | 0.7 | 0.28 | 188.8 | 81.29 | 189.5 | 81.57 | 118.3 | ||
| 10 | 49.6 | 11.3 | 22.84 | 0.2 | 0.37 | 38.1 | 76.80 | 38.3 | 77.16 | 18.1 | ||
| 5—10 | 724.3 | 156.2 | 21.56 | 2.4 | 0.33 | 565.8 | 78.11 | 568.1 | 78.44 | 330.8 | ||
| 油松人工林 | 5 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | 0.0 | |||||
| Pinus tabulaeformis | 6 | 83.3 | 26.6 | 31.94 | 0.8 | 1.02 | 55.8 | 67.04 | 56.7 | 68.06 | 34.6 | |
| plantation | 7 | 188.0 | 43.0 | 22.87 | 2.5 | 1.35 | 142.5 | 75.78 | 145.0 | 77.13 | 125.6 | |
| 8 | 171.1 | 37.7 | 22.04 | 2.1 | 1.22 | 131.3 | 76.74 | 133.4 | 77.96 | 50.1 | ||
| 9 | 232.3 | 37.8 | 16.29 | 2.7 | 1.18 | 191.7 | 82.53 | 194.5 | 83.71 | 170.9 | ||
| 10 | 49.6 | 11.2 | 22.56 | 0.5 | 0.98 | 37.9 | 76.46 | 38.4 | 77.44 | 21.6 | ||
| 5—10 | 724.3 | 156.3 | 21.59 | 8.7 | 1.20 | 559.3 | 77.21 | 568.0 | 78.41 | 402.6 | ||
降水进入森林后进行了一连串的再分配,形成了冠层截留、直接穿透雨、干流、枯落物渗漏水和土壤渗漏水。因植被特征差异,各样地的降雨分配存在差异。由表 3可知,2011年生长季(5月24日—10月20日)的截留量(率)在油松人工林最大,达到156.3 mm(21.59%),其次为辽东栎次生林的156.2 mm(21.56%)、桦木次生林的145.6 mm(20.11%)、华北落叶松人工林的124.9 mm(17.25%)、华山松次生林的102.3 mm(14.13%)、野李子灌丛的86.2 mm(11.91%)。目前国内研究的林冠截留率大多介于10%—35%[14, 15],但一些地区有时会超过50%[16, 17]。本研究中截留率接近于徐丽宏等[18]2004—2005年生长季对六盘山主要植被类型的林冠层截留结果。
各主要植被类型的干流量月统计表明,干流量随降雨量增大而升高,但干流率均不高。影响干流的因素很多,包括树种、树干粗细、直立程度、树皮干燥程度等。在各样地的干流量(率)中,以油松人工林最大,达到8.7 mm(1.20%);野李子灌丛次之,达到4.7 mm(0.65%);桦木次生林最小,为1.3 mm(0.18%)。
由表 3可看出,各样地的月穿透水量随林外降水量增大而增大,其在各水文分量中都是最大。各样地的穿透水量(mm)依次为野李子灌丛(633.4)>华山松林(620.2)>华北落叶松林(596.8)>桦木林(577.3)>辽东栎林(565.8)>油松林(559.3),分别占同期降水量的87.44%、85.63%、82.40%、79.71%、78.11%、77.21%。
枯落物渗漏水大小与各林分样地的枯落物数量和分解状况等有关,由大到小依次为野李子灌丛(525.0 mm)>华山松林(424.3 mm)>华北落叶松林(407.2 mm)>油松林(402.6 mm)>桦木林(368.3 mm)>辽东栎林(330.8 mm),并随各月降水量增大而增大。
土壤渗漏水与立地条件、植被生长耗水、土壤机械组成等多种因素密切相关。基于本文测定,华北落叶松林和华山松林在2011生长季的土壤(0—30 cm)渗漏水分别为244.2和275.9 mm。
2.2 各水文分量的TOC浓度变化从表 4可看出,在降水通过森林各层次的过程中,TOC浓度均发生了改变。雨季大气降水的TOC平均浓度为(7.34±0.098)mg/L;由于降水通过林冠时和树体接触发生淋洗作用,TOC浓度明显增加,并在干流中达到最大。通过各样地枯落物层的渗漏水的TOC浓度比林冠穿透水继续明显增加。在经过主根系土壤层后,由于土壤过滤作用,华北落叶松林和华山松林的土壤渗漏水TOC浓度均低于其枯落物渗漏水TOC浓度。在本文研究的森林流域,溪水主要是雨水入渗到土壤后形成的壤中流和基流组成的,溪水的TOC浓度远低于林外降水,说明土壤具有突出的TOC过滤作用。需要说明的是,表 4中假设林外降水的TOC平均浓度对所有样地都一样,各样地对应的溪水TOC浓度为各样地所在子流域的溪水实测均值。
| 水文分量 Water components | 华北落叶 松人工林 Larix principis- rupprechtii plantation | 华山松次生林 Pinus armandii secondary forest | 桦木次生林 Betula platyphylla secondary forest | 野李子灌丛 Prunus salicina shrub | 辽东栎次生林 Quercus liaotungensis secondary forest | 油松人工林 Pinus tabulaeformis plantation |
| 林外降水Precipitation | 7.34±0.098 | 7.34±0.098 | 7.34±0.098 | 7.34±0.098 | 7.34±0.098 | 7.34±0.098 |
| 穿透水Throughfall | 21.92±0.200 | 17.11±0.209 | 11.47±0.084 | 12.41±0.259 | 11.05±0.124 | 16.23±0.169 |
| 干流Stemflow | 56.89±0.285 | 25.68±0.229 | 23.57±0.131 | 18.12±0.171 | 21.04±0.146 | 37.90±0.436 |
| 枯落物渗漏水Humus leakage | 22.29±0.304 | 24.72±0.210 | 22.90±0.213 | 24.64±0.217 | 23.94±0.206 | 28.55±0.345 |
| 土壤渗漏水Soil leakage | 17.63±0.200 | 24.04±0.175 | ||||
| 溪水Stream water | 3.88±0.087 | 3.88±0.087 | 3.88±0.087 | 3.45±0.069 | 3.59±0.057 | 3.59±0.057 |
降水转化中的TOC浓度(mg/L)变化对华北落叶松林表现为林外降水(7.34)<土壤渗漏水(17.63)<穿透水(21.92)<枯落物渗漏水(22.29)<干流(56.89),对华山松林表现为林外降水(7.34)<穿透水(17.11)<土壤渗漏水(24.04)<枯落物渗漏水(24.72)<干流(25.68),对桦木林为林外降水(7.34)<穿透水(11.47)<枯落物渗漏水(22.9)<干流(23.57),对油松林为林外降水(7.34)<穿透水(16.23)<枯落物渗漏水(28.55)<干流(37.9),对野李子灌丛为林外降水(7.34)<穿透水(12.41)<干流(18.12)<枯落物渗漏水(24.64),对辽东栎林为林外降水(7.34)<穿透水(11.05)<干流(21.04)<枯落物渗漏水(23.94)。
大气降水进入森林后TOC浓度增加,是因降水对叶面或树皮上有机碳的淋洗或淋溶[10],因而冠层结构和组成是重要影响因素。在测定的6种森林样地中,穿透水的TOC浓度(mg/L)为华北落叶松林(21.92)>华山松林(17.11)>油松林(16.23)>灌丛(12.41)>桦木林(11.47)>辽东栎林(11.05),表现为针叶林的TOC淋洗或淋溶能力较阔叶林强,这不同于罗艳等[11]对鼎湖山3种主要林型总有机碳浓度的研究结果,这可能与具体林分的冠层叶量有关,也可能与不同地点的降雨特征差异有关,还有待深入定量研究。
降水在树皮中停留时间和树皮形态及可溶性TOC含量是影响干流TOC浓度的主要因素[2, 10]。针叶树(尤其华北落叶松和油松)树皮一般较粗糙,并有分层结构,故相对其它树种可将雨水滞留更长时间,所以其干流TOC浓度明显高于阔叶树种,也高于本身的其他水文分量。华山松的干流TOC浓度在3个针叶树中最低,是因其树皮光滑且量少。对其它3个阔叶树种,因其树皮较光滑,滞水能力相对较差,所以干流TOC浓度也低,与枯落物渗漏水相近。
枯落物渗漏水的TOC浓度整体而言大于穿透水,主要是因对枯落物的淋洗淋溶造成的。不同样地的枯落物渗漏水TOC浓度增加程度存在一些差异,这与枯落物的厚度和组成有关。
2.3 林内各作用层的TOC通量由表 5可知,2011年生长季林外降水携带的TOC通量为53.17 kg/hm2。各林分样地的林下降水(穿透水和干流)携带的TOC通量均高于林外降水,说明林冠层具有TOC的净淋出作用。各样地的生长季林下降水TOC通量的大小(kg/hm2)排序为华北落叶松林(132.28)>华山松林(106.56)>油松林(94.10)>野李子灌丛(79.49)>桦木林(66.52)>辽东栎林(63.01)。
| 水文分量 Water components | 华北落叶松 人工林 Larix principis- rupprechtii plantation | 华山松次生林 Pinus armandii secondary forest | 桦木次生林 Betula platyphylla secondary forest | 野李子灌丛 Prunus salicina shrub | 辽东栎次生林 Quercus liaotungensis secondary forest | 油松人工林 Pinus tabulaeformis plantation |
| 林外降水Precipitation | 53.17 | 53.17 | 53.17 | 53.17 | 53.17 | 53.17 |
| 穿透水Throughfall | 130.82 | 106.11 | 66.22 | 78.63 | 62.51 | 90.80 |
| 干流Stemflow | 1.46 | 0.45 | 0.31 | 0.86 | 0.50 | 3.30 |
| 林下降水Rainfall under canopy | 132.28 | 106.56 | 66.52 | 79.49 | 63.01 | 94.10 |
| 枯落物渗漏水Humus leakage | 90.76 | 104.90 | 84.35 | 129.35 | 79.21 | 114.93 |
| 土壤层渗漏水Soil leakage | 43.04 | 66.33 |
各样地枯落物渗漏水的生长季TOC通量(kg/hm2)排序为野李子灌丛(129.35)>油松林(114.93)>华山松林(104.90)>华北落叶松林(90.76)>桦木林(84.35)>辽东栎林(79.21)。
经过主根系层土壤后的渗漏水的生长季TOC通量(kg/hm2)为华山松林(66.33)>华北落叶松(43.04),这可能和华山松林土壤结构比较疏松和大孔隙较多有关。
2.4 各作用层间TOC通量的差别在降水接触植被表面、枯落物和土壤时,都会发生TOC的淋失、吸附和吸收等过程,加之水量变化,造成了TOC通量在各作用层的差别(表 6)。
| 比较的作用层 Layers to be compared | 华北落叶松人工林 Larix principis-rupprechtii plantation | 华山松次生林 Pinus armandii secondary forest | 桦木次生林 Betula platyphylla secondary forest | 野李子灌丛 Prunus salicina shrub | 辽东栎次生林 Quercus liaotungensis secondary forest | 油松人工林 Pinus tabulaeformis plantation |
| 林下降水-林外降水 Rainfall under canopy-Precipitation | 77.65 | 52.94 | 13.05 | 25.46 | 9.34 | 37.63 |
| 枯落物渗漏水-林下降水 Humus leakage-Precipitation | -41.52 | -1.67 | 17.83 | 49.86 | 16.19 | 20.83 |
| 土壤层渗漏水-枯落物渗漏水 Soil leakage-Humus leakage | -47.71 | -38.57 | ||||
| 土壤层渗漏水-林外降水 Soil leakage-Precipitation | -10.12 | 13.16 |
林冠层表面积巨大,一般会有降水在林冠层的TOC净淋洗作用[19],包括对树体表面积累的干沉降的洗脱作用,这导致树冠对林地TOC输入的“源”作用。在本研究中,各林分样地的林下降水携带TOC通量(kg/hm2)相对林外降水均有不同程度的增加,其中华北落叶松林增幅最大(77.65),华山松林(52.94)和油松林(37.63)次之,表明针叶林冠层(针叶、枝条和和树皮)的TOC净淋出作用很强;相比之下,桦木林和辽东栎林的TOC通量仅增大了13.05和9.34,说明阔叶林冠层的TOC净淋出作用不强;野李子灌丛的TOC通量增幅也不是很大(25.46)。
枯落物层渗漏水与林下降水携带的TOC通量之差,可指示枯落物层对输入林地的TOC通量的作用,其值在华北落叶松林和华山松林内均为负值,说明其枯落物层具有TOC的净吸附或净储存的“汇”作用,也说明这两种森林内部的TOC增加主要源于雨水对冠层的淋洗。而在其他四个样地的枯落物层渗漏水的TOC通量均大于林下降水,说明这几种森林样地的枯落物层具有净增加林地TOC输入通量的“源”作用。
本文仅测定了华北落叶松林和华山松林2种森林样地的主根系层土壤渗漏水,其携带的TOC通量(kg/hm2)相对于枯落物渗漏水均降低了,其减幅比较接近,分别为47.71和38.57,说明土壤层对输出主根系层的TOC通量具有净吸附固定的“汇”作用。
林地主根系层土壤渗漏水的TOC通量与林外降水TOC通量的差,可以表明森林生态系统对林外降水输入的TOC通量的作用。在本研究中,表现为华北落叶松林生态系统(仅考虑到主根系层的0—30 cm)对降水输入TOC通量有净固定作用,其值为10.12 kg/hm2;华山松林生态系统对降水输入TOC通量有净淋出作用,其值为13.16 kg/hm2。
3 讨论和结论在宁夏六盘山地区,林外降水的TOC浓度(7.34 mg/L)明显高于一些其他地区的报道值(表 7),如广东鼎湖山(2.4 mg/L)[12]、中国台湾关刀溪(4.7 mg/L)和一些国外温带森林研究结果(1.0—2.9 mg/L)[8, 9, 10, 20, 21, 22, 23, 24, 25],这可能和地处干旱地区的六盘山降水量少有关。
| 试验地点 Site | 测定项目 Items | 森林类型 Forest type | 林外降水 Precipitation | 穿透水 Throughfall | 干流 Stemflow | 土壤渗漏水 Soil leakage | 溪水 tream ater | 参考文献References | |
| 0—15 cm | 15—30 cm | ||||||||
| *DOC:可溶性有机碳 | |||||||||
| 美国新罕布什尔州 Hubbard Brook,New Hampshire,USA | DOC | 硬木阔叶林 | 1.1 | 33.9 | 5.9 | 3.0 | 3.1 | [21] | |
| 加拿大魁北克省希莱尔 Mont St,Hilaire,Quebec | DOC | 硬木阔叶林 | 2.0 | 12.3 | 47.6 | 18.0 | [22] | ||
| 美国华盛顿奥林匹克公园 Olympic National Park, Washington,USA | DOC | 针叶林 | 1.5±1.1 | 10.5±17.9 | 25.5±9.3 | [23] | |||
| 硬木阔叶林 | 1.5±1.1 | 7.3±5.5 | 26.3±16.3 | 9.0±4.6 | 2.9±1.1 | 1.0±1.0 | |||
| 波多黎各卢科伊罗山 Luquillo Mountains,Puerto Rico | DOC | 1.0 | 6.2 | 1.9 | [10] | ||||
| 美国威斯康星州下威斯康星河 Lower Wisconsin River Valley,Wisconsin,USA | DOC | 北美赤松 | 2.9±0.2 | 11.8±0.9 | [24] | ||||
| 新西兰华兰 Westland,New Zealand | DOC | 榉木林 | 1.4±0.7 | 16.0±12.3 | 35.6±68.6 | 55.7±37.6 | 4.5±1.7 | [25] | |
| 日本熊本县 | DOC | 针叶林 | 1.0 | 4.3 | 11.5 | [9] | |||
| Kumamoto,Japan | 硬木阔叶林 | 1.0 | 3.1 | 7.1 | |||||
| 美国马萨诸塞州哈佛森林 | DOC | 针叶林 | 1.8±0.7 | 24.7±3.2 | [8] | ||||
| Harvard Forest, Massachusetts,USA | 硬木阔叶林 | 1.8±0.7 | 29±6.9 | ||||||
| 中国台湾关刀溪 | DOC | 杉木林 | 4.7 | 7.0 | 30.8 | 8.8 | 7.7 | 2.7 | [20] |
| Guandaushi,Taiwan,China | 次生阔叶 | 4.7 | 9.9 | 10.0 | 15.5 | 13.5 | |||
| 天然阔叶林 | 4.7 | 8.3 | 7.2 | 21.3 | 11.0 | ||||
| 广东鼎湖山 | TOC | 马尾松林 | 2.4±1.2 | 12.9±7.9 | 52.1±25.6 | 12.8±4.4 | 12.1±6.8 | 5.1±5.3 | [12] |
| 季风林 | 2.4±1.2 | 14.6±12.2 | 18.6±9.7 | 10.8±8.3 | 5.5±1.5 | 2.0±0.5 | |||
| 混交林1 | 2.4±1.2 | 14.2±13.6 | 21.5±10.7 | 13.6±4.3 | 10.2±5.7 | 1.8±0.7 | |||
| 混交林2 | 2.4±1.2 | 13.2±13.1 | 23.9±11.8 | 11.9±5.3 | 5.4±2.9 | ||||
由于雨水对树体表面的淋洗[26]和冲刷[10],在林外降水转为穿透水和干流过程中,TOC浓度明显增大。对六盘山所有测试树种,TOC浓度均在干流中达到最高,变化在18.12—56.89 mg/L,略高于其它地区的测定值范围(7.1—52.1 mg/L)[8, 9, 10, 11, 20, 21, 22, 23, 24, 25]。六盘山主要森林类型穿透降水的TOC浓度变化在11.05—21.92 mg/L,平均15.03 mg/L,稍高于广东鼎湖山(12.9—14.6 mg/L)[12],高于中国台湾中部关刀溪(7.0—9.9 mg/L)[20],但在其他地区的变化范围(3.1—33.9 mg/L)之内。这些地区差别,可能既与树种形态特征有关,也与区域降水特征有关[9]。
在林下降水转为枯落物层渗漏水过程中,其TOC浓度变化与树种特征、枯落物数量和组成、生长物候期等均有关[27, 28]。六盘山主要森林类型枯落物渗漏水的TOC浓度变化在22.29—28.55 mg/L,相互差别不大,但整体上低于广东鼎湖山的马尾松林(31.7)、针阔混交林(28.3)和季风林(25.5)[13],这可能与六盘山地区温度低枯落物分解慢有关。
土壤渗漏水TOC含量与土壤有机质含量密切相关,并一般随土壤酸度增加而降低[29]。在六盘山典型森林植被中,土壤渗漏水(0—30 cm)的TOC浓度(mg/L)变化在17.63—24.04,高于广东鼎湖山(5.4—12.1)[12]和中国台湾关刀溪(7.7—11.0)[20],这可能与南方土壤有机质含量低和土壤严重酸化导致有机质分解缓慢有关[30]。
在六盘山香水河小流域几个子流域的溪水中,TOC浓度较接近,变化在3.45—3.88 mg/L,显著低于从林外降水到土壤渗漏水的各转换环节,说明深层土壤的TOC吸附拦截作用不容忽视,这与国内外其他地方研究结果相同[10, 20, 21, 23, 25],并处在相近的数值范围内。
本文在六盘山的研究表明,随着雨水穿过林冠和植被表面接触,发生了生长季内的TOC通量变化,并在不同树种林分之间具有显著差别。相对于林外降水的TOC通量(53.17 kg/hm2),由穿透降水和干流组成的林下降水的TOC通量在所有林分样地都增大了,表明所有森林类型的林冠层均有TOC净淋出作用。但整体看来,林冠的TOC净淋出作用在针叶林内很大,在阔叶林内较弱,其原因可能是阔叶树的叶面光滑因此与针叶树林冠相比难以拦截和储存大量干沉降,或阔叶树的叶片革质化程度高从而被雨水浸出TOC较少等,其具体原因有待继续深入研究。
生长季内六盘山不同林分枯落物渗漏水的TOC通量变化在79.21—129.35 kg/hm2,平均为100.58 kg/hm2。在研究的几种森林样地中,仅华北落叶松人工林和华山松次生林的枯落物渗漏水的TOC通量低于林下降水的TOC通量,说明这两种森林样地的枯落物层起着净固定TOC的“汇”作用;而其他四种森林样地的枯落物层TOC通量高于林下降水,说明其枯落物层对TOC通量起着净输出的“源”作用。
由于工作量原因,本文在6个样地中仅测定了华北落叶松林和华山松林的主根系层土壤(0—30 cm)渗漏水的生长季TOC通量,二者数值分别为43.04和66.33 kg/hm2,它们都大大低于枯落物渗漏水的TOC通量,说明土壤层起着最重要的吸收固定TOC的“汇”作用;溪水中的TOC含量降到了3.45—3.88 mg/L,仅为林外降水TOC含量的一半左右,这说明土壤层起着过滤和固定TOC的巨大作用。
林内不同水文学过程的TOC浓度和通量变化,可反映森林内部伴随雨水转化发生的碳转运过程。可以认为,在六盘山地区,林冠层具有对林外降水输入TOC通量的净增加(淋溶和淋洗)作用;枯落物层具有对林下降水输入TOC通量的减少或增加作用,表现为某种程度的缓冲作用,即在林下降水TOC通量低时增加和高时减少,同时这个缓冲作用可能和不同森林类型的枯落物的数量、性质及分解程度等有关,其具体作用机制和不同因素的贡献都还有待深入探讨;但主根系层(土壤层)可大大降低枯落物层渗漏水输入的TOC通量,起着净固定作用。
综合来看,林外降水为森林生态系统输入了相当数量的TOC(“源”作用)。对于森林生态系统内随降水发生的TOC转化而言,作为“TOC活库”的林冠层具有增加TOC通量的“源”作用,作为“TOC死库”的枯落物层可起增加TOC通量的“源”作用或减少TOC通量的“汇”作用。由于林地土壤层(0—30 cm土层渗漏水)的TOC输出通量低于林下降水和枯落物渗漏水的TOC输入通量,因而说森林土壤能发挥固定TOC的“库”作用。
| [1] | Lindberg S E, Lovett G M, Richter D D, Johnson D W. Atmospheric deposition and canopy interactions of major ions in a forest. Science, 1986, 231(4734): 141-145. |
| [2] | Puckett L J. Estimates of ion sources in deciduous and coniferous through-fall. Atmospheric Environment, 1990, 24: 545-556. |
| [3] | Ma X H. Forest Hydrology. Beijing: China Forestry Press, 1993, 43-45. |
| [4] | Yang X, Wang M X. Reviews of several aspects of terrestrial carbon cycling. Progress In Geography, 2001,16(3): 427-435. |
| [5] | Melillo J M, Mcguire A D, Kicklighter D W, Moore B, Vorosmarty C J, Schloss A L. Global climate change and terrestrial net primary production. Nature, 1993, 363(6426): 234-240. |
| [6] | Raich J W, Schlesinger W H. The Global carbon dioxide flux in soil respiration and its relationship to vegetation. Tellus B, 1992, 44(2): 81-99. |
| [7] | Greenberg A E. Standard Methods for Examination of Waters and Watersheds. Washington, DC: American Public Health Association, 1992, 64(3/4): 51-59. |
| [8] | Currie W S, Aber J D, Mcdowell W H, Boone R D. Vertical transport of dissolved organic C and N under long-term N amendments in pine and hardwood forests. Biogeochemistry, 1996, 35(3): 471-505. |
| [9] | Inagaki M, Sakai M, Ohnuki Y. The effects of organic carbon on acid rain in a temperate forest in Japan. Water Air Soil Pollution, 1995, 85(4): 2345-2350. |
| [10] | McDowell W H. Internal nutrient fluxes in a Puerto Rican rain forest. Journal of Tropical Ecology, 1998, 14(4): 521-536. |
| [11] | Luo Y, Zhou G Y, Zhang D Q, Guan L L, Ouyang X J, Chu G W.Study on the concentration of total organic carbon in the forest hydrological processes of three main forest types in Dinghushan during a rain season. Acta Ecologica Sinica, 2004, 24(12): 2973-2978. |
| [12] | Yin G C, Zhou G Y, Zhang D Q, Wang X, Chu G W, Liu Y. Dynamics of total organic carbon (TOC) in hydrological processes in coniferous and broad-leaved mixed forest of Dinghushan. Chinese Journal of Applied Ecology, 2005, 16(9): 1655-1660. |
| [13] | Qiao Y N, Yin G C, Luo Y. Dynamics of total organic carbon (TOC) in hydrological processes and its contributions to soil organic carbon pools of three successional forest ecosystems in southern China. Ecology and Environmental Sciences, 2009, 18(6): 2300-2307. |
| [14] | Zhang G, Zeng G M, Jiang Y M, Huang G H, Li J B, Yao J M, Tan W, Xiang R, Zhang X L. Modelling and measurement of two-layer-canopy interception losses in a subtropical evergreen forest of central-south China. Hydrology and Earth System Sciences Discussions, 2006, 10(1): 65-77. |
| [15] | Whitehead D, Kelliher F M. A canopy water balance model for a Pinus radiata stand before and after thinning. Agricultural and Forest Meteorology, 1991, 55(1/2): 109-126. |
| [16] | Chang Z Y, Bao W K, He B H, Yang Y C, He Q H. Interception and distribution effects of mixed artificial Pinus tabulae formis and Pinus armandi forests on precipitation in the upper reaches of Minjiang River. Journal of Soil and Water Conservation,2006, 20(6) 37-40. |
| [17] | Lankreijer H, Hendriks M J, Klaassen W. A comparison of models simulating rainfall interception of forests. Agricultural and Forest Meteorology, 1993, 64 (3/4): 187-199. |
| [18] | Xu L H, Shi Z J, Wang Y H, Xiong W, Yu P T. Canopy interception characteristics of main vegetation types in Liupan Mountains of China. Chinese Journal of Applied Ecology, 2010,21(10): 2487-2493. |
| [19] | Chiwa M, Crossley A, Sheppard L J, Sakugawa H, Cape J N. Throughfall chemistry and canopy interactions in a Sitka spruce plantation sprayed with six different simulated polluted mist treatments. Environmental Pollution, 2004, 127(1): 57-64. |
| [20] | Liu C P, Sheu B H. Dissolved organic carbon in precipitation, throughfall, stemflow, soil solution, and stream water at the Guandaushi subtropical forest in Taiwan. Forest Ecology and Management, 2003, 172(2/3): 315-325. |
| [21] | McDowell W H, Likens G E. Origin, composition, and flux of dissolved organic carbon in the Hubbard Brook Valley. Ecological Monographs, 1988, 58(3): 177-195. |
| [22] | Dalva M, Moore T R. Sources and sinks of dissolved organic carbon in a forested swamp catchment. Biogeochemistry, 1991, 15(1): 1-19. |
| [23] | Edmonds R L, Thomas T B, Blew R D. Biogeochemistry of an old-growth forested, Olympic National Park, Washington. Journal of the American Water Resources Association, 1995, 31: 409-419. |
| [24] | Quideau S A, Bockheim J G. Biogeochemical cycling following planting to red pine on a sandy prairie soil. Journal of Environmental Quality, 1997, 26(4): 1167-1175. |
| [25] | Moore T R, Jackson R J. Dynamics of dissolved organic carbon in forested and disturbed catchments, Westland, New Zealand. 2. Larry River. Water Resources Research, 1989, 25(6): 1331-1339. |
| [26] | Turkey H B. The leaching of substances from plants. Annual Review of Plant Physiology, 1970, 21(1): 305-324. |
| [27] | Peichl M, Moore T R, Arain M A, Dalva M, Brodkey D, McLaren J. Concentrations and fluxes of dissolved organic carbon in an age-sequence of white pine forests in Southern Ontario, Canada. Biogeochemistry, 2007, 86(1): 1-17. |
| [28] | Don A, Kalbitz K. Amounts and degradability of dissolved organic carbon from foliar litter at different decomposition stages. Soil Biology and Biochemistry, 2005, 37(12): 2171-2179. |
| [29] | James B R, Riha S J. pH buffering in forest soil organic horizons: Relevance to acid precipitation. Journal of Environmental Quality, 1986, 15: 229-234. |
| [30] | Guo X M. Study of affects of acidification on organic matter stability in red soils from south China. Guangzhou: South China Normal University, 2008,38-39. |
| [3] | 马雪华. 森林水文学. 北京: 中国林业出版社, 1993, 43-45. |
| [4] | 杨昕, 王明星. 陆面碳循环研究中若干问题的评述. 地球科学进展, 2001, 16(3): 427-435. |
| [11] | 罗艳, 周国逸, 张德强, 官丽莉, 欧阳学军, 褚国伟. 鼎湖山三种主要林型水文学过程中总有机碳浓度对比. 生态学报, 2004, 24(12): 2973-2978. |
| [12] | 尹光彩, 周国逸, 张德强, 王旭, 褚国伟, 刘艳. 鼎湖山针阔叶混交林水文学过程中总有机碳动态. 应用生态学报, 2005, 16(9): 1655-1660. |
| [16] | 常志勇, 包维楷, 何丙辉, 杨以翠, 何其华. 岷江上游油松与华山松人工混交林对降雨的截留分配效应. 水土保持学报, 2006, 20(6): 37-40. |
| [18] | 徐丽宏, 时忠杰, 王彦辉, 熊伟, 于澎涛. 六盘山主要植被类型冠层截留特征. 应用生态学报, 2010, 21(10): 2487-2493. |
| [30] | 郭杏妹. 酸化作用对华南红壤中有机质稳定性的影响研究. 广州: 华南师范大学, 2008,38-39. |