 2014, Vol. 34
2014, Vol. 34文章信息
- 孙守家, 孟平, 张劲松, 贾长荣, 任迎丰
- SUN Shoujia, MENG Ping, ZHANG Jinsong, JIA Changrong, REN Yingfeng
- 太行山南麓山区栓皮栎-扁担杆生态系统水分利用策略
- Ecosystems water use patterns of Quercus variabilis and Grewia biloba based on stable hydrogen and oxygen isotopes in the south aspect of Taihang Mountains
- 生态学报, 2014, 34(21): 6317-6325
- Acta Ecologica Sinica, 2014, 34(21): 6317-6325
- http://dx.doi.org/10.5846/stxb201302030225
-
文章历史
- 收稿日期:2013-2-2
- 网络出版日期:2014-3-13






2. 济源市国有大沟河林场, 济源 454650
2. Dagou-he National Forest Farm, Jiyuan 454650, China
太行山区南麓地处华北中心,对华北平原起着极为重要的生态保障与调节作用,但土层薄瘠、土石裸露和坡度较大等劣质立地条件导致植物生长困难。加上近现代人类不合理的土地利用,造成植被严重破坏,水土流失加剧,形成许多荒山,使得该区域的生态环境十分脆弱。为了改善当地的生态环境,急需对裸露山区进行植被恢复,但该地区处于干旱半干旱季风气候带,降水较少且集中在7—9月,加上土壤总量少、储水能力差以及岩石渗漏性强等因素,水资源亏缺仍然是生态建设的主要限制性因素。
植物采取不同的策略来适应水分匮缺的环境,是否能在干旱的环境下生存与植物的水分来源密切相关。氢氧稳定同位素被认为是水的指纹,近年来被广泛应用到植物水分来源研究中[1, 2],已有研究证明除少数盐生或旱生植物外[3, 4],大部分植物根系在吸水过程和运输至未栓化树枝之前不会发生同位素分馏[5],木质部导管(管胞)内氢氧稳定同位素保持与来源水分相同的同位素组成。通过对比木质部水分与各水源同位素组成差异,可以区分出植物对不同水源的相对使用量[6]。当水分来源较少时,可以用二项或三项线性混合模型[7]获得植物的水分来源比例[8, 9],但线性模型不适合计算多个水分来源,Phillips的IsoSource模型为定量区分植物对多个水源的利用比例提供了方便[10, 11],且该模型可以根据氢和氧2种同位素值来计算植物吸收多个水源的相对比例范围,近几年来结合IsoSource模型分析植物水源的研究越来越多[12, 13]。目前,对太行山区水分问题的研究已由单纯分析土壤水的运移规律及空间异质性[14],转向植物的抗旱特征[15]和水分来源的研究[16, 17]。但由于方法限制,对环境条件脆弱的太行山南麓森林生态系统中不同植物水分利用策略研究较少。
栓皮栎(Quercus variabilis Blume.)是太行山区的典型优势树种,在有限的水分条件下与扁担杆(Grewia biloba)形成了稳定的自然群落。栓皮栎-扁担杆生态系统表现出较强的生态适应性,可能的原因是栓皮栎与扁担杆:1)有不同的水分来源;2)控制水分耗散上存在差异,从而能兼顾“开源”与“节流”的关系来有效利用水分。因此,本研究选择栓皮栎-扁担杆生态系统为试验对象,运用稳定同位素技术和使用IsoSource模型定量区分其水分来源,结合气孔导度、蒸腾速率和叶片相对含水量的变化来分析栓皮栎与扁担杆在控制水分耗散上的差异,从而确定其水分适应机制,进而为太行山南麓山区荒山绿化和植被恢复的树种配置研究提供科学参考。
1 材料与方法 1.1 试验地概况试验于2011年在中国林业科学研究院黄河小浪底森林生态站进行(图 1),该站隶属于中国森林生态系统定位研究网络(CFERN),地处河南省济源市境内的太行山南段与黄河流域的交界处,定位站中心地区海拔410 m,属暖温带大陆性季风气候,年均气温12.4—14.3 ℃,年日照时数2367.7 h,年日照率54%,历年平均降水量641.7 mm,其中,6—9月为438.0 mm,占全年的68.3%。

|
| 图 1 小浪底森林生态站位置 Fig. 1 Location of Xiaolangdi forest ecology observation station |
观测区的土壤为棕壤,但土层较薄,厚度一般为20—50 cm(图 2),土被分布极不均匀,部分基岩出露,土层下为松散的石灰岩风化层和基岩裂隙,土壤水分易下渗入风化岩体和基岩裂隙形成裂隙水。当裂隙水遇到不透水的页岩层,易潜水出露形成泉水[18]。观测区内有一口四季不断的泉眼,为本研究提供泉水,一口60 m的深水井提供深层地下水。

|
| 图 2 研究区的土壤结构 Fig. 2 Soil texture in study area |
3 块样地均位于阳坡,面积为20m×20m,栓皮栎和扁担杆组成乔灌生态系统,平均高度分别为11.6 m和1.7 m,栓皮栎生长在生态系统上层,郁闭度为0.84,扁担杆生长在栓皮栎林下,约占总盖度的60.56%。于2011年4月25日(旱季初期)、6月14日(旱季后期)、7月10日(雨季初期)和9月23日(雨季后期)分别进行采样。两个树种各选6株成熟植株,每株选已木质化的1年生枝条作为1个样品,为避免蒸腾引发同位素分馏,快速剥去树皮后装入10 ml的采样瓶并用帕拉胶密封,在所选植株下使用洛阳铲分别钻取0—10 cm、10—20 cm、20—40 cm和40—60 cm深度的土壤,迅速装入试剂瓶以避免分馏,一起储藏在0—5℃的便携式冷藏包内,带回实验室后冷冻(低于-20℃)保存。降雨时,对单次雨量超过5 mm的降水均进行收集,每次降水收集3瓶。在采集枝条样品的前一天和当天各采集泉水和地下水样品1次,每次3瓶。雨水、泉水和地下水样品用帕拉胶密封后冷藏保存(1—4℃)。
1.2.2 样品处理与测定用低温真空抽提仪器来提取土壤和枝条样品中的水,雨水、泉水和地下水样品使用0.45μm水系过滤器滤去杂质,使用DLT-100液态水同位素分析仪(Los Gatos Research Inc.,USA)测定液态水样品的氢氧同位素,输出结果是以相对于V-SMOW (Vienna Standard Mean Ocean Water)的千分率(‰)给出,:
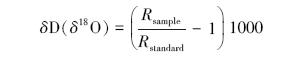
式中,Rsample 和Rstandard分别表示样品和国际通用标准物D/H和18O/16O的摩尔比率,氘同位素比率(δD)的测量精度是±0.5‰,氧同位素比率(δ18O)的测量精度是±0.2‰。
1.2.3 土壤含水量测定在林地10 cm深的土壤中安装水分传感器EC-H2O (Decagon Inc.,美国),通过AR5数据采集器(Avalon Inc.,美国)自动记录土壤含水量(θv),设定1 min采集1次,每10 min输出1组平均值。
1.2.4 叶片蒸腾及相对含水量测定蒸腾测定选择树冠中上部长势一致1—2年生枝条上的成熟叶片,使用Li-6400 (LI-COR公司,美国)测定蒸腾速率和气孔导度,外加红光LED光源,光照强度为1 000 μmol m-2 s-1,每株树测定6片,同时收集叶片放入密封袋中回实验室称量初始鲜重后迅速放入清水中浸泡5 h后,取出并擦拭干叶片表面水分,称取饱和鲜重,经105℃杀青30 min后,在75℃下烘到恒重,称干重后计算:
叶片 相对含水量(%)= (初始鲜重-干重)/(饱和鲜重-干重)×100%
1.3 数据处理为了方便用IsoSource模型计算,将土壤水划分为0—20 cm、20—40 cm和40—60 cm 3层,其中0—20 cm土壤水是根据Snyder[1]方法将0—10cm和10—20 cm同位素值的进行加权获得0—20 cm同位素值,权重是各层土壤含水量占总含水量的百分比。水在岩石裂隙中基本不受蒸发分馏的影响[19],裂隙水遇到隔水层时易形成泉水,故泉水与裂隙水有着相同或者相似的同位素值。深层地下水同位素相对稳定,受短期内降水的影响较小,直接从深井中采集。初步判断水分来源后,将木质部水、土壤水、泉水和地下水的同位素值代入IsoSource模型[11](免费获取地址http://www.epa.gov/wed/pages/models/stableIsotopes/isosource/isosource.htm)来计算不同植物对可能水源的利用比例。因水分来源多,将来源增量(Increment)和平衡公差(Tolerance)分别设置为2%和0.5[12]。使用Excel 2010进行整理数据并作图,用IBM SPSS Statistics 20分析软件对不同深度土壤进行One-way ANOVA分析并用最小显著差数法(LSD)进行多重比较,统计分析水平为P<0.05。
2 结果与分析 2.1 降水分布、雨水同位素特征及土壤相对含水量变化图 3结果显示,3—6月底共降雨11次,单次降雨最大量为23.8mm,总降雨量为80.50 mm,降水总量较少,属于旱季;7月至10月中旬共降雨28次,单次降雨最大量为90.3 mm,总降雨量为643.6 mm,降水较多,属于雨季。雨水δ18O值在-14.84‰—-1.94‰之间变化,δD值约为-106.50‰—-1.15‰。其中5月份降雨的δD和δ18O值较大,分别为-7.86‰和-2.84‰,8月份降雨的δD和δ18O值较小,分别为-84.77‰和-11.96‰。

|
| 图 3 降水分布及其氧同位素值的变化 Fig. 3 Precipitation distribution and δ18O values in 2011 |
3—6月的土壤含水量均值为14.14%,7—10月的均值为25.01%,雨季土壤含水量高于旱季约76.87%(图 4)。4月25日和6月14日的土壤含水量分别为13.45%和10.23%,旱季初期土壤含水量低于均值约4.87%,但旱季后期的土壤含水量低于旱季均值高达27.65%,表明旱季后期土壤缺水较严重。7月10日和9月21日土壤含水量分别为25.84%和25.44%,略高于雨季的均值,土壤水分充沛。

|
| 图 4 土壤含水量变化,圆点分别代表4个测试日期 Fig. 4 Variations of soil water content and the circular points represent four measurement date |
表 1显示测定时期内土壤δ18O和δ D值分别在-15.83‰—-0.18 ‰和-96.91‰—-11.26 ‰之间变化,但受环境因素影响,旱季土壤δ18O和δD值分别在-8.74‰—-0.18 ‰和-64.52‰—-11.26 ‰之间变化,明显高于雨季-15.83‰—-9.31 ‰和-96.91‰—-44.31 ‰的变化范围。不同测定时间的土壤水分的δ18O和δD值存在明显差别,图 5结果显示4月25日和6月14日的0—20 cm土壤δ18O和δD值最高,20—40 cm次之,40—60 cm最低,0—20 cm的δ18O和δD值与20—40 cm、40—60 cm差异显著。7月10日0—20 cm土壤δ18O和δD值略高于20—40 cm和40—60 cm,9月23日20—40 cm土壤δ18O和δD值略高于0—20 cm和40—60 cm,但三者之间差异并不显著。
| 测定季节 Season | 深度/cm Depth | 样品 Sample | 氧同位素 δ18O value ‰ | 氘同位素 δD value ‰ | ||||||
| 最大值 Max | 最小值 Min | 平均值 mean | 标准差 SD | 最大值 Max | 最小值 Min | 平均值 mean | 标准差 SD | |||
| 旱季 | 0—20 | 21 | -0.18 | -4.01 | -2.28 a | 1.06 | -11.26 | -32.59 | -22.50 a | 6.38 |
| Dry season | 20—40 | 20 | -4.51 | -6.91 | -5.58 b | 0.84 | -37.22 | -56.62 | -46.25 b | 6.47 |
| 40—60 | 23 | -4.5 | -8.74 | -6.66 b | 1.25 | -36.66 | -64.52 | -48.07 b | 7.68 | |
| 雨季 | 0—20 | 22 | -9.31 | -15.26 | -12.21 a | 1.80 | -44.31 | -90.58 | -70.94 a | 17.84 |
| Wet season | 20—40 | 22 | -9.90 | -15.13 | -12.58 a | 1.40 | -52.58 | -94.56 | -76.94 a | 15.11 |
| 40—60 | 24 | -9.35 | -15.83 | -12.66 a | 2.08 | -55.32 | -96.91 | -78.29 a | 12.36 | |
泉水的δ18O和δD值小幅波动,4月25日、6月14日、7月10日和9月23日δ18O和δD值分别为-7.01‰和-43.25‰、-7.22‰和-48.19‰、-8.59‰和-55.38‰、-9.67‰和-60.21‰,呈现逐渐降低趋势。测定期间地下水的δ18O和δD值分别为-9.05‰和-61.35‰、-8.71‰和-59.98‰、-8.47‰和-68.77‰、-8.87‰和-64.19‰,同位素值相对稳定(图 5)。
栓皮栎和扁担杆的木质部氢氧同位素值在旱季和雨季差异较大,4月25日和6月14日栓皮栎的δ18O和δD值明显低于扁担杆,而在7月10日和9月23日栓皮栎的δ18O和δD值高于扁担杆(图 5)。旱季,栓皮栎同位素值比扁担杆更接近泉水的同位素值。在雨季扁担杆同位素值比栓皮栎更接近降水的同位素值,而距离泉水和地下水的同位素值更远。

表 2结果显示,4月25日栓皮栎水分主要来源于20—40 cm土壤,其比例均值占总水分的81.90%,0—20 cm和40—60 cm分别占1.4%和9.6%;扁担杆水分主要来源于0—20 cm,约占总水分的51.2%,20—40 cm和40—60 cm分别占34.7%和9.4%。此外,栓皮栎和扁担杆灌木对泉水吸收比例均值分别占2.4%和2.6%,对地下水吸收比例均值分别占4.6%和2.1%。6月14日栓皮栎吸收水分主要来自于40—60 cm土壤,约占49.7%,扁担杆吸收水分主要来自于20—40 cm,占总水分的47.8%。栓皮栎利用泉水和地下水较多,分别约占19.6%和4.9%,而扁担杆利用泉水和地下水的比例仅为1.2%和1.9%。7月10日栓皮栎水分主要来源于20—40 cm,占总水分的39.6%,0—20 cm和40—60 cm水分分别占18.6%和19.3%,扁担杆水分主要来源于0—20 cm土壤,约占46.1%,20—40 cm和40—60 cm水分分别占23.2%和17.2%。栓皮栎利用泉水比例达到17.9%,而扁担杆仅有6.3%。9月23日栓皮栎和扁担杆水分主要来源于0—20 cm,分别约占52.3%和64.8%,泉水利用比例下降,分别占2.6%和4.3%。
| 测定时间 Date | 树种 Species | 水分来源 Water sources/% | ||||
| 0—20 cm Soil layer | 20—40 cm Soil layer | 40—60 cm Soil layer | 泉水 Spring water | 地下水 Ground water | ||
| *最小值—最大值的平均值 | ||||||
| 4月25日Apr. 25 | 栓皮栎Q. variabilis | 1.4 (0—8) | 81.9 (64-98) | 9.6 (0—32) | 2.4 (0—12) | 4.6 (0—16) |
| 扁担木G. biloba | 51.2 (46-60) | 34.7 (16-52) | 9.4 (0—32) | 2.6 (0—12) | 2.1 (0—10) | |
| 6月14日Jun. 14 | 栓皮栎Q. variabilis | 10.5 (2-20) | 15.3 (0—66) | 49.7 (2-98) | 19.6 (0—80) | 4.9 (0—22) |
| 扁担杆G. biloba | 45.1 (32-54) | 47.8 (44-52) | 4.0 (0—16) | 1.2 (0—6) | 1.9 (0—8) | |
| 7月10日 Jul. 10 | 栓皮栎Q. variabilis | 18.6 (0—62) | 39.6 (0—74) | 19.3 (0—50) | 17.9 (0—54) | 4.7 (0—20) |
| 扁担杆G. biloba | 46.1 (0—90) | 23.2 (0—64) | 17.2 (0—56) | 6.3 (0—20) | 7.1 (0—22) | |
| 9月23日Sept.23 | 栓皮栎Q. variabilis | 52.3 (26-92) | 20.7 (0—68) | 22.1 (0—66) | 2.6 (0—8) | 2.3 (0—8) |
| 扁担杆G. biloba | 64.8 (24-90) | 16.6 (0—54) | 7.0 (0—16) | 4.3 (0—16) | 7.2 (0—21) | |
旱季与雨季的栓皮栎和扁担杆的蒸腾速率变化规律存在差异。图 6结果显示,4月25日和6月14日,栓皮栎蒸腾速率日变化均成双峰变化曲线,最大值在10:00,12:00的蒸腾速率比10:00和14:00分别下降了13.48%和8.98%,7月10日和9月23日的蒸腾速率成现单峰曲线,最大值在12:00左右。扁担杆蒸腾日变化速率均成单峰曲线,4月25日和6月14日的最大值在10:00左右,7月10日和9月23 日的最大值延迟到12:00。从树种上来看,旱季和雨季中扁担杆的日均蒸腾速率均比栓皮栎小,分别小了46.94%和31.19%。

|
| 图 6 试验期间栓皮栎和扁担杆蒸腾速率、气孔导度和相对含水量变化 Fig. 6 Variations of transpiration rate,stomatal conductance and leaf relative water content of Q. variabilis and G. biloba in experimental date |
4月25日和6月14日,栓皮栎气孔导度均成双峰变化,最大值在10:00,12:00的气孔导度比10:00和14:00分别下降了17.79%和12.13%(图 6),7月10日和9月23日的气孔导度均成单峰曲线,最大值在12:00左右。扁担杆气孔导度均成单峰曲线变化,4月25日和6月14日的最大值在10:00左右,10:00—14:00变化平缓;7月10日和9月23日的最大值在12:00。从树种上来看,旱季和雨季中扁担杆的日均气孔导度均比栓皮栎小,分别小了30.58%和26.50%。
4月25日和6月14日,栓皮栎叶片相对含水量从8:00至12:00迅速下降,分别下降了约8.70%和7.98%,随后逐渐升高,呈现“V”型变化,7月10日和9月23日的叶片相对含水量变化较缓,最小值比最大值分别下降了约2.11%和4.14%。扁担杆叶片相对含水量变化相对平缓,4个测定日内最小值比最大值分别下降了约4.36%,3.99%,0.64%和0.95%。从树种上来看,旱季和雨季中扁担杆叶片相对含水量比栓皮栎高,分别高了5.07%和6.45%(图 6)。
3 讨论 3.1 旱雨季中栓皮栎和扁担杆的水分来源差异稳定同位素方法既能避免对植物地下根系的破坏,又能区分和量化植物的多个可能水分来源和比例[21],适合用于山区植物水分利用策略的研究。植物是否能在干旱的环境下生存与其水分来源密切相关,本文结果显示随着旱季的延续,栓皮栎和扁担杆水分主要来源发生变化,分别从旱季初期的0—20 cm和20—40 cm增加到旱季后期的20—40 cm和40—60 cm,且旱季后期栓皮栎还利用了19.6%的泉水。表明栓皮栎和扁担杆均采取了“开源”的方式吸收深层土壤水分以适应干旱环境,这与以往研究[9, 22]观测到降水不稳定时植物更倾向于利用深层土壤水的现象相似。尽管由于地质条件限制,无法完整地调查栓皮栎和扁担杆的根系分布,但已有的研究表明深根性树木主要利用深层土壤水或地下水[6, 9],浅根系植物则主要利用来自最近降雨或浅层土壤水[23],扁担杆倾向于利用浅层土壤水分,栓皮栎则利用深层土壤水分较多,水分来源不同可能是由于根系分布上的差异所导致。除了土壤水分,栓皮栎在旱季还利用了部分泉水和地下水,达到19.6%和4.9%,远高于扁担杆的1.2%和1.9%,可能是因为栓皮栎根系较深,部分根系已经生长到岩石裂隙中,当土壤水分不能完全满足其生长需要,栓皮栎会利用更深的风化基岩层水分来维持生存[24]。Rose等[2]的相关研究表明在旱季当土壤水分匮乏时Pinus jeffreyi主要利用风化岩层储存的水分,Querejeta等[19]对尤卡坦半岛喀斯特地区和Nie等[25]对基岩生境的相关研究也得出类似的结论。由于栓皮栎和扁担杆水分来源不同,使得它们在旱季能避开用水冲突,为其形成稳定的生态系统提供了基础。
雨季来临后,栓皮栎和扁担杆水分来源逐步变浅,分别由旱季后期的40—60 cm和20—40 cm上升为雨季初期的20—40 cm和0—20 cm,以及雨季后期二者水分均主要来源于0—20 cm土壤。栓皮栎-扁担杆生态系统在雨季中对表层土壤水分的利用比例明显增加,其原因可能来自两个方面:一方面是雨季中表层土壤水分有效性增加,另一方面是土壤表层相对疏松,枯枝落叶形成的有机质在表层土壤中积累,水分供给充足时,表层活性根数量会大量增加[17],且表层根系吸收水分时消耗能量相对较少,植物优先利用表层水分[26]。从旱季初期到雨季后期,栓皮栎和扁担杆的主要水分来源经历了从浅到深再从深到浅的过程,出现此种现象的原因可能与栓皮栎和扁担杆根系的二态性有关[27]。不过,根系的二态性转化需要过程,虽然雨季初期栓皮栎的主要水分来源于20—40 cm,深度比旱季后期有所升高,但IsoSource结果显示其0—20 cm土壤水分仅占23.2%,并且还利用了17.9%的泉水,到了雨季后期栓皮栎0—20 cm的水分来源才占主要部分,这表明栓皮栎根系在雨季初期并没有完全改变其功能根的深度,只有当降水增加到一定的阈值时,栓皮栎表层根系才能形成并保持吸收表层土壤水分的功能[28]。与栓皮栎相比,扁担杆较少地利用以泉水和地下水,可能是基岩裂隙的泉水和地下水位置较深,而扁担杆是灌木,深层根系较少且活性差,在表层土壤水分充足时较少或者不使用泉水和地下水,这与Nie等[25]观测到的雨季中鹅掌柴主要利用泉水有所不同。
3.2 栓皮栎和扁担杆在控制水分耗散上的差异叶片是蒸散耗水的关键部位,其含水量的高低能灵敏地反映出植株水分盈亏程度[29]。本文显示无论在旱季初期还是后期,栓皮栎和扁担杆叶片相对含水量均低于雨季,反映出旱季栓皮栎和扁担杆树体水分比雨季亏缺严重。虽然处于同一生态系统,但旱季中栓皮栎和扁担杆叶片相对含水量日变化趋势不同,栓皮栎成“V”型日变化,可能是因为其生长在生态系统的上层,叶片在较强太阳辐射下快速失水,扁担杆则生长在生态系统下层,太阳辐射较弱,含水量成“U”型缓慢变化。叶片含水量下降是植物对水分亏缺最常见的反应,是一种降低叶片消耗的机制[30]。当水分亏缺时,植物利用生理机制来优化叶片蒸腾以减少水分耗散[31],起到一定“节流”作用,将有限的水分来满足其正常生长需要。本文显示栓皮栎在旱季中蒸腾速率和气孔导度均成双峰曲线的日变化,中午部分气孔关闭,从而降低蒸腾来调节体内水分运输,这对高蒸腾情况下维持栓皮栎体内水分平衡非常必要[32],在一定程度上减少对土壤水的消耗,改善了对土壤水利用时效性[33]。旱季扁担杆的蒸腾速率和气孔导度日变化是单峰曲线,表明其蒸腾速率和气孔导度尚未达到一定的阈值,没有蒸腾午休现象[31],然而旱季中扁担杆蒸腾速率和气孔导度均比栓皮栎小,分别小了46.94%和30.58%,表明扁担杆是通过较小的蒸腾来降低水分耗散。栓皮栎-扁担杆生态系统两种树木长期面临不同环境条件,因而在其生长、形态和结构方面会产生不同的反应模式[34]。栓皮栎采用了高蒸腾但蒸腾午休方式来优化水分蒸散,而扁担杆采取生长在生态系统下层和较低的叶片蒸腾速率来降低水分的耗散[35]。
4 结论针对栓皮栎-扁担杆生态系统水分利用的研究表明,雨季栓皮栎和扁担杆水分来源相同,均主要来源于表层土壤,但旱季中栓皮栎较多主要利用深层土壤水分和泉水,扁担杆倾向于利用浅层土壤水分,二者水分来源的不同,使得栓皮栎与扁担杆在干旱季节避开用水冲突。同时,在旱季栓皮栎通过部分气孔关闭降低体内水分耗散的策略来适应水分匮缺的环境,而扁担杆生长在栓皮栎-扁担杆生态系统下层,通过栓皮栎遮挡和较低的蒸腾来降低水分的耗散。栓皮栎和扁担杆通过不同的策略合理地利用旱季中有限的水分,其组成的生态系统能适应太行山南麓山区脆弱的生态环境。
| [1] | Snyder K A, Williams D G. Water sources used by riparian trees varies among stream types on the San Pedro River, Arizona. Agricultural and Forest Meteorology, 2000, 105(1/3): 227-240. |
| [2] | Rose K L, Graham R C, Parker D R. Water source utilization by Pinus jeffreyi and Arctostaphylos patula on thin soils over bedrock. Oecologia, 2003, 134(1): 46-54. |
| [3] | Lin G H, Sternberg L da S L. Hydrogen isotopic fractionation by plant roots during water uptake in coastal wetland plants // Ehleringer J R, Hall A E, Farquhar G D, eds. Stable Isotopes and Plant Carbon-Water Relations. New York: Academic Press, New York. 1993: 497-510. |
| [4] | Ellsworth P Z, Williams D G. Hydrogen isotope fractionation during water uptake by woody xerophytes. Plant and Soil, 2007, 291(1/2): 93-107. |
| [5] | Brunel J P, Walker G R, Kennett-Smith A K. Field validation of isotopic procedures for determining sources of water used by plants in a semi-arid environment. Journal of Hydrology, 1995, 167(1/4): 351-368. |
| [6] | Dawson T E, Mambelli S, Plamboeck A H, Templer P H, Tu K P. Stable isotopes in plant ecology. Annual Review of Ecology and Systematics, 2002, 33(1): 507-559. |
| [7] | White J W C, Cook E R, Lawrence J R. The D/H ratios of sap in trees: implications for water sources and tree ring D/H ratios. Geochimicaet Cosmochimica Acta, 1985, 49(1): 237-246. |
| [8] | Sekiya N, Yano K. Do pigeon pea and sesbania supply groundwater to intercropped maize through hydraulic lift? Hydrogen stable isotope investigation of xylem waters. Field Crops Research, 2004, 86(2/3): 167-173. |
| [9] | McCole A A, Stern L A. Seasonal water use patterns of Juniperus ashei on the Edwards Plateau, Texas, based on stable isotopes in water. Journal of Hydrology, 2007, 342(3/4): 238-248. |
| [10] | Phillips D L, Gregg J W. Source partitioning using stable isotopes: coping with too many sources. Oecologia, 2003, 136(2): 261-269. |
| [11] | Phillips D L, Newsome S D, Gregg J W. Combining sources in stable isotope mixing models: alternative methods. Oecologia, 2005, 144(4): 520-527. |
| [12] | Wang P, Song X F, Han D M, Zhang Y H, Liu X. A study of root water uptake of crops indicated by hydrogen and oxygen stable isotopes: a case in Shanxi Province, China. Agricultural Water Management, 2010, 97(3): 475-482. |
| [13] | Nie Y P, Chen H S, Wang K L Yang J. Water source utilization by woody plants growing on dolomite outcrops and nearby soils during dry seasons in karst region of Southwest China. Journal of Hydrology, 2012, 420(14): 264-274. |
| [14] | Wang Q R, Tan X F, Gao J. Spatiotemporal variability of soil moisture under different land use patterns of Taihang Mountains. Journal of Soil and Water Conservation, 2008, 22(4): 100-110. |
| [15] | Zhao Y, Chen Z, Fan W, Wang Q, Yang X T. Water utilization characteristics of 7 typical plant species in hilly region of Taihang Mountains. Science of Soil and Water Conservation, 2010, 8(5): 61-66. |
| [16] | Sun S J, Meng P, Zhang J S, Huang H, Wan X C. Deuterium isotope variation and water use in an agroforestry system in the rocky mountainous area of North China. Acta Ecologica Sinica, 2010, 30(14): 3665-3671. |
| [17] | Sun S J, Meng P, Zhang J S, Wan X C. Variation in soil water uptake and its effect on plant water status in juglans regia L. during dry and wet seasons. Tree Physiology, 2011, 31(12): 1378-1389. |
| [18] | Fan K F, Yang D C. Geomorphologic system in Taihang Mountain area. Journal of Changchun Institute of Technology: Natural Science Edition, 2006, 7(1): 51-53, 62-62. |
| [19] | Querejeta J I, Estrada-Medina H, Allen M F, Jiménez-Osornio J J, Ruenes R. Utilization of bedrock water by Brosimum alicastrum trees growing on shallow soil atop limestone in a dry tropical climate. Plant and Soil, 2006, 287(1/2): 187-197. |
| [20] | Durand J L, Bariac T, Ghesquière M, Biron P, Richard P, Humphreys M, Zwierzykovski Z. Ranking of the depth of water extraction by individual grass plants, using natural 18O isotope abundance. Environmental and Experimental Botany, 2007, 60(1): 137-144. |
| [21] | Schwinning S, Ehleringer J R. Water use trade-offs and optimal adaptations to pulse-driven arid ecosystems. Journal of Ecology, 2001, 89(3): 464-480. |
| [22] | Liu Y H, Xu Z, Duffy R, Chen W L, An S Q, Liu S R, Liu F D. Analyzing relationships among water uptake patterns, rootlet biomass distribution and soil water content profile in a subalpine shrubland using water isotopes. European Journal of Soil Biology, 2011, 47(6): 380-386. |
| [23] | Hubbert K R, Beyers J L, Graham R C. Roles of weathered bedrock and soil in seasonal water relations of Pinus jeffreyi and Arctostaphylos patula. Canadian Journal of Forest Research, 2001, 31(11): 1947-1957. |
| [24] | Nie Y P, Chen H S, Wang K L, Tan W, Deng P Y, Yang J. Seasonal water use patterns of woody species growing on the continuous dolostone outcrops and nearby thin soils in subtropical China. Plant and Soil, 2011, 341(1/2): 399-412. |
| [25] | Schenk H J. The shallowest possible water extraction profile: a null model for global root distributions. Vadose Zone Journal, 2008, 7(3): 1119-1124. |
| [26] | Williams D G, Ehleringer J R. Intra- and interspecific variation for summer precipitation use in Pinyon-juniper woodlands. Ecological Monographs, 2000, 70(4): 517-537. |
| [27] | Dawson T E, Pate J S. Seasonal water uptake and movement in root systems of Australian phraeatophytic plants of dimorphic root morphology: a stable isotope investigation. Oecologia, 1996, 107(1): 13-20. |
| [28] | Wang J H, Zhao C J, Huang W J, Guo X W, Li H X. Effect of soil water content on the wheat leaf water content and the physiological function. Journal of Triticeae Crops, 2001, 21(4): 42-47. |
| [29] | Schonfeld M A, Johnson R C, Carver B F, Mornhinweg D W. Water relations in winter wheat as drought resistance indicators. Crop Science, 1988, 28(3): 512-516. |
| [30] | Sun J K, Zhang W H, Lu Z H, Liu X C. Effects of drought stress on gas exchange characteristics and protective enzyme activities in Elaeagnus angustifolia and Grewia. biloba G. Don var. parviflora seedlings. Acta Ecologica Sinica, 2009, 29(3): 1330-1340. |
| [31] | Lo Gullo M A, Salleo S. Water storage in the wood and xylem cavitation in 1-year-old twigs of Populus deltoides Bartr. Plant, Cell and Environment, 1992, 15(4): 431-438. |
| [32] | Salleo S, Nardini A, Pitt F, Lo Gullo M A. Xylem cavitation and hydraulic control of stomatal conductance in Laurel (Laurus nobilis L.). Plant, Cell and Environment, 2000, 23(1): 71-79. |
| [33] | Álvarez-Cansino L, Zunzunegui M, Díaz Barradas M C, Esquivias M P. Physiological performance and xylem water isotopic composition underlie gender-specific responses in the dioecious shrub Corema album. Physiologia Plantarum, 2010, 140(1): 32-45. |
| [34] | Bacelar E A, Moutinho-Pereira J M, Gonçalves B C, Ferreira H F, Correia C M. Changes in growth, gas exchange, xylem hydraulic properties and water use efficiency of three olive cultivars under contrasting water availability regimes. Environmental and Experimental Botany, 2007, 60(2): 183-192. |
| [14] | 王齐瑞, 谭晓风, 高峻. 太行山山前坡地不同土地利用方式下土壤水分的时空变异特征. 水土保持学报, 2008, 22(4): 100-103, 110-110. |
| [15] | 赵勇, 陈桢, 樊巍, 王谦, 杨喜田. 太行山低山丘陵区7种典型植物水分利用特征. 中国水土保持科学, 2010, 8(5): 61-66. |
| [16] | 孙守家, 孟平, 张劲松, 黄辉, 万贤崇. 华北石质山区核桃——绿豆复合系统氘同位素变化及其水分利用. 生态学报, 2010, 30(14): 3717-3726. |
| [18] | 樊克锋 杨东潮. 论太行山地貌系统. 长春工程学院学报: 自然科学版, 2006, 7(1): 51-53, 62-62. |
| [28] | 王纪华, 赵春江, 黄文江, 郭晓维, 李鸿祥. 土壤水分对小麦叶片含水量及生理功能的影响. 麦类作物学报, 2001, 21(4): 42-47. |
| [30] | 孙景宽, 张文辉, 陆兆华, 刘新成. 沙枣(Elaeagnus angustifolia)和孩儿拳头(Grewia. biloba G. Don var. parviflora)幼苗气体交换特征与保护酶对干旱胁迫的响应. 生态学报, 2009, 29(3): 1330-1340. |